DOI 10.17480/psk.2018.62.5.306
대식세포에서 Pam3CSK4로 유도된 면역관용의 NF-κB/MAPK 신호 조절을 통한 항염증 효과
한상철 · 강나진 · 박경민 · 박지선 · 엄영은 · 강희경 · 유은숙# 제주대학교 의학전문대학원 의학과
(Received May 14, 2018; Revised August 30, 2018; Accepted September 15, 2018)
Anti-inflammatory Effect of Tolerance Induced by Pam3CSK4 in Murine Macrophage RAW 264.7 Cells via Inhibitor of NF- κB/MAPK Signaling Pathway
Sang-Chul Han, Na-Jin Kang, Kyungmin Park, Jiseon Park, Youngeun Eum, Hee-Kyoung Kang, and Eun-Sook Yoo#
Department of Medicine, Jeju National University School of Medicine 102 Jejudaehak-ro, Jeju-si, Jeju Special Self-Governing Province, 63243
Abstract — Endotoxin tolerance is an essential mechanism for immune-homeostatic response that suppresses excessive inflammatory cytokine production. It is induced by repeated exposure to endotoxins, such as lipopolysaccharide (LPS). Many recent studies have investigated LPS tolerance, but the molecular mechanism for Pam3CSK4-tolerance in mouse mac- rophage is poorly understood. Pam3CSK4 binds to TLR1/2 and regulates several downstream signaling pathways, including NF-κB and MAPK pathways. In this study, pretreatment with Pam3CSK4 inhibited TNF-α and IL-6 production after sec- ondary Pam3CSK4 challenge in a time- and dose-dependent manner without cell cytotoxicity. Nontolerized RAW 264.7 cells responded to Pam3CSK4 challenge with the degradation of IκBα and phosphorylation of p38, ERK and JNK. On the other hand, activation of IκBα and MAPKs was diminished in Pam3CSK4-tolerized cells. We also investigated whether pre- treatment with Pam3CSK4 regulates the expression of signaling inhibitors TNFAIP3, SOCS1 and SOCS3. Compared with the nontolerized macrophages, the expression of A20, SOCS1 and SOCS3 was increased in Pam3CSK4-tolerized mac- rophages. These results indicate that Pam3CSK4-tolerized macrophages suppress pro-inflammatory responses and induce the expression of signaling inhibitors of TLR mechanism.
Keywords tolerance, Pam3CSK4, TNF-α inducible protein 3, suppressor of cytokine signaling 1/3
서 론 (Introduction)
세균의 지질단백의 아실화된 아미노 말단을 모방한 합성된 tripalmitoylated lipopeptide인 Pam3CysSerLys4 (Pam3CSK4)는 대식세포 표면의 Toll-like receptor 1/2 (TLR1/2)를 자극하여 하 위 세포신호전달경로인 extracellular signal-regulated kinase
(ERK), c-Jun N-terminal kinase (JNK), p38 kinase를 포함하는 mitogen-activated protein kinase (MAPK)와 nuclear factor kappa-B (NF-κB)를 활성화시키게 됨에 따라 inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2)를 발현시켜 nitric oxide (NO), prostaglandin E2 (PGE2)를 생성하게 하고 염 증성 사이토카인 interleukin (IL)-6, TNF-α, IL-1β의 발현을 증 가시켜 염증반응을 유도한다.1-3)대식세포에 있어 염증성 사이토 카인 및 단백질의 발현은 NF-κB에 의해 조절된다.4)무자극 상 태에서 NF-κB는 inhibitor of kappa B (IκB)와 결합하여 불활성 상태로 세포질에 존재하나 Pam3CSK4와 같은 자극이 가해지면 IκB는 IκB kinase (IKK)에 의해 인산화되어 분해되고 유리된 NF-κB는 세포핵으로 이동하여 다양한 염증성 인자들의 발현을 유도하게 된다.5-7)
