S. Y. Lee ( ) ・ J. L. Lee
National Institute of Horticultural & Herbal Science, Rural Development Administration, Suwon 440-441, Korea e-mail: [email protected]
D. Y. Kim
Bio-Crop Development Division, National Academy Agricultural Science, Rural Development Administration, Suwon 441-707, Korea
Increase of resistance to oxidative stress induced by methyl viologen in progeny from a cross between two transgenic Petunia lines with NDPK and SOD genes
Su Young Lee ・ Jung Lim Lee ・ Dool Yi Kim
Received: 2 August 2011 / Accepted: 26 August 2011
ⓒKorean Society for Plant Biotechnology
Abstract This study was conducted to investigate how to enhance resistance to oxidative stress in petunia progeny ob- tained by a crossing between transgenic plants, MnSOD (SOD2) (T4) and NDPK2 (T2), to develop transgenic petunia much more resistant to environmental stress. At the treatment of MV 200 μM, the progeny was significantly less damaged than its parental plants (SOD2- or NDPK2-transgenic lines) as well as wild type plants, implying its resistance to oxidative stress was enhanced compare to that of SOD2- or NDPK2- transgenic plants. In an expression of 11 quantitative traits, the progeny remained similar to control plants, although it infrequently displayed slightly longer or wider than either parental or wild type plants. In the expression of 6 qualitative traits, there was no significant difference between parental or non-transgenic control plants.
Keywords Environmental stress, Morphological traits, PCR, Petunia hybrida
Introduction
Many researchers have studied to develop plants resistant to the abiotic stresses through the transfer of superoxide dismutase (SOD) or ascorbate peroxidase (APX) or nucleo- tide diphosphate kinase (NDPK) genes in many crops (Fang
et al. 2002; Kim et al. 2005; Lee et al. 2009a, 2009b, 2010; Moon et al. 2003; Tang et al. 2004a, 2004b, 2007).
These crop cultivars that are more resistant to environmen- tal stress (or abiotic stress) are needed in times of unpre- dictable weather condition. Recent developments in plant genetic engineering are aimed at increasing resistance by integration of multiple transgenes into plants genome (multiple-transgene-stacking) and coordinated expression of these transgenes in transgenic plants (François et al.
2002). Also, there were recent reports on transgenic plants that are more resistant to abiotic stess through the intro- duction of two genes (SOD and APX genes) (Kwon et al.
2002; Lim et al. 2007).
Since petunia is mainly used to be planted on the roadside in company with pansy in Korea, there is a need to develop cultivar resistant to abiotic stresses such as rain- fall, humidity, and air pollution. The Floricultural Research Division at National Institute of Horticultural & Herbal Science has previously developed the MnSOD (SOD2) transgenic petunia (T4) as well as NDPK2 (T2) transgenic petunia pure lines (Lee et al. 2009a). This study used conventional breeding technique (crossing) as means for multi-transgene-stacking methods to get transgenic petunia line more resistant to oxidative stress than the previously developed transgenic lines with SOD2 and NDPK2 genes, respectively. After that, this study identified how to enhance the resistance of its progeny to oxidative stresses induced by methyl viologen (MV). Furthermore, this study investi- gated 17 morphological traits in progeny to identify that there were no significant differences between the progeny and its wild type or parental plants in other morphological characters besides the resistance to oxidative stress contro- lled by the transfer of the genes.
