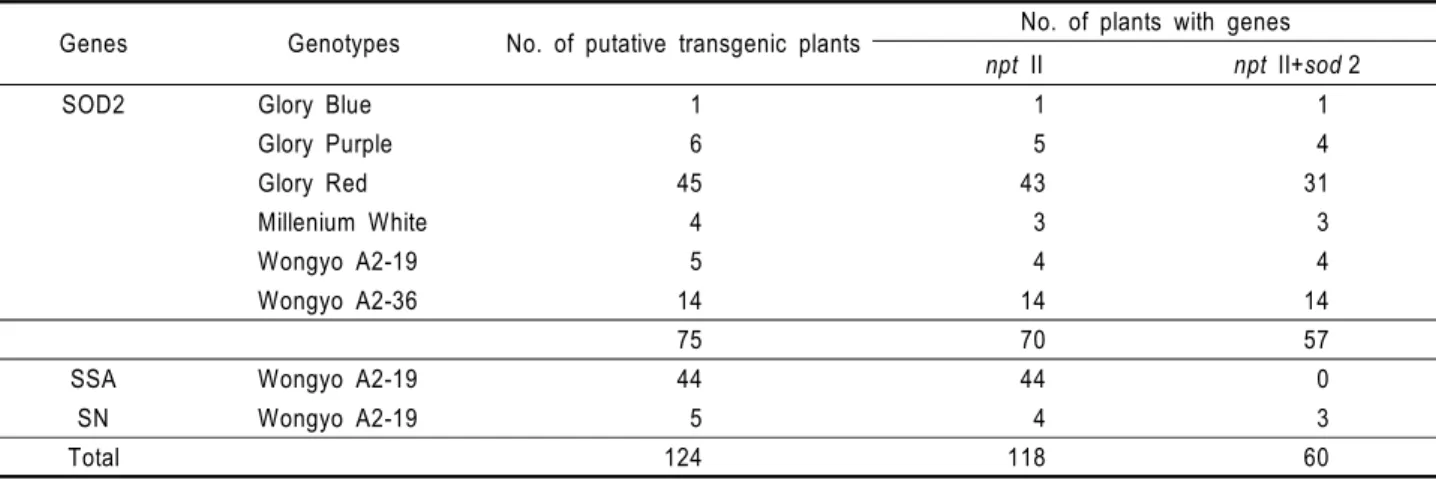
Transfer of SOD2 or NDP kinase 2 genes into purebred lines of petunia
Su Young Lee
1*・ Bong Hee Han
1・ Eun Woon Noh
2・ Sang-Soo Kwak
31
Floricultural Research Division, National Institute of Horticultural & Herbal Science, Rural Development Administration, Suwon 441-310, Korea
2
Department of Forest Genetic Resources, Korea Forest Research Institute, Suwon 441-350, Korea
3