#
Corresponding Author Eun-Sook Yoo
Department of Medicine, Jeju National University School of Medicine 102 Jejudaehak-ro, Jeju-si, Jeju Special Self-Governing Province, 63243
Tel.: 064-754-3847 Fax.: 064-702-2687 E-mail: [email protected]
Short Report
종설전사 조절 단백질인 TNF-α inducible protein 3 (TNFAIP3)로 알려진 A20은 세포 내 유비퀴틴 편집 단백질로써 TLR에 대한 NF-κB의 표적 유전자이며 정상적인 상태에서는 NF-κB가 일시 적으로 활성을 나타내거나 NF-κB가 활성화되면서 이차적으로 발현되지만, 음성 되먹임 작용을 통해 결과적으로 NF-κB의 활 성을 억제하는 중요한 인자이다.8,9) A20의 표적단백질은 TLR의 하위 신호전달경로인 TNF receptor associated factor 6 (TRAF6), NF-κB essential modulator (NEMO)와 Transforming growth factor β activating kinase 1 (TAK1) 등이 있고 A20은 K63 linked ubiquitin을 제어함으로써 이러한 단백질들의 활성을 억제
한다.10,11)결국 A20의 활성화는 다양한 질병의 발생을 억제하는
유전자의 역할을 하며 A20 유전자 변이, 비활성화, 결손 및 메 틸화는 제어되지 않는 NF-κB의 활성화를 초래하여 눈, 갑상샘, 타액샘, 간 등의 MALT 림프종 또는 부착부염, 강직성 관절염 및 류마티스관절염의 원인이 된다.12-14)
Suppressor of cytokine signaling (SOCS) 단백질은 염증 유 발 및 염증 억제 반응에 관여하는 Signal Transducer and Activator of Transcription (STAT) 단백질의 하위 표적 물질이 며 세포 내 다양한 기전들을 feed-back하여 다양한 신호 전달과 정을 억제하는 역할을 한다.15,16) SOCS의 발현을 증가시키는 cytokine에는 IL-2, IL-4, IL-6, IL-10, macrophage colony- stimulating factor (M-CSF), Granulocyte-macrophage colony- stimulating factor (GM-CSF), Interferon-γ 등이 있다.17) SOCS family member들 중 SOCS1은 Janus tyrosine kinase (JAK)1, JAK2, JAK3 등과 결합하여 다양한 신호전달 과정을 억제하고 T 세포에 IL-6를 자극하면 SOCS1에 의한 신호전달이 T helper 1 (Th1) 세포 반응을 억제하며 SOCS1의 feed-back이 차단될 경우 관절염이 발생한다고 알려져 있다.18-21) 또한 SOCS1유전자가 deletion된 쥐는 태어난 후 1~3주 내에 사망을 하지만 이러한 쥐에 IFN-γ 유전자도 같이 deletion시킨 쥐는 주산기 사망을 보 이지 않아 SOCS1의 작용에 있어 IFN-γ는 중요한 조절인자 임 이 확인되었다.22,23) IL-10, Transforming growth factor-beta (TGF-β)와 같은 염증 억제성 cytokine에 의한 신호전달에는 주 로 SOCS3가 관여한다.24,25)류마티스 관절염 환자의 활막 조직 에서 SOCS3의 발현이 증가되어 있고 antigen 또는 collagen으 로 유도된 관절염 동물 모델에 SOCS3유전자를 관절에 주입하 면 관절염이 완화됨을 보였으며 Dextran sulfate sodium (DSS) 로 유도된 대장염 쥐의 대장조직에서도 SOCS3의 발현이 대장 염 증상완화에 중요한 인자라고 알려져 있다.26)
면역 체계는 면역을 증진하는 면역반응과 체내에 존재하는 물 질 혹은 외부 물질에 과잉 면역을 억제 및 조절하는 면역관용으 로 구성되며, 이러한 두 가지 메커니즘이 균형을 이룸으로써 면 역 항상성이 유지된다. 면역관용에 있어 대식세포와 같은 항원 제시세포는 가장 먼저 면역반응을 조절하는 기능을 담당하고 있
다. 항원제시세포의 표면에 발현되는 molecule들은 T 세포 표면 의 molecule과 결합하여 신호전달 작용을 하거나 다양한 사이토 카인의 발현을 통해 면역관용을 유도하기 때문에 항원제시세포에 의한 면역관용 유도 메커니즘 연구는 대단히 중요하다.27,28)따라 서, 본 연구에서는 대식세포를 이용하여 면역관용 유도를 통해 염 증성 인자의 발현을 확인하고 그 작용기전을 조사하고자 하였다.
