평야지 적응성 향상을 위한 벼흰잎마름병 및 줄무늬잎마름병 저항성 유전자 집적 조생 계통 개발
박현수1* ・ 남정권1・ 김기영1・ 김우재1・ 정지웅1・ 백만기1・ 김정주1・ 조영찬1・ 이점호2・ 김보경1・ 안상낙3
1농촌진흥청 국립식량과학원, 2농촌진흥청 국립식량과학원 중부작물부, 3충남대학교 농업생명대학
Development of Early Maturing Rice Lines with Genes Conferring Resistance to Bacterial Blight and Rice Stripe Virus for Enhancing the
Adaptability in Plain Area
Hyun-Su Park
1*, Jeong-Kwon Nam
1, Ki-Young Kim
1, Woo-Jae Kim
1, Ji-Ung Jeong
1, Man-Kee Baek
1, Jeong-Ju Kim
1, Young-Chan Cho
1, Jeom-Ho Lee
2, Bo-Kyeong Kim
1, and Sang-Nag Ahn
31
National Institute of Crop Science, RDA, Wanju 565-851, Korea
2
Department of Central Area, NICS, RDA, Suweon 441-707, Korea
3
Department of Agronomy, Chungnam National University, Daejeon 305-764, Korea
Abstract : This study was conducted to develop the early maturing rice lines with genes conferring resistance to bacterial blight and rice stripe virus to enhance the adaptability in plain area. Unkwang carrying Xa3 was used as a recurrent parent and SR30075 carrying Xa4+xa5+Xa21+Stvb-i was used as a donor parent. RL1(Resistant Line, BC
1F
7), RL2, RL3, RL4, and RL5(BC
2F
6) were bred through bio-assay of K3a race inoculation and phenotypic selection of agronomic traits. The presence of introduced genes was confirmed by testing the resistance levels against bacterial blight and rice stripe virus and then double-checked by using DNA marker. RL1 has all target genes, Xa3+xa5+Xa21+Stvb-i. RL2, RL3, and RL5 have Xa3+Xa21+Stvb-i whereas RL4 has only Xa21. Rice lines carrying Stvb-i showed resistance reaction to rice stripe virus. The combinations of bacterial blight resistant genes(Xa3+xa5+Xa21 and Xa3+Xa21) were found to be promising, as the rice lines carrying these genes enhanced a strong resistant reaction against 16 bacterial blight isolates. Also, the inoculation of K3a race did not alter the brown rice yield, ripened grain ratio and kernel quality of brown rice compared to control. Although RL1 containing all the target resistance genes showed excellent resistance performance, it is not suitable to cultivate in plain area due to instability to lodging, 80% yield level than Unkwang, and low grain quality. RL5 backcrossed twice with Unkwang was found to be a promising line due to its effective resistance gene combination, Xa3+Xa21+Stvb-i and good agronomic traits such as stability to lodging, higher yield and quality compared to Unkwang.
Keywords : Rice, Early maturing, Bacterial blight, Rice stripe virus, Resistance gene
*Corresponding author (E-mail: [email protected], Tel: +82-63-840- 2256, Fax: +82-63-840-2119)
(Received on May 6, 2015. Revised on May 20, 2015.
Accepted on May 28, 2015.)
118
http://dx.doi.org/10.9787/KJBS.2015.47.2.118 Print ISSN: 0250-3360
Copyright ⓒ 2015 by the Korean Society of Breeding Science
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
서 언
우리나라에서 벼는 생육시기와 출수기에 따라 조생종, 중 생종 및 중만생종으로 구별한다(Kim 2010). 조생종 벼는 중 생종이나 중만생종 벼에 비해 생육 기간이 짧아 일찍 수확할
수 있으며, 저온에 대한 내성이 강하여 대체로 기온이 낮은 산간 지방을 중심으로 재배되고 있다. 또한 조생종 벼는 일반 적으로 미질과 생산성이 낮아 중생종이나 중만생종 벼에 비 해 재배선호도가 낮았다(Ha et al. 2003). 하지만 최근 평야지 에서 추석 전 햅쌀 출하, 이모작 등 작부체계의 다양화, 수확 시기의 다변화를 통한 노동력 분산을 위해 조생종 벼 면적이 크게 증가하고 있는 추세여서 이에 적응한 조생종 벼 품종 개 발이 요구되는 실정이다.
평야지는 중산간지에 비해 병해충 발생이 많아 평야지에
적응하는 조생종 벼 품종을 개발하기 위해서는 병해충에 대 한 저항성 향상이 필요하다. 특히, 조생종 벼는 중생종과 중만 생종 벼에 비해 벼흰잎마름병 및 줄무늬잎마름병에 대한 저 항성이 취약하다. 우리나라 자포니카 타입 중 벼흰잎마름병에 대한 저항성에 대한 연구결과 조생종, 중생종, 중만생종의 저 항성 품종 비율이 각각 6, 50, 60%로 조생종이 중생, 중만생 종 벼에 비해 낮았다(Park et al. 2011). 벼 저항성 품종 육종 시 문제시되고 있는 K3a균계에 대한 저항성 품종은 중만생종 으로 진백(2008년 개발), 안백, 만백, 새신(2014) 등 4 품종이 개발되었고 중생종으로 강백(2006), 신백(2010), 해품(2013) 등 3 품종이 개발되었으나 조생종으로 개발된 품종은 전무한 실정이다(Kim et al. 2009, Park et al. 2013, Shin et al.
2011). 또한 2014년까지 농촌진흥청에서 개발된 329개 품종 중 줄무늬잎마름병에 저항성인 품종 비율이 조생종 20.9, 중 생종 62.0, 중만생종 71.8%로 조생종이 줄무늬잎마름병에 취 약하여 병 발생에 따른 피해가 우려되고 있다.