DOI:http://dx.doi.org/10.5010/JPB.2011.38.3.215 Research Article
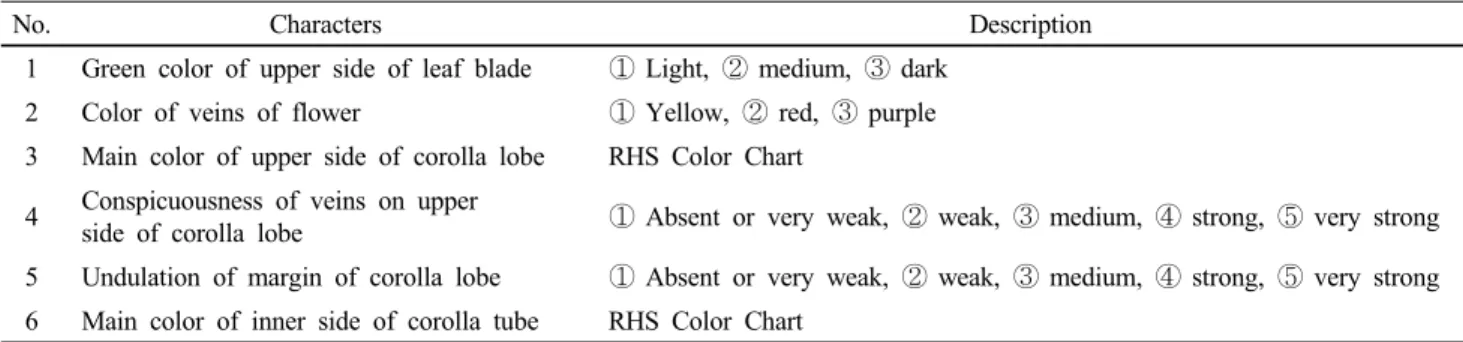
Table 1 List of six qualitative characters examined in progeny from a cross of transgenic petunia lines, NDPK2-5-3 (T2) and SOD2-2-1-1-35 (T4)
No. Characters Description
1 Green color of upper side of leaf blade ① Light, ② medium, ③ dark 2 Color of veins of flower ① Yellow, ② red, ③ purple 3 Main color of upper side of corolla lobe RHS Color Chart
4 Conspicuousness of veins on upper
side of corolla lobe ① Absent or very weak, ② weak, ③ medium, ④ strong, ⑤ very strong 5 Undulation of margin of corolla lobe ① Absent or very weak, ② weak, ③ medium, ④ strong, ⑤ very strong 6 Main color of inner side of corolla tube RHS Color Chart
PCR analysis
Integration of transgene in the progeny was identified thr- ough PCR analysis. DNA was isolated from leaves of the progeny grown for 45 days after sowing in the greenhouse by using DNeasy plant mini kit (QIAGEN Co.) and quanti- fied by using NanoDrop (Nano Co.). PCR reaction was con- ducted as described by Lee et al. (2009a) using NDPK2 specific primers (forward; 5’-GAG GCT ATT CGG CTA TGA CTG-3’, reverse; 5’-ATC GGG AGC GGC GAT ACC GTA-3’) and SOD2 gene specific primers(forward;
5'-TAC TGG AGA TGA ATA TGA GC-3', reverse; 5'-CA G CAG GCG GCA AAT GAT TA-3'), respectively.
Treatment of MV on whole plant
Seedlings of progeny grown for 20 days after sowing on a plastic box containing 400g of culture medium mixed wi- th perlite (SungHyun Perlite Co., South Korea) and com- mercial compost were transplanted to plastic pots (15cm) with just above culture medium. After 15 days in the gr- eenhouse, the progeny was sprayed with about 3.75 mL of MV solutions in 0, 100, and 200 μM dissolved in 20%
acetone solvent supplemented with 0.1% (w/v) Tween 20 at 3 plants per treatment. Each treatment was repeated three times. The percentage of leaf damage that appeared on the leaves in the transgenic plants after MV spraying was eval- uated at 5 days after treatment [0 (no damage) to 100 (com- pletely killed).
while its flowers were blooming.
Results and Discussion
Identification of the transfer of SOD2 and NDPK2 genes in progeny
In common, after proceeding generations of transgenic plants, stability or inheritance of transgene in progeny plants has been identified according to PCR analysis (Chareonpor- nwattana et al. 1999; Sriskandarajah et al. 2007; Zhang et al. 2005). However, there was a recent report on presence of inserted genes in transgenic plant by fluorescence in situ hybridization (FISH) (Choi et al. 2009). This study, also, the integration of two transgenes in progeny obtained from the crossing between the independently transformed petunia lines, SOD2-2-1-1-35 and NDPK2-5-3 obtained from petunia pure line Wongyo A2-36, was analyzed using PCR.