실험방법 (Experimental methods)
시약 및 기기
Dulbecco’s Modified Eagle Medium (DMEM)과 fetal bovine serum (FBS)은 GIBCO (Grand Island, NY, USA)에서 구입하였고 Pam3Cys-Ser-(Lys)4 (Pam3CSK4)는 InvivoGen (San Diego, CA, USA)에서 구입하여 사용하였다. 또한, Anti- IκB-α, anti-p38, anti-JNK, anti-ERK, anti-phospho-p38, anti- phospho-JNK, anti-phospho-ERK, anti-SOCS1, anti-SOCS3, anti-A20/TNFAIP3 항체는 Cell Signaling Technology (Beverly, MA, USA)에서 구입하였고, β-actin 항체는 Sigma Chemical Co.
(St. Louis, MO, USA)에서 구입하여 사용하였다. 사이토카인 측 정을 위해 Mouse IL-6와 TNF-α ELISA duoset kit를 R&D system (St. Louis, MO. USA)에서 구입하여 사용하였다.
세포배양
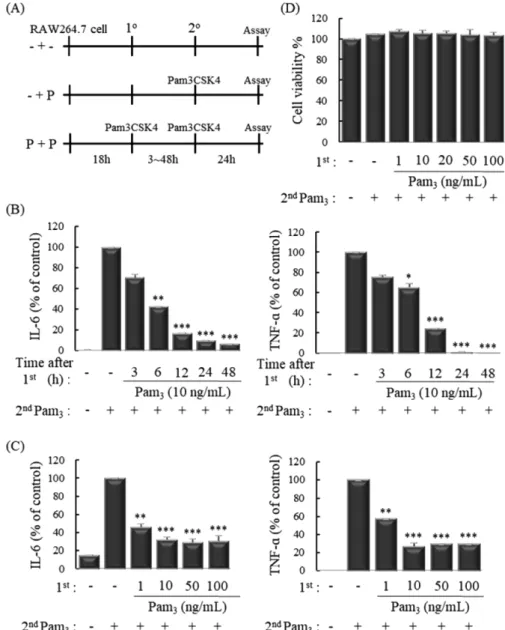
마우스 대식세포주인 RAW 264.7 세포는 ATCC (Rockville, MD, USA)에서 분양 받았고 세포주는 10% FBS와 1% 항생제 가 첨가된 DMEM을 이용하여 37oC, 5% CO2조건하에 배양하 였다. 면역관용 유도를 위해, RAW 264.7 세포에 1차 자극으로 Pam3CSK4 (10 ng/mL)를 24시간 동안 처리하였고 다시 2차 자 극으로 Pam3CSK4 (10 ng/mL)를 다양한 시간 동안 처리하여 면 역관용을 유도하였다(Fig. 1A).
세포 생존능 평가
세포 생존능은 EZ-cytox enhanced cell viability assay kit (itBIO, Korea)를 이용하여 측정하였다. RAW 264.7 세포에 1차 자극으로 Pam3CSK4 (1~100 ng/mL)를 24시간 동안 처리하였고 다시 2차 자극으로 Pam3CSK4 (10 ng/mL)를 24시간 동안 처리 후 배지에 WST-1 solution (2-(4-Iodophenyl)- 3-(4-nitrophenyl)- 5-(2,4-disulfophenyl)-2H-tetrazolium)을 넣고 3시간 반응시켰다.
각 well의 흡광도는 VerasaMax ELISA microplate reader (Molecular Devices Inc., CA, USA)를 이용하여 450 nm에서 측 정하였다.
Enzyme-linked Immunosorbent Assay (ELISA) RAW 264.7 세포에 1차 자극으로 Pam3CSK4 (1~100 ng/mL)
를 3~48시간 동안 처리하였고 다시 2차 자극으로 Pam3CSK4 (10 ng/mL)를 24시간 동안 처리하였다. 세포 배양액 내 IL-6와 TNF-α 농도는 ELISA kit를 이용하여 측정하였다. 간단히 말하 면, 배양액 100 uL를 각 96 well plates에 분주하여 상온에서 2 시간 반응시킨 후 마우스 IL-6, TNF-α 항체와 avidin-HRP를 이 용하여 반응하였고 기질반응 용액 A/B를 분주하여 빛을 차단한 상태에서 20분 경과 후 반응정지 용액을 이용하여 반응을 정지 시켰다. 흡광도 측정은 ELISA reader (450 nm)를 이용하였다.
Western Blot Assay
Pam3CSK4로 면역관용이 유도된 RAW 264.7 세포에 protein lysis buffer를 처리하고 단백질을 분리하여 실험에 사용하였다.