벼의 병해충에 대한 피해를 가장 적극적으로 대처하는 방 법은 저항성 품종을 개발하고 이를 재배하는 것이다(Khush 1989). 저항성 품종 개발을 위한 육종사업은 효과적인 저항성 원을 확보하고 이를 우량 품종으로 도입하는 일련의 과정을 거 친다. 우리나라는 벼흰잎마름병 저항성 육종사업을 지속적으로 수행하여 왔다. 초기에는 우리나라 우점 균계였던 K1에 저항성 인 Xa1 유전자를 이용하였고, 이후 K1, K2 및 K3 균계에 저항 성인 Xa3 유전자를 이용하여 많은 저항성 품종이 개발되었다.
Xa3 유전자를 단일 저항성원으로 하여 개발된 우수한 품종들의
대면적 재배가 장기화됨에 따라서 병원균 레이스 집단의 변화 와 새로운 변이균이 발생하여 피해가 확산되었다(Park et al.2013). 최근 벼흰잎마름병 저항성 유전자의 다양화를 위해 방 글라데시 품종인 DV85에서 xa5 유전자가 도입된 계통이 실질 적인 품종 개발로 이루어져 강백이 육성되었고, Xa3 와 xa5 유 전자가 결합된 진백이 육성되어 변이균인 K3a에 대응할 방안을 마련하였다(Shin et al. 2011). 또한 중생종인 수원345호와 조 생찰벼인 상주찰벼 배경에 벼흰잎마름병 저항성 유전자가 도입 된 근동질계통을 개발하여 육종사업에 활용하고 있으며 저항성 유전자를 표지할 수 있는 분자표지를 개발하여 세 개의 저항성 유전자 Xa3+xa5+Xa21 조합의 익산575호 등 다양한 저항성 유 전자가 집적된 저항성 계통을 육성하고 있다(Kim et al. 2011, Park et al. 2013, Park et al. 2014a). 우리나라 자포니카 벼 품 종의 줄무늬잎마름병 저항성은 인디카 Modan 유래 저항성 유 전자 Stvb-i가 유래되어 안정적으로 이용되고 있다(Kwon et al.
2012). 1975년에 최초의 저항성 품종인 낙동벼가 개발(Chung et al. 1975)된 이후로 저항성 품종개발에 Stvb-i 유전자가 지속적 으로 활용되고 있으나 Stvb-i 유전자의 저항성 붕괴에 따른 문 제는 보고되고 있지 않다(Park et al. 2014b).
평야지에서 조생종 벼 품종 재배가 증가하고 있으나 우리 나라에서 육성된 조생종 110개 품종 중 벼흰잎마름병에 저항 성인 품종은 Xa3 유전자를 단독으로 가지고 있는 것으로 추 정되는 운광(2004년 개발), 금오3호(2005), 주남조생(2006), 운미(2007), 조평(2010), 산호미(2012), 화왕(2012), 운일찰 (2014), 해담쌀(2014) 등 9개 품종 뿐이고, Stvb-i 유전자를 가지고 있는 저항성 품종의 비율이 20.9%로 평야지 재배시 벼흰잎마름병 및 줄무늬잎마름병 발생으로 피해가 우려되고 있다. 본 연구는 조생종 벼의 평야지 적응성을 향상시키고자 벼흰잎마름병 및 줄무늬잎마름병에 대한 저항성 유전자가 집 적된 저항성 계통을 개발하고 이들의 저항성 성능과 수량성 등 농업형질을 분석하여 육종사업에 반영하고자 수행하였다.
재료 및 방법
시험재료 및 재배방법
조생종 벼 품종인 운광과 조평, 그리고 운광을 반복친으로 하 여 육성된 5개 저항성 계통을 시험에 이용하였다. 공시 재료를 국립식량과학원 익산 벼 시험포장에서 2014년 4월 10일 파종 하여 5월 10일 재식거리 30 x 15 cm로 주당 3본씩 구당 180주 를 3반복으로 이앙하여 생산력 검정시험을 수행하였다. 벼흰잎 마름병 접종에 따른 수량성 및 품질 변이를 조사하기 위해 5월 3일 파종하여 6월 1일에 무접종구와 접종구를 재식거리 30 x 15 cm로 주당 1본씩 구당 60주를 3반복으로 나누어 이앙하였 다. 벼흰잎마름병 대표균계에 대한 저항성 반응을 조사하기 위 해 재식거리 30 x 15 cm로 주당 1본씩 30주를 이앙하였고, 광 범위 저항성 반응을 조사하기 위해 플라스틱 상자(78 × 45 × 18 cm)에 품종 및 계통을 주당 3본씩 16주 이앙하였다. 시비량 은 N-P2O5-K2O를 90-45-57 kg/ha으로 질소는 기비 : 분얼비 : 수비를 50 : 20 : 30 비율로 분시하였고, 인산은 전량 기비로, 칼륨은 기비 : 수비를 70 : 30 비율로 분시하였다. 기타 재배관 리는 농촌진흥청 표준 재배법에 준하여 실시하였다.
벼흰잎마름병 및 줄무늬잎마름병 저항성 검정
벼흰잎마름병 접종에 따른 수량성 및 품질 변이를 조사하 기 위해 무접종구는 균을 묻히지 않고, 접종구는 병원성이 강
한 HB1009 (K3a균계) 균주를 이용하여 시험구 전개체에 대 해서 최고분얼기에 엽선단 3 cm 부위를 가위 절엽접종하였다 (Kauffman et al. 1973). 벼흰잎마름병 대표균계에 대한 저항 성 반응은 HB1013 (K1 균계), HB1014 (K2), HB1015 (K3), HB1009 (K3a) 균주를 이용하여 최고분얼기에 균주 별로 3 주씩 접종하였다. 접종 후 3주 후에 각각의 개체에서 가장 긴 병반을 가진 3개 엽의 병반장을 측정하여 평균 병반장 길이가 5 cm 이하는 저항성(R; resistant), 5-10 cm는 중도저항성(MR;
moderately resistant), 10 cm 이상은 이병성(S; susceptible) 으로 질적저항성을 구분하였다. 벼흰잎마름병에 대한 광범위 저항성 검정을 위해서 우라나라의 우점 균주인 HB1014(K2 균계), HB1015 (K3), HB1009 (K3a)와 Xa3 유전자를 침해 하는 것으로 알려진 HB2024, HB2038, HB3055, HB3079 (K4), HB4024, HB4040, HB4044 (K5), HB4079, HB6142, HB6159와 Xa3 유전자를 침해하지 못하나 Xa21 유전자를 침해하는 HB2010, HB3011, HB4027 균주(Park et al. 2013) 등 총 16개 균주를 이용하였다. 플라스틱 상자에 이앙된 16 개체에 한 개체 당 한 균주씩 엽선단 약 3 cm 부위를 가위 절엽접종하여 접종 3주 후에 가장 긴 병반을 가진 3개 엽의 병반장을 측정하여 평균한 값을 이용하였다. 질적 저항성은 대표균계에 대한 저항성 반응 기준에 추가로 병반장 1 cm 이 하는 고도저항성(HR; highly resistant)으로 구분하였다.