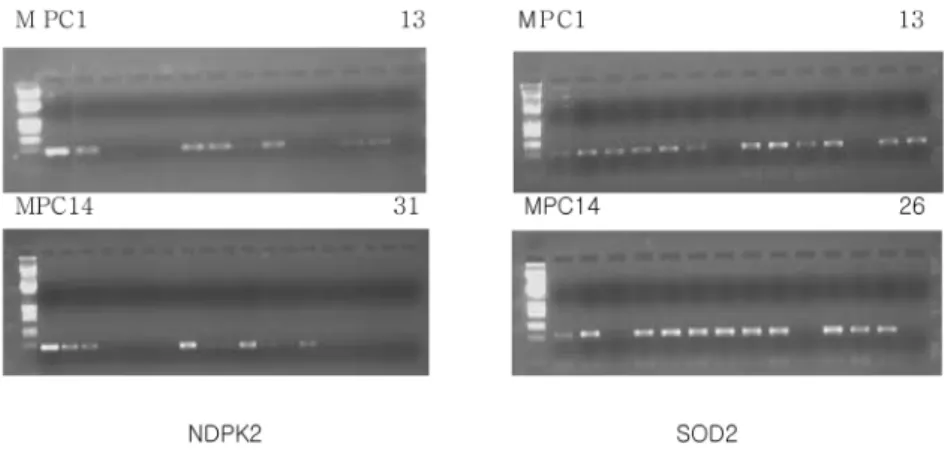
The number of plants with two genes (SOD2 and NDPK2) was 23 out of 56 seedlings of the progeny (Table 2 and Fig. 1).
Enhancement of resistance to oxidative stress induced by MV in progeny
Methyl viologen (MV, paraquat) has been used to identify the resistance of transgenic plants to oxidative stress because it induces treatment of superoxide anion (O2-) (singlet oxy- gen) as well as hydroxyl and peroxy radicals. (Kim et al.
Fig. 1 PCR analysis of progeny (31 plants) from a crossing of transgenic petunia lines, NDPK2-5-3 (T2) and SOD2-2-1-1-35 (T4), using NDPK2 or SOD2 specific primers. M, λEcoRⅠ/HindⅢ; PC, positive control
Table 2 The number of transgenic plants with SOD2 and/or NDPK2 gene based on PCR analysis in progeny from a cross of transgenic petunia lines, NDPK2-5-3 (T2) and SOD2-2-1-1-35 (T4)
No. of analyzed plants
No. of plants with both SOD2 and NDPK2 genes
No. of plants with NDPK2 gene
No. of plants with SOD2 gene
No. of plants without both SOD2 and NDPK2 genes
56 23 16 6 11
Table 3 Visible injury (%) of progeny from a cross of transgenic petunia lines, NDPK2-5-3 (T2)and SOD2-2-1-1-35 (T4), 5 days after spraying different methyl viologen (MV) (paraquat) concentrations in the greenhouse
Genotypes Visual injury (%)
MV 0 µM MV 100 µM MV 200 µM
NDPK2-5-3 (T2) 0.0±0.0* 3.0±1.7 23.3±15.3
SOD2-2-1-1-35 (T4) 0.0±0.0 10.0±0.0 38.3±2.9
NDPK2-5-3ⅹSOD2-2-1-1-35 0.0±0.0 10.0±5.0 16.7±11.5
A2-36 (Non-transgenic control) 0.0±0.0 11.7±2.9 65.0±21.8
* Mean±standard deviation
Fig. 2 Responses of progeny from a cross of transgenic petunia lines, NDPK2-5-3 (T2) and SOD2-2-1-1-35 (T4), 5 days after spraying at different methyl viologen (MV) (paraquat) concentrations (0, 100, and 200 µM) in the greenhouse. MV solutions were sprayed star- ting from the bottom (0 µM) to the third row (200 µM). A NDP K2-5-3, B SOD2-2-1-1-35, C Progeny from a cross of NDPK2- 5-3 and SOD2-2-1-1-35, D Non-tansgenic control plants
2005; Kim et al. 1986; Kwon et al. 2002; Lim et al. 2007;
Moon et al. 2003; Suntres 2002; Tang et al. 2008). In case the transgenic plant is less damaged after treatment with MV than wild type, it will be concluded that the resistance of transgenic plant to oxidative stress is more increased than that of wild type plant. After the progeny from the cross of transgenic Petunia pure lines, NDPK2-5-3 and
SOD2-2-1-1-35, was treated with MV of three solutions (0, 100 and 200 μM), leaf injury caused by MV was obser- ved next day after the treatment and became severe therea- fter like in previous study (Lee et al. 2010). Visible injury of the progeny on the fifth day after treatment was shown in Table 3 and Figure 2. The progeny with both SOD2 and NDPK2 genes was less damaged than the non-transgenic