단백질 농도는 bovine serum albumin (BSA)을 사용하여 표준 화하였고, 단백질 정량은 bradford assay (Bio-Rad, Herculers, CA, USA)를 이용하였다. 동량의 lysate를 8% mini gel sodium docecyl sulfate poly-acrylamide gel electrophoresis (SDS- PAGE)를 이용하여 변성 분리한 후, PVDF membrane (Bio-Rad)
Fig. 1 − Pretreatment with Pam3CSK4 suppresses the expression of inflammatory cytokines on secondary challenge by Pam3CSK4. (A) Experimental design: - + -, no treatment; - + P, no treatment (1
o), followed by Pam3CSK4 challenge (2
o); P + P, pretreatment and challenge with Pam3CSK4. (B and C) ELISA of IL-6 and TNF-α in culture supernatants of murine macrophage RAW 264.7 cells stimulated for various times and doses with Pam3CSK4 (10 ng/mL) and challenged for 24h with Pam3CSK4 (10 ng/mL). (D) Cell viability was analyzed by WST-1 assay. The measurements of IL-6 and TNF- α were done in triplicate and are shown as mean±S.D.
*P < 0.05; **P < 0.01; ***P < 0.001 compared to - + P.
에 이식 (transfer)하였다. 이러한 membrane은 5% skim milk buffer로 blocking하였고, 다양한 1차 항체로 반응시킨 후 HRP- conjugated 2차 항체로 반응시켰다. 반응이 완료된 membrane은 WEST-ZOL plus western blot detection system (iNtRON Biotechnology, Korea)을 이용하여 발광시킨 후 X-ray 필름에 감 광시켜 현상하였다.
통계처리
본 연구의 통계처리는 Student’s t-test 분석을 이용하여 검증 하였고 값은 평균과 표준편차로 표현했다.
결과 및 고찰 (Results and discussion)
TLR1/2 배위자에 의한 면역관용 유도
염증반응은 활성화된 단핵세포나 대식세포가 분비한 사이토카 인 등에 의해 시작되는 것으로 알려져 있고 대표적인 염증성 사 이토카인인 IL-6와 TNF-α는 Th1 면역반응을 유도하여 자가면 역성 염증질환과 염증성 장질환 등을 일으킬 수 있다.29-31)따라 서, 마우스 대식세포에 TLR1/2 배위자인 Pam3CSK4의 반복 자 극에 의해 면역관용이 유도되는지 확인하였다. RAW 264.7 세포 에 1차 자극으로 Pam3CSK4 (1, 10, 50, 100 ng/mL)를 3, 6, 12, 24, 48시간 동안 처리하였고 다시 2차 자극으로 Pam3CSK4 (10 ng/mL)를 24시간 동안 처리 후 ELISA를 이용하여 사이토카 인 발현을 확인한 결과, IL-6와 TNF-α의 생성이 Pam3CSK4의 반복 자극에 의해 시간 및 농도 의존적으로 유의성 있게 감소되 는 것을 확인할 수 있었다(Fig. 1B, C). 이와 같이 Pam3CSK4로 유도된 면역관용이 RAW 264.7 세포에 대한 독성으로 인해 나 타나는 영향은 아닌지 확인하기 위하여 세포 생존능을 확인해 보 았다. RAW 264.7 세포에 1차 자극으로 Pam3CSK4 (1, 10, 20, 50, 100 ng/mL)를 24시간 동안 처리하였고 다시 2차 자극으로 Pam3CSK4 (10 ng/mL)를 24시간 동안 처리 후 WST-1 solution 을 이용하여 세포 생존능을 확인한 결과, Pam3CSK4의 다양한 농도의 반복 자극은 세포 생존능에 영향을 미치지 않았다(Fig.
1D). 이러한 결과들은 Pam3CSK4로 유도된 면역관용이 RAW 264.7 세포의 사멸에 의해 사이토카인 생성이 감소하여 나타나 는 것이 아니라 면역관용 발생에 의하여 나타나는 것임을 의미한다.