벼줄무늬잎마름병 검정은 망실을 이용한 대량 검정법으로 바이러스 보독충의 방사 및 계대 사육으로 보독충이 충분히 유지된 망실에서 계통당 1.5 g을 조파하였으며, 파종 후 30일 경 이병성 대비품종 일품벼가 병징을 나타낼 때 저항성과 감 수성으로 판정하였다(Kwak et al. 2007).
저항성 유전자 확인
Genomic DNA 추출은 BioSprint 96 (Qiagen Co., Düren, Germany)을 이용하였다. 샘플을 TissueLyserⅡ (Qiagen Co., Düren, Germany)를 이용하여 마쇄한 후 BioSprint 96 DNA Plant Kit (Qiagen Co., Düren, Germany)를 이용하여 DNA 를 추출하였다. 저항성 유전자 Xa3, xa5, Xa21 및 Stvb-i를 확인하기 위해 대상 유전자와 밀접하게 연관된 DNA 분자 표 지인 9643.T4, 10603.T10Dw, U1/I1 및 InDel 7을 각각 이 용하였다(Kwon et al. 2012, Park et al. 2013, Wang et al.
1996). PCR은 10 ng의 DNA와 AccuPower® PCR PreMix (Bioneer Co., Daejoen, Korea)를 이용하여 My-Genie 96 Thermal block (Bioneer Co., Daejoen, Korea)에서 수행하
였다. PCR 반응은 9643.T4는 94℃에서 5분간 초기변성 후 94℃ 40초, 63℃ 40초, 72℃ 1분간 총 40회 반복하고, 72℃
에서 5분간 반응하였다. 10603.T10Dw과 U1/I1은 95℃에서 4분간 초기변성 후 95℃ 30초, 65℃ 또는 56℃ (U1/I1) 30 초, 72℃ 1분간 총 35회 반복하고, 72℃에서 10분간 반응하였 다. InDel 7은 94℃에서 5분간 초기변성 후 94℃ 30초, 58℃
30초, 72℃ 1분간 총 45회 반복하고, 72℃에서 5분간 반응하 였다. 증폭된 PCR 산물 4 ㎕를 9643.T4는 2U Taqα Ⅰ와 6 5℃에서 3시간, 10603.T10Dw는 Rsa Ⅰ와 37℃에서 3시간 제한효소 처리하였다. U1/I1과 InDel 7 의한 증폭 산물은 제한 효소 처리 없이 전기영동에 이용하였다. 2% agarose gel에서 전기영동을 실시하여 EtBr로 염색한 후 UV transilluminator (MiniBIS Pro, DNR Bio-Imaging Systems Ltd., Jerusalem, Israel)를 이용하여 유전자형을 판정하였다.
수량 및 품질 관련 형질 조사
벼흰잎마름병 접종에 따른 수량성 및 품질 변이를 조사하 기 위해 출수 후 40일에 접종구와 대조구에서 3주를 예취하 여 등숙률을 측정하고, 20주를 예취하여 정조수량을 측정하 였다. 제현기(SY88-TH, Ssangyong Ltd., Incheon, Korea)로 제영하여 현미수량을 구하고 RN300 (Kett Co., Ltd., Tokyo, Japan)을 이용하여 현미 외관품위를 조사하였다. 생산력 검정 시험에 공시된 재료의 출수기를 조사하고 성숙기에 시험구별 로 10개체에 대해서 간장, 수장, 수수를 측정하였다. 출수 후 40일에 시험구별로 3주를 예취하여 등숙률 및 수당립수를 측 정하고, 수량조사는 100주를 예취하여 정조수량을 측정한 다 음 이 중 1.5 kg에 대해서 수량조사현미기(LST, Gwangyang, Korea)로 제영하여 정현비율을 측정하고, 100주 정조수량에 정현비율을 곱하여 현미수량을 구하고 10a당 수량으로 환산 하였다. 제현된 현미를 가지고 현미천립중을 측정하였다. 현 미를 시험용 정미기(VP-32T, Yamamoto Co., Ltd., Yamagata, Japan)를 이용하여 도정 후 RN300 (Kett Co., Ltd., Tokyo, Japan)을 이용하여 백미에 대한 외관품위를 조사하였다. 백미 의 단백질 및 아밀로스 함량은 Infratec 1241 Grain Analyzer (Foss Tecator, Hoganas, Sweden)을 이용하였고, 취반미의 윤기치는 식미검정기인 MA-90B (Toyo Co., Tokyo, Japan) 을 이용하여 측정하였다.