Pam3CSK4에 의한 면역관용 유도와 MAPK 신호전달의 상 관성
Pam3CSK4는 세포막에 존재하는 TLR1/2와 결합하여 Myeloid differentiation primary response 88 (MyD88)을 활성화시키고 이후 NF-κB, MAPKs와 같은 다양한 신호전달 경로를 활성화 시 킨다.32)활성화된 NF-κB와 MAPKs는 다양한 염증성 매개물질 의 합성을 촉진시키고 염증성 사이토카인 (IL-6, TNF-α, IL-1β
등)의 분비를 증가시켜 염증반응을 유도한다.33) 따라서, Pam3CSK4에 의한 면역관용 유도의 기전을 확인하기 위하여, RAW 264.7 세포에 1차 자극으로 Pam3CSK4 (10 ng/mL)를 24 시간 동안 처리하였고 다시 2차 자극으로 Pam3CSK4 (10 ng/
mL)를 제시된 시간 동안 처리 후 Western blot을 이용하여 NF- κB의 핵 내로 이동을 억제시키는 인자인 IκB와 MAPKs의 발현 을 확인하였다. 2차 Pam3CSK4만을 처리했을 때, IκB의 degradation이 시간 의존적으로 감소되었고 p38, JNK, ERK의 인산화가 15분에 가장 증가한 것을 확인하였다. 반면에 Pam3CSK4의 반복 자극은 감소되었던 IκB의 degradation을 회 복시켰고 p38, JNK, ERK의 인산화를 감소시켰다(Fig. 2A, B).
따라서 Pam3CSK4로 유도된 면역관용은 MAPKs와 NF-κB의 활 성화 억제를 통해 염증성 사이토카인의 생성을 억제시켜 면역관 용 발생을 나타내는 것임을 의미한다.
Pam3CSK4로 유도된 면역관용이 SOCS family와 A20의 활성화에 미치는 영향
이전 연구결과에서 SOCS1, SOCS3, A20 등의 신호 전달과정 을 억제하는 단백질의 증가가 MAPKs와 NF-?B를 불활성화 시 킨다는 보고가있었다.34)따라서, Pam3CSK4로 유도된 면역관용 이 SOCS family와 A20의 활성화에 미치는 영향을 확인하기 위 하여, RAW 264.7 세포에 1차 자극으로 Pam3CSK4 (10 ng/mL) 를 24시간 동안 처리하였고 다시 2차 자극으로 Pam3CSK4
Fig. 2 − Pretreatment with Pam3CSK4 suppresses TLR1/2 signaling.
(A and B) Immunoblot analysis of total I κBα and
phosphorylated (p-) p38, JNK and ERK in RAW 264.7 cells
stimulated for 24 h with Pam3CSK4 (10 ng/mL) and
challenged for various times with Pam3CSK4 (10 ng/mL).
(10 ng/mL)를 제시된 시간 동안 처리 후 Western blot을 이용하 여 SOCS1, SOCS3, A20의 발현을 확인하였다. Pam3CSK4를 반 복 자극한 결과, SOCS1, SOCS3, A20의 발현이 반복 자극을 하 지 않은 군과 비교하여 12, 24시간에 증가되었다(Fig. 3). 따라서 염증성 신호 전달과정을 억제하는 단백질인 SOCS1, SOCS3, A20의 활성화는 MAPKs와 NF-κB의 활성화를 억제함으로써 Pam3CSK4에 의한 면역관용 발생을 유도한다고 생각할 수 있다.
결 론 (Conclusion)
면역관용은 외부항원에 대한 면역반응을 억제하는 등의 다양 한 신호전달 기전이 존재하고 이식거부반응, 자가면역질환, 알레 르겐에 의한 면역질환 등의 치료법으로 제시될 수 있다.35,36)
이전 연구에 의하면 장상피세포 또는 대식세포에 TLR4의 배위 자인 lipopolysaccharide (LPS)를 반복적으로 노출시키면 다양한 염 증성 인자들과 TLR의 발현이 감소한다는 보고가 있었다.37,38)따 라서 본 연구에서는 RAW 264.7 세포에서 Pam3CSK4로 유도된 면역관용의 발생 기전을 연구하였다. Pam3CSK4의 반복 자극은 세포독성 없이 IL-6와 TNF-α의 생성을 농도 및 시간의존적으로 억제하였고 이러한 염증성 사이토카인의 생성을 유도하는 신호 전달 경로인 MAPKs의 불활성화 유도 및 IκB의 degradation을 억제함으로써 NF-κB가 핵 내로 이동하는 것을 저해하였다. 또 한 염증성 신호 전달과정을 억제하는 단백질인 SOCS1, SOCS3, A20을 활성화시킴으로써 Pam3CSK4에 의한 면역관용 발생을 유 도한다고 생각된다. 이러한 결과는 Pam3CSK4의 반복 자극은 면역관용을 유도하여 과도한 면역반응을 억제하고, 면역관용의 발생 기전에 대한 기초 자료로 사용될 수 있을 것이라 사료된다.