통계 분석
통계분석은 SAS 프로그램(Version 9.2, SAS Institute Inc.,
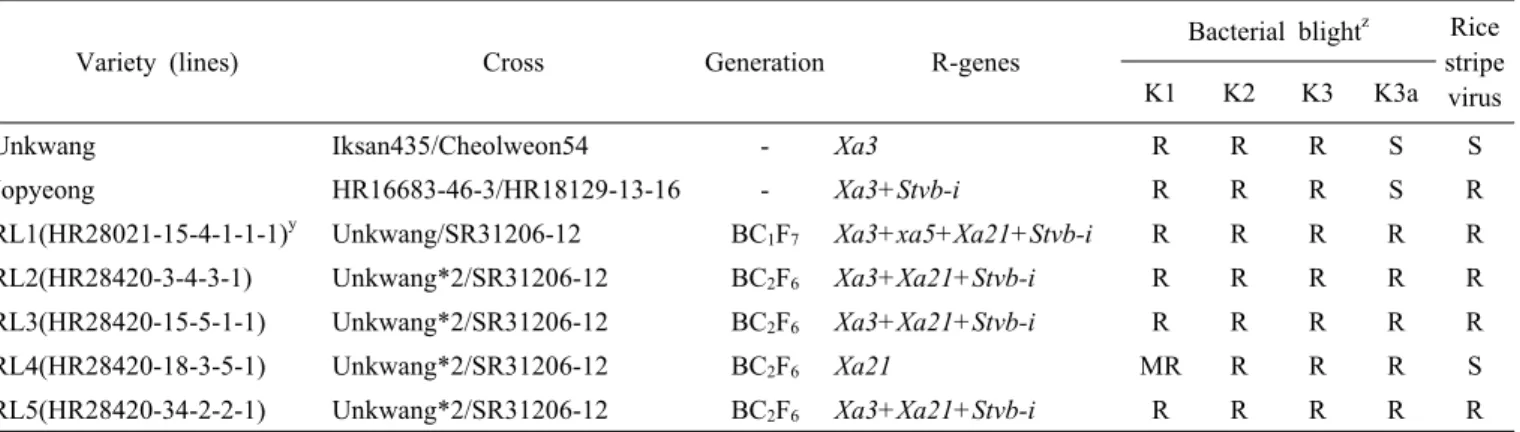
Fig. 1. Pedigree diagram of breeding lines.
Table 1. Pedigree and resistance genes of varieties and breeding lines and their resistance reaction to bacterial blight and rice stripe virus by bio-assay.
Variety (lines) Cross Generation R-genes Bacterial blight
zRice
stripe virus
K1 K2 K3 K3a
Unkwang Iksan435/Cheolweon54 - Xa3 R R R S S
Jopyeong HR16683-46-3/HR18129-13-16 - Xa3+Stvb-i R R R S R
RL1(HR28021-15-4-1-1-1)
yUnkwang/SR31206-12 BC
1F
7Xa3+xa5+Xa21+Stvb-i R R R R R
RL2(HR28420-3-4-3-1) Unkwang*2/SR31206-12 BC
2F
6Xa3+Xa21+Stvb-i R R R R R
RL3(HR28420-15-5-1-1) Unkwang*2/SR31206-12 BC
2F
6Xa3+Xa21+Stvb-i R R R R R
RL4(HR28420-18-3-5-1) Unkwang*2/SR31206-12 BC
2F
6Xa21 MR R R R S
RL5(HR28420-34-2-2-1) Unkwang*2/SR31206-12 BC
2F
6Xa3+Xa21+Stvb-i R R R R R
z
R, MR, and S mean resistant, moderately resistant, and susceptible to disease, respectively
y
RL means resistant line and notation in parenthesis is pedigree number
Cary, NC, USA)을 이용하였다. 농업형질에 대한 평균을 기술 통계법으로 구하였고, 평균간 비교는 PROC ANOVA로 분산분석 후 유의성이 있을 경우 5% 유의수준에서 Duncan’s Multiple Range test (DMRT)로 분석하였으며, PROC t-test 를 이용하여 균계 접종에 따른 대조구와 접종구간의 농업형 질 차이를 비교하였다.
결 과
저항성 계통 육성 및 저항성 유전자 확인
벼흰잎마름병과 줄무늬잎마름병에 저항성인 조생종 벼 품
종을 개발하기 위해서 조생종 품종으로 Xa3유전자를 가지고 있는 운광을 모본으로 하고 중만생종이며 벼흰잎마름병 저항 성 유전자 Xa4+xa5+Xa21과 줄무늬잎마름병 저항성 유전자
Stvb-i을 가지고 있는 SR30075조합의 계통을 수여친으로 이
용하였다. F2 분리세대에 저항성 유전자 표지 분자마커를 이용 하여Xa3는 이형접합체이고 xa5, Xa21, Stvb-i는 동형접합체
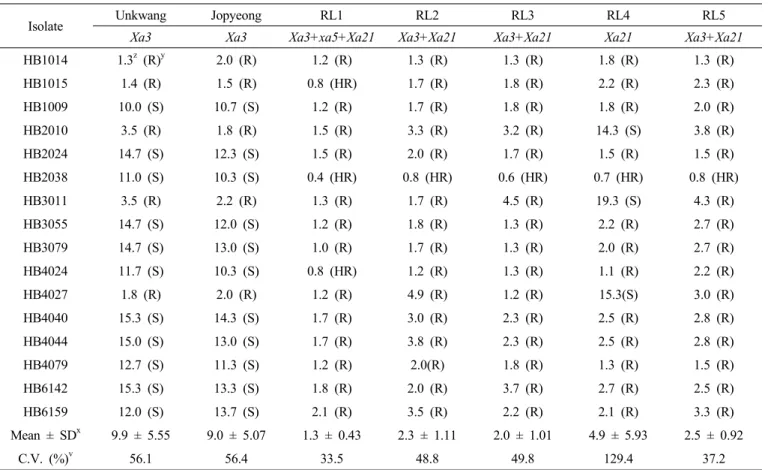
인 F3계통을 선발하였고 이를 운광에 여교배하였다(Fig. 1). 1 회 여교배한 HR28021과 2회 여교배한 HR28420 조합에 대해 서 벼흰잎마름병 K3a균계에 대한 생물검정과 농업형질 조사 를 통해 BC1F7 세대 HR280021-15-4-1-1-1 (이하 RL1 표기) 과 BC2F6 세대 HR28420-3-4-3-1 (RL2), HR28420-15-5-1-1Fig. 2. PCR analysis of lines to confirm resistance genes using the gene specific DNA marker. Xa3, xa5, Xa21, and Stvb-i was confirmed by PCR product amplified with primer, 9643.T4 (Taq α I, A), 10603.T10Dw (cleaved by Rsa I, B), U1/I1 (C), and InDel 7 (D), respectively. The white stars represent the lines carrying resistance genes.
M: DNA size marker, 1: IR24, 2: IRBB3, 3: IRBB5, 4:
IRBB21, 5: Unkwang, 6: Jopyeong, 7: RL1, 8: RL2, 9:
RL3, 10: RL4, 11: RL5.
Table 2. Resistance reaction of varieties and breeding lines against each of 16 Korean Xanthomonas oryzae pv. oryzae.
Isolate Unkwang Jopyeong RL1 RL2 RL3 RL4 RL5
Xa3 Xa3 Xa3+xa5+Xa21 Xa3+Xa21 Xa3+Xa21 Xa21 Xa3+Xa21
HB1014 1.3
z(R)
y2.0 (R) 1.2 (R) 1.3 (R) 1.3 (R) 1.8 (R) 1.3 (R)
HB1015 1.4 (R) 1.5 (R) 0.8 (HR) 1.7 (R) 1.8 (R) 2.2 (R) 2.3 (R)
HB1009 10.0 (S) 10.7 (S) 1.2 (R) 1.7 (R) 1.8 (R) 1.8 (R) 2.0 (R)
HB2010 3.5 (R) 1.8 (R) 1.5 (R) 3.3 (R) 3.2 (R) 14.3 (S) 3.8 (R)
HB2024 14.7 (S) 12.3 (S) 1.5 (R) 2.0 (R) 1.7 (R) 1.5 (R) 1.5 (R)
HB2038 11.0 (S) 10.3 (S) 0.4 (HR) 0.8 (HR) 0.6 (HR) 0.7 (HR) 0.8 (HR)
HB3011 3.5 (R) 2.2 (R) 1.3 (R) 1.7 (R) 4.5 (R) 19.3 (S) 4.3 (R)
HB3055 14.7 (S) 12.0 (S) 1.2 (R) 1.8 (R) 1.3 (R) 2.2 (R) 2.7 (R)
HB3079 14.7 (S) 13.0 (S) 1.0 (R) 1.7 (R) 1.3 (R) 2.0 (R) 2.7 (R)
HB4024 11.7 (S) 10.3 (S) 0.8 (HR) 1.2 (R) 1.3 (R) 1.1 (R) 2.2 (R)
HB4027 1.8 (R) 2.0 (R) 1.2 (R) 4.9 (R) 1.2 (R) 15.3(S) 3.0 (R)
HB4040 15.3 (S) 14.3 (S) 1.7 (R) 3.0 (R) 2.3 (R) 2.5 (R) 2.8 (R)
HB4044 15.0 (S) 13.0 (S) 1.7 (R) 3.8 (R) 2.3 (R) 2.5 (R) 2.8 (R)
HB4079 12.7 (S) 11.3 (S) 1.2 (R) 2.0(R) 1.8 (R) 1.3 (R) 1.5 (R)
HB6142 15.3 (S) 13.3 (S) 1.8 (R) 2.0 (R) 3.7 (R) 2.7 (R) 2.5 (R)
HB6159 12.0 (S) 13.7 (S) 2.1 (R) 3.5 (R) 2.2 (R) 2.1 (R) 3.3 (R)
Mean ± SD
x9.9 ± 5.55 9.0 ± 5.07 1.3 ± 0.43 2.3 ± 1.11 2.0 ± 1.01 4.9 ± 5.93 2.5 ± 0.92
C.V. (%)
v56.1 56.4 33.5 48.8 49.8 129.4 37.2
z
Average lesion length was measured using 3 leaves mostly inoculated
y
HR: Highly resistant (< 1 cm lesion length) R: Resistant (1-5 cm), MR: Moderately resistant (5-10 cm), S: Susceptible (> 10 cm)
x
SD: Standard deviation,
vC.V.: Coefficient of variation
(RL3), HR28420-18-3-5-1 (RL4), HR28420-34-2-2-1 (RL5) 등 총 5개 우량계통을 선발하였다(Table 1). 저항성유전자를 표지하는 분자마커를 이용하여 이들 계통들이 보유한 저항성 유전자를 확인한 결과 운광은
Xa3, 조평은 Xa3와 Stvb-i를
가지고 있었으며 선발된 저항성 계통 중 RL1은 Xa3+xa5+Xa21+Stvb-i가 집적되어 있었고, RL2, RL3 및 RL5는 Xa3+
Xa21+Stvb-i 유전자 조합이었으며 RL4은 Xa21 유전자를 보
유하고 있었다(Fig. 2).벼흰잎마름병 및 줄무늬잎마름병 저항성 생물검정
육성계통과 운광, 조평에 대해서 벼흰잎마름병 대표균계 및 줄무늬잎마름병 생물검정을 실시하였다. 운광과 조평은 벼 흰잎마름병 K1, K2, K3균계에 저항성이었으며, 운광은 줄무 늬잎마름병에 이병성이나 조평은 저항성 반응을 나타냈다. 육 성계통 중 벼흰잎마름병 K1균계에 중도저항성과 줄무늬잎마 름병에 이병성을 나타낸 RL4를 제외한 4계통 모두 벼흰잎마
Table 3. Comparison of the control and K3a inoculation about rice yield, ratio of ripened grain, and perfect kernel of brown rice.
Variety(lines)
Control K3a inoculation
Rough rice yield
(g)
Brown rice yield
(g)
Ratio of ripened grain
(%)
Perfect kernel of brown rice
(%)
Rough rice yield
(g)
Brown rice yield
(g)
Ratio of ripened grain
(%)
Perfect kernel of brown rice
(%)
Unkwang 512 412 80.2 80.9 435
**349
**69.8
**71.7
**Jopyeong 481 392 82.4 81.3 411
**335
**73.3
**73.1
**RL1 450 369 80.9 86.5 444
ns363
ns80.3
ns85.2
nsRL2 483 391 82.6 84.5 471
ns379
ns80.5
ns82.7
nsRL3 489 396 79.3 83.4 480
ns388
ns77.6
ns81.8
*RL4 520 418 75.9 78.5 510
ns409
ns74.8
ns77.3
nsRL5 519 417 76.7 83.6 507
ns407
ns75.9
ns82.9
nsz
ns,
*, and
**mean no significant, significant at p < 0.05, and p < 0.01 by t-test, respectively
Table 4. Yield-related traits of rice varieties and breeding lines at yield trial.