Conflict of Interests
본 논문에는 이해관계 충돌의 여지가 없음.
감사의 말씀 (Acknowledgments)
이 논문은 2017년도 제주대학교 연구교수 기간에 연구되었으 며, 한국연구재단의 지원을 받아 수행된 연구임(2016R1A2B4 014050).
References
1) Akira, S. and Takeda, K. : Toll-like receptor signalling. Nat Rev Immunol. 4, 499 (2004).
2) Gomez, P. F., Pillinger, M. H., Attur, M., Marjanovic, N., Dave, M., Park, J., Bingham, C. O., Al-Mussawir, H. and Abramson, S. B. : Resolution of inflammation: prostaglandin E2 dissociates nuclear trafficking of individual NF-kappaB subunits (p65, p50) in stimulated rheumatoid synovial fibroblasts. J Immunol. 175, 6924 (2005).
3) Jeong, D. H., Kim, K. B., Kim, M. J., Kang, B. K. and Ahn, D. H. : Anti-inflammatory activity of ethanolic extract of Sargassum micracanthum. J Microbiol Biotechnol. 23, 1691 (2013).
4) Pahan, K., Sheikh, F. G., Liu, X., Hilger, S., McKinney, M. and Petro, T. M. : Induction of nitric-oxide synthase and activation of NF-kappaB by interleukin-12 p40 in microglial cells. J Biol Chem. 276, 7899 (2001).
5) D'Acquisto, F., Iuvone, T., Rombola, L., Sautebin, L., Di-Rosa, M. and Carnuccio, R. : Involvement of NF-kappaB in the regulation of cyclooxygenase-2 protein expression in LPS- stimulated J774 macrophages. FEBS lett. 418, 175 (1997).
6) Makarov, S. S. : NF-kappa B in rheumatoid arthritis: a pivotal regulator of inflammation, hyperplasia, and tissue destruction.
Arthritis Res. 3, 200 (2001).
7) Chen, Z., Hagler, J., Palombella, V. J., Melandri, F., Scherer, D., Ballard, D. and Maniatis, T. : Signal-induced site-specific phosphorylation targets I kappa B alpha to the ubiquitin- proteasome pathway. Genes Dev. 9, 1586 (1995).
8) Vereecke, L., Beyaert, R. and van-Loo, G. : The ubiquitin- editing enzyme A20 (TNFAIP3) is a central regulator of immunopathology. Trends Immunol. 30, 383 (2009).
9) Thome, M. and Tschopp, J. : TCR-induced NF-kappaB activation:
a crucial role for Carma1, Bcl10 and MALT1. Trends Immunol.
24, 419 (2003).
10) Du, M. Q. : MALT lymphoma: many roads lead to nuclear factor-kappab activation. Histopathology. 58, 26 (2011).
11) Boone, D. L., Turer, E. E., Lee, E. G., Ahmad, R. C., Wheeler, M. T., Tsui, C., Hurley, P., Chien, M., Chai, S., Hitotsumatsu, O., McNally, E., Pickart, C. and Ma, A. : The ubiquitin- modifying enzyme A20 is required for termination of Toll-like receptor responses. Nat Immunol. 5, 1052 (2004).
12) Chanudet, E., Ye, H., Ferry, J., Bacon, C. M., Adam, P., Muller- Hermelink, H. K., Radford, J., Pileri, S. A., Ichimura, K.,
Fig. 3 − Pretreatment with Pam3CSK4 induces SOCS1, SOCS3 and
A20 expression. Immunoblot analysis of SOCS1, SOCS3
and A20 in RAW 264.7 cells stimulated for 24h with
Pam3CSK4 (10 ng/mL) and challenged for various times
with Pam3CSK4 (10 ng/mL).
Collins, V. P., Hamoudi, R. A., Nicholson, A. G., Wotherspoon, A. C., Isaacson, P. G. and Du, M. Q. : A20 deletion is associated with copy number gain at the TNFA/B/C locus and occurs preferentially in translocation-negative MALT lymphoma of the ocular adnexa and salivary glands. J Pathol. 217, 420 (2009).