Variety (lines)
Heading date (DAS
z)
Culm length (cm)
Panicle length
(cm)
No. of panicles per hill
No. of spikelets
per panicle
Ratio of ripened
grain (%)
1,000 grain weight of brown rice
(g)
Rough rice yield
(kg/10a)
Brown/
rough rice ratio
(%)
Brown rice yield
(kg/10a)
Index of brown rice yield
y(%)
Unkwang 95bcd
x68b 21ab 13b 113a 79.2b 22.9ab 737abc 79.7a 588abc 100
Jopyeong 91e 74a 20b 16a 94c 84.1a 21.1c 694c 79.7a 553c 94
RL1 96ab 72a 22a 14ab 90c 77.6b 20.3d 609d 77.2b 471d 80
RL2 93d 64d 20ab 15ab 89c 84.2a 21.1c 711bc 78.0b 555c 94
RL3 94cd 62d 21ab 14ab 96bc 78.3b 22.5b 720abc 77.8b 560bc 95
RL4 97a 66bc 21ab 13b 102b 74.7c 22.7ab 761ab 79.5a 605a 103
RL5 96ab 66c 22a 15ab 93c 72.8d 23.1a 763a 78.4b 598ab 102
z
Day after seedling
y
Brown rice yield of Unkwang was a standard
x
Means with the same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
름병 K1, K2, K3, K3a균계와 줄무늬잎마름병에 강한 저항성반응을 나타냈다(Table 1). 저항성 계통의 벼흰잎마름병에 대 한 광범위 저항성 반응을 확인하기 위해서 국내 수집 16개 균 주에 대한 저항성 반응을 조사하였다(Table 2). 벼흰잎마름병 저항성 유전자 Xa3를 가지고 있는 운광과 조평은 HB1014, HB1015, HB2010, HB3011, HB4027 균주에 대해서만 저항 성을 나타냈으며, Xa21를 가지고 있는 RL4는 HB2010, HB3011, HB4027에 이병성을 보였고 나머지 균주에 대해서 는 저항성을 나타냈다. Xa3과 Xa21유전자를 함께 가지고 있 는 RL2, RL3, RL5와 Xa3, xa5, Xa21 세 개의 저항성 유전 자가 집적된 RL1의 경우 16개 균주 모두에 대해서 저항성 반 응을 나타냈으며 저항성은 세 개의 유전자가 집적된 RL1이 두 개의 저항성 유전자가 집적된 RL2, RL3, RL5에 비해 강 한 저항성 반응을 보였다.
최근 문제시되고 있는 K3a 균계에 대한 저항성 검정을 한 결과 운광과 조평은 K3a 균계 접종시 수량, 등숙률 및 현미 정상립이 통계적으로 유의하게 감소하였으나 RL1 등 저항성 계통은 대조구와 큰 차이가 없었다(Table 3).
육성계통의 농업형질 특성
선발된 저항성 계통과 반복친인 운광 및 평야지 적응 조생 품종인 조평의 농업형질 특성을 조사하였다(Table 4). 출수기 는 조평이 가장 빨랐으며 RL2, RL3, 운광, RL1, RL5, RL4 순으로 저항성 계통 모두 조생종에 속하였다. 간장은 RL1 (72 cm)이 조평(74)과 비슷한 수준이었으며 나머지 계통들은 반복친인 운광(68)에 비해서도 작은 단간 계통이었고 수장은 RL1과 RL5(22 cm)가 길고 나머지는 비슷한 수준(20, 21)이 었다. 수수는 조평(16개)이 가장 많았고 RL2와 RL5(15),
Tabel 5. Quality-related traits of milled rice of varieties and breeding lines at yield trial.
Variety (lines) Head rice (%)
Opaque rice (%)
Damaged rice (%)
Cracked rice (%)
Colored rice (%)
Protein content (%)
Amylose content (%)
Glossiness (TOYO value)
Unkwang 81.2c
z10.3a 0.4b 2.9c 5.2a 5.2b 19.0b 69.1a
Jopyeong 81.8c 7.5b 0.6b 4.7b 5.4a 6.0a 18.1c 66.4b
RL1 87.5a 7.5b 0.4b 3.1c 1.5b 6.1a 20.4a 64.2c
RL2 85.0b 7.9b 0.1c 5.2b 1.8b 6.1a 20.0a 66.3b
RL3 83.8bc 10.5a 0.2bc 4.3bc 1.2b 5.8ab 19.6ab 65.9b
RL4 79.4d 11.8a 1.3a 6.5a 1.0b 5.5b 18.5c 65.8b
RL5 83.2bc 10.1a 0.3bc 5.0b 1.4b 5.7ab 19.4ab 70.2a
z
Means with the same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
RL1과 RL3 (14), 운광과 RL4(13) 순이었으며 수당립수는반복친인 운광이 113개로 가장 많았고 RL4(102개), RL3 (96), 조평(94), RL5(93), RL1(90), RL2(89) 순이었다. 등숙 률은 RL2(84.2%)와 조평(84.1)이 높았으며 운광(79.2), RL3 (78.3)와 RL1(77.6)이 중간, RL4(74.7), RL5(72.8) 순으로 낮았다. 천립중은 RL5(23.1 g)이 운광(22.9), RL4(22.7)와 RL3(22.5)과 비슷한 수준이었으며 조평(21.1)과 RL2(21.1), RL1(20.3) 순으로 가벼웠다. 정조 수량은 RL5(763 kg/10a) 와 RL4(761)가 반복친인 운광(737)보다 높았고 RL3(720), RL2(711), 조평(694), RL1(609) 순으로 낮았다. 정현비율은 운광(79.7%), 조평(79.7)과 RL4(79.5)가 RL5(78.4), RL2 (78.0), RL3(77.8), RL1(77.2)에 비해 높았다. 현미 수량은 RL4(605 kg/10a)와 RL5(598)가 반복친인 운광의 588 kg/10a에 비해 높았으며 RL3(560)과 RL2(555)가 조평(553) 과 비슷하였고 RL1은 471 kg/10a로 운광 대비 80%의 낮은 수준이었다.
생산력검정시험의 백미를 이용하여 외관품위 및 품질 특성 에 대해 조사하였다(Table 5). 외관품위조사에서 RL4를 제외 한 저항성계통들이 운광과 조평에 비해 정상립이 높고 피해 립 및 착색립이 낮은 등 외관품위가 양호하였으며 그 중 RL1 이 가장 양호하였다. 단백질 함량은 운광(5.2%)이 가장 낮았 고 RL4(5.5), RL5(5.7)와 RL3(5.8), 조평(6.0), RL1(6.1), RL2(6.1) 순으로 높았으며, 아밀로스 함량은 조평(18.1%)이 가장 낮고 RL4(18.5), 운광(19.0), RL5(19.4)와 RL3(19.6), RL2(20.0)와 RL1(20.4) 순으로 높았다. 취반미의 기계적 윤 기치는 RL5(70.2)와 운광(69.1)이 높았고 조평(66.4), RL2 (66.3), RL3(65.9)와 RL4(65.8)이 중간이었으며 RL1(64.2) 이 낮았다.
고 찰
조생종 벼 품종이 평야지에서 추석 전 조기 햅쌀 출하, 이 모작 등 착부체계의 다양화, 수확시기의 다변화를 통한 노동 력 분산을 위해 재배면적이 크게 증가하고 있는 추세이나 조 생종 벼 품종은 다른 생태형에 비해 벼흰잎마름병 및 줄무늬 잎마름병에 대한 저항성이 취약하여 병 발생에 따른 피해가 우려되고 있다.
2004년에 개발된 조생종 최고품질 벼 품종인 운광은 도복 에 안정적이고 다수성이며 밥맛이 좋아서 2008년부터 조생종 벼 품종 중에 가장 많은 재배면적을 차지하고 있다. 또한 우 리나라에서 육성된 조생종 품종 중에서는 처음으로 벼흰잎마 름병 저항성 유전자 Xa3를 도입하였다. 하지만 줄무늬잎마름 병에 대한 저항성이 없고 Xa3유전자가 병원성이 강한 흰잎마 름병 균계에 의해 저항성이 붕괴된 상황에서 평야지에서 재 배될 경우 병 발생에 따른 피해가 발생할 수 있다. 이를 보완 하고자 운광을 모본으로 하고 세 개의 벼흰잎마름병 저항성 유전자 Xa4+xa5+Xa21가 집적되어 있고(Suh et al. 2013) 줄 무늬잎마름병 저항성 유전자
Stvb-i를 가지고 있는 SR30075
조합의 계통을 저항성 유전자 수여친으로 활용하여 저항성 육종사업을 수행하였다. 운광에 SR30075를 교배하고 저항성 유전자에 대한 분자표지를 이용하여 Xa3는 이형접합체이고xa5, Xa21, Stvb-i는 동협접합체인 F
3 계통 SR31206-12를 선발하여 운광에 여교배하였다. Xa3와 Xa4는 11번 염색체에 매우 밀접하게 연관(Yoshimura et al. 1992, Yoshimura et al. 1995)되어 있어 이 중 Xa3를 목표유전자로 하였다. Park et al. (2013, 2014a)은 Xa3는 자포니카 일본 품종인 Wase Aikoku 3로부터 유래하여 1991년에 우리나라 화영 품종에Fig. 3. Radial graph of reaction of resistant lines carrying resistance genes, Xa3, xa5, and Xa21 against each of 16 Korean Xanthomonas oryzae pv. oryzae isolates.
도입된 이후 지금까지 열악형질 수반에 대한 문제 없이 안정 적으로 이용되고 있는 저항성 유전자인 반면 Xa4는 인디카 품종 TKM 6로 부터 유래하여 우리나라 자포니카 품종 육성 에 활용된 적이 없기 때문에 Xa4 도입에 따른 파악되지 않은 문제점이 발생할 수 있으므로 면밀한 검토가 필요하다고 하 였다. 분자표지 선발 후 운광에 1회 여교배한 HR28021과 2 회 여교배한 HR28420 조합에 대해서 Xa3의 저항성을 붕괴 시켜 피해가 확산되고 있는 K3a균계(Kim et al. 2009, Noh et al. 2003)에 대한 저항성 생물검정과 초형과 수량성 등 농 업형질에 대한 조사와 선발을 통해서 5개 저항성 계통을 육성 하였다(Table 1, Fig. 1). 분자표지를 이용하여 저항성 유전자 를 확인한 결과 이들 중 1회 여교배한 계통 RL1만 네 개의 목표 저항성 유전자 Xa3+xa5+Xa21+Stvb-i를 가지고 있었고 2회 여교배한 계통인 RL2, RL3 및 RL5는 Xa3+Xa21+Stvb-i 를, RL4는 Xa21을 단독으로 가지고 있었다. K3a균계에 대해 서 Xa3+xa5+Xa21, Xa3+Xa21 뿐만 아니라 Xa21도 저항성 을 나타내기 때문에 흰잎마름병에 대한 생물검정에서 하나의 균계를 이용할 경우에는 K3a에 대해서 Xa21보다 저항성 수 준이 낮은 xa5나, Xa3와 같이 균계 특이적 이병성을 나타내 는 저항성 유전자의 효과를
Xa21이 질적으로 보완(Park et
al. 2013)하기 때문에 유전자 집적 여부를 구분하기 어렵다.저항성 계통들의 유전자 집적이 다른 것은 분자표지를 이용 한 선발 없이 K3a균계에 대한 생물검정과 초형, 수량성 등 농 업형질에 대한 선발강도를 높여서 선발을 해나가는 과정에서 목표 유전자가 유실되었기 때문으로 생각한다. Park et al.