13) Chanudet, E., Huang, Y., Ichimura, K., Dong, G., Hamoudi, R.
A., Radford, J., Wotherspoon, A. C., Isaacson, P. G., Ferry, J.
and Du, M. Q. : A20 is targeted by promoter methylation, deletion and inactivating mutation in MALT lymphoma.
Leukemia. 24, 483 (2010).
14) Plenge, R. M., Cotsapas, C., Davies, L., Price, A. L., de-Bakker, P. I., Maller, J., Pe'er, I., Burtt, N. P., Blumenstiel, B., DeFelice, M., Parkin, M., Barry, R., Winslow, W., Healy, C., Graham. R.
R., Neale, B. M., Izmailova, E., Roubenoff, R., Parker, A. N., Glass, R., Karlson, E. W., Maher, N., Hafler, D. A., Lee, D. M., Seldin, M. F., Remmers, E. F., Lee, A. T., Padyukov, L., Alfredsson, L., Coblyn, J., Weinblatt, M. E., Gabriel, S. B., Purcell, S., Klareskog, L., Gregersen, P. K., Shadick, N. A., Daly, M. J. and Altshuler, D. : Two independent alleles at 6q23 associated with risk of rheumatoid arthritis. Nat Genet. 39, 1477 (2007).
15) Egwuagu, C. E., Yu, C. R., Zhang, M., Mahdi, R. M., Kim, S.,J.
and Gery, I. : Suppressors of cytokine signaling proteins are differentially expressed in Th1 and Th2 cells: implications for Th cell lineage commitment and maintenance. J Immunol.
168, 3181 (2002).
16) Krebs, D. L. and Hilton, D. J. : SOCS: physiological suppressors of cytokine signaling. J Cell Sci. 113, 2813 (2000).
17) Ito, S., Ansari, P., Sakatsume, M., Dickensheets, H., Vazquez, N., Donnelly, R. P., Larner, A. C. and Finbloom, D. S. : Interleukin-10 inhibits expression of both interferon alpha- and interferon gamma- induced genes by suppressing tyrosine phosphorylation of STAT1. Blood. 93, 1456 (1999).
18) Naka, T., Narazaki, M., Hirata, M., Matsumoto, T., Minamoto, S., Aono, A., Nishimoto, N., Kajita, T., Taga, T., Yoshizaki, K., Akira, S. and Kishimoto, T. : Structure and function of a new STAT-induced STAT inhibitor. Nature. 387, 924 (1997).
19) Endo, T. A., Masuhara, M., Yokouchi, M., Suzuki, R., Sakamoto, H., Mitsui, K., Matsumoto, A., Tanimura, S., Ohtsubo, M., Misawa, H., Miyazaki, T., Leonor, N., Taniguchi, T., Fujita, T., Kanakura, Y., Komiya, S. and Yoshimura, A. : A new protein containing an SH2 domain that inhibits JAK kinases. Nature. 387, 921 (1997).
20) Yasukawa, H., Misawa, H., Sakamoto, H., Masuhara, M., Sasaki, A., Wakioka, T., Ohtsuka, S., Imaizumi, T., Matsuda, T., Ihle, J. N. and Yoshimura, A. : The JAK-binding protein JAB inhibits Janus tyrosine kinase activity through binding in the activation loop. EMBO j. 18, 1309 (1999).
21) Ernst, M., Inglese, M., Waring, P., Campbell, I. K., Bao, S., Clay, F. J., Alexander, W. S., Wicks, I. P., Tarlinton, D. M., Novak, U., Heath, J. K. and Dunn, A. R. : Defective gp130- mediated signal transducer and activator of transcription (STAT) signaling results in degenerative joint disease, gastrointestinal ulceration, and failure of uterine implantation.
J Exp Med. 194, 189 (2001).
22) Alexander, W. S., Starr, R., Fenner, J. E., Scott, C. L., Handman, E., Sprigg, N. S., Corbin, J. E., Cornish, A. L., Darwiche, R., Owczarek, C. M., Kay, T. W., Nicola, N. A., Hertzog, P. J., Metcalf, D. and Hilton, D. J. : SOCS1 is a critical inhibitor of interferon gamma signaling and prevents the potentially fatal neonatal actions of this cytokine. Cell. 98, 597 (1999).