(2012)은 Xa21가 새로운 변이균인 K3a균계에는 강한 저항성 을 발휘해서 저항성원으로써의 가치가 인정되나 K1균계에는 이병성이므로 이를 보완해야 한다고 하였다. 이번 실험에서
Xa21을 단독으로 가지고 있는 저항성 계통 RL4의 경우에는
K1 균계로 쓰인 HB1013 균주에 대해서 중도저항성을 나타 냈는데 이는 검정 당시 균주의 활력이 떨어졌거나 RL4의 다 른 유전적 배경에 의해서 저항성 반응이 다소 다르게 나온 것 으로 생각되었고 다른 K1 균계인 HB2010, HB3011, HB4027 균주의 경우 2013년 Park et al.의 보고와 같이 이병성 반응 (Fig. 3)을 나타내 이에 대한 보완이 필요하였다. Xa21에 Xa3 이 결합된 Xa3+Xa21 조합의 계통인 RL2, RL3, RL5의 경우Xa3+xa5+Xa21 세 개의 저항성 유전자 집적 계통 RL1에 비
해 저항성 반응이 다소 약하였지만 16개 균주 모두에 대해서 저항성 반응(Table 2)을 나타냈으며 K3a 균계 접종시 현미 수량, 등숙률 및 현미 완전미율이 대조구와 차이가 없어(Table3) 벼흰잎마름병 저항성 증진을 위한 유망한 조합으로 생각되 었다. 줄무늬잎마름병의 경우 Stvb-i 저항성 유전자가 없는 운 광과 RL4만 생물검정에서 이병성을 나타냈다. Stvb-i는 1975 년에 최초의 저항성 품종인 낙동벼가 개발된 이후로 저항성 품종개발에 지속적으로 이용되고 있으나 Stvb-i에 따른 열악 형질 수반이나 저항성 붕괴에 따른 문제는 보고되지 않아 (Park et al. 2014b) 저항성 유전자가 도입된 RL1, RL2, RL3, RL5의 경우 줄무늬잎마름병 저항성 계통으로 활용가치 가 있다고 판단되었다.
평야지에 적응하는 조생종 품종으로써 농민의 선택을 받기 위해서는 벼흰잎마름병과 줄무늬잎마름병에 대한 저항성 증 진과 함께 초형, 수량성, 미질 등 다른 농업형질 특성도 우수 하여야 한다. 운광에 여교배 1회 된 RL1의 경우 목표 저항성 유전자를 모두 갖추고 있고 저항성도 우수하였지만 간장이 크고 도복에 대한 안정성이 운광에 비해 떨어지며 수량성이 운광의 80% 수준으로 공시 재료 중 가장 낮았고, 단백질과 아밀로스 함량은 높고 취반미의 윤기치가 낮아 식미 측면에 서 좋지 않은 특성을 나타냈다(Table 4, 5). 여교배가 2회 되 어 선발된 계통들은 모두 간장이 운광보다 작은 단간으로 도 복에 안정적인 특성을 나타냈으며, RL4와 RL5의 경우 운광 보다 높은 수량성을 나타내 다수성 계통으로 유망시 되었다.
RL5는 백미 완전미율이 운광에 비해 높고 취반미의 식미치 도 운광과 비슷하게 높아 미질 및 식미 측면에서 좋은 특성을
나타냈다. 따라서 저항성 계통 RL5는 벼흰잎마름병 및 줄무 늬잎마름병에 효과적인 Xa3+Xa21+Stvb-i 저항성 유전자 조 합으로 도복에 안정적이고 수량이 높으며 미질이 양호한 우 량계통으로 이를 조생종 벼 품종의 평야지 적응성 향상을 위 한 자원으로 적극 활용할 계획이다.
적 요
본 연구는 조생종 벼의 평야지 적응성을 향상시키고자 벼 흰잎마름병 및 줄무늬잎마름병에 대한 저항성 유전자가 집적 된 저항성 계통을 개발하고 이들의 저항성 성능검정과 수량 성 등 농업형질을 분석하여 육종사업에 반영하고자 수행하였 다. Xa3를 가지고 있는 운광과 Xa4+xa5+Xa21+Stvb-i를 가 지고 있는 SR30075 조합 계통을 모본으로 하여 분자표지를 이용하여 저항성 유전자가 집적된 F3 계통을 선발하였고 운 광에 여교배하였다. 여교배 이후에 K3a균계에 대한 생물검정 및 수량성 등 농업형질에 대한 조사를 통해 RL1 (BC1F7), RL2-5 (BC2F6) 등 5개 조생종 저항성 계통을 육성하였다. 분 자표지로 저항성 유전자를 확인한 결과 RL1은 Xa3+xa5+Xa21+
Stvb-i를 가지고 있었고 RL2, RL3와 RL5 는 Xa3+Xa21+
Stvb-i, RL4는 Xa21를 보유하고 있었다. 줄무늬잎마름병은
저항성 유전자 Stvb-i의 유무에 의해서 저항성이 결정되었다.벼흰잎마름병 유전자 조합
Xa3+xa5+Xa21과 Xa3+Xa21의
계통의 경우 K1, K2, K3, K3a 균계 및 16개 수집 균주에 대 해서 저항성 반응을 나타냈으며, K3a균계 접종시에 현미수 량, 등숙률 및 현미 완전미율이 대조구와 차이가 없어 저항성 증진을 위한 유망조합으로 판단되었다. RL1은 목표 저항성 유전자를 가지고 있고 저항성 성능도 우수하였으나 간장이 크고 도복에 불안정하였으며 수량이 운광의 80%로 낮고 미 질이 좋지 않았다. RL5는 운광에 여교배가 2회 되어 선발된 계통으로 저항성 유전자 Xa3+Xa21+Stvb-i를 가지고 있어 벼 흰잎마름병 및 줄무늬잎마름병에 대한 효과적인 저항성원으 로 판단되었고, 운광보다 단간으로 도복에 안정적인 특성을 나타냈으며 다수성이고 미질이 양호하였다.사 사
본 논문은 농촌진흥청 연구사업(과제번호: PJ00871006)의 지원에 의해 이루어진 것임.
REFERENCES