23) Marine, J. C., Topham, D. J., McKay, C., Wang, D., Parganas, E., Stravopodis, D., Yoshimura, A. and Ihle, J. N. : SOCS1 deficiency causes a lymphocyte-dependent perinatal lethality.
Cell. 98, 609 (1999).
24) Berlato, C., Cassatella, M. A., Kinjyo, I., Gatto, L., Yoshimura, A. and Bazzoni, F. : Involvement of suppressor of cytokine signaling-3 as a mediator of the inhibitory effects of IL-10 on lipopolysaccharide-induced macrophage activation. J Immunol.
168, 6404 (2002).
25) Shouda, T., Yoshida, T., Hanada, T., Wakioka, T., Oishi, M., Miyoshi, K., Komiya, S., Kosai, K., Hanakawa, Y., Hashimoto, K., Nagata, K. and Yoshimura, A. : Induction of the cytokine signal regulator SOCS3/CIS3 as a therapeutic strategy for treating inflammatory arthritis. J Clin Invest. 108, 1781 (2001).
26) Suzuki, A., Hanada, T., Mitsuyama, K., Yoshida, T., Kamizono, S., Hoshino, T., Kubo, M., Yamashita, A., Okabe, M., Takeda, K., Akira, S., Matsumoto, S., Toyonaga, A., Sata, M. and Yoshimura, A. : CIS3/SOCS3/SSI3 plays a negative regulatory role in STAT3 activation and intestinal inflammation. J Exp Med. 193, 471 (2001).
27) Hogquist, K. A., Baldwin, T. A. and Jameson, S. C. : Central tolerance: learning self-control in the thymus. Nat Rev Immunol. 5, 772 (2005).
28) Walker, L. S. and Abbas, A. K. : The enemy within: keeping self-reactive T cells at bay in the periphery. Nat Rev Immunol.
2, 11 (2002).
29) Umetsu, D. T. and DeKruyff, R. H. : Th1 and Th2 CD4+ cells in the pathogenesis of allergic diseases. Pro Soc Exp Biol Med.
215, 11 (1997).
30) Underhill, D. M., Ozinsky, A., Smith, K. D. and Aderem, A. : Toll- like receptor-2 mediates mycobacteria-induced proinflammatory signaling in macrophages. Pro Nati Acad Sci U S A. 96, 14459 (1999).
31) Leal, I. S., Florido, M., Andersen, P. and Appelberg, R. : Interleukin-6 regulates the phenotype of the immune response to a tuberculosis subunit vaccine. Immunology. 103, 375
(2001).
32) Rhee, S. H. and Hwang, D. : Murine TOLL-like receptor 4 confers lipopolysaccharide responsiveness as determined by activation of NF kappa B and expression of the inducible cyclooxygenase. J Biol Chem. 275, 34035 (2000).
33) Waetzig, V., Czeloth, K., Hidding, U., Mielke, K., Kanzow, M., Brecht, S., Goetz, M., Lucius, R., Herdegen, T. and Hanisch, U. K. : c-Jun N-terminal kinases (JNKs) mediate pro- inflammatory actions of microglia. Glia. 50, 235 (2005).
34) Park, S. H., Park-Min, K. H., Chen, J., Hu, X. and Ivashkiv, L. B. : Tumor necrosis factor induces GSK3 kinase-mediated cross- tolerance to endotoxin in macrophages. Nat Immunol. 12, 607 (2011).
35) Marrack, P., Kappler, J. and Kotzin, B. L. : Autoimmune
disease: why and where it occurs. Nat Med. 7, 899 (2001).
36) Calderon, M. A., Casale, T. B., Togias, A., Bousquet, J., Durham, S. R. and Demoly, P. : Allergen-specific immunotherapy for respiratory allergies: from meta-analysis to registration and beyond. J Allergy Clin Immunol. 127, 30 (2011).
37) Otte, J. M., Cario, E. and Podolsky, D. K. : Mechanisms of cross hyporesponsiveness to Toll-like receptor bacterial ligands in intestinal epithelial cells. Gastroenterology. 126, 1054 (2004).
38) Lee, S. K., Kim, T., Kim, Y. K., Choi, C. H., Yang, K. M., Chae, B. and Kim, W. H. : Cellular differentiation-induced attenuation of LPS response in HT-29 cells is related to the down- regulation of TLR4 expression. Biochem Biophys Res Commun.
18, 337 (2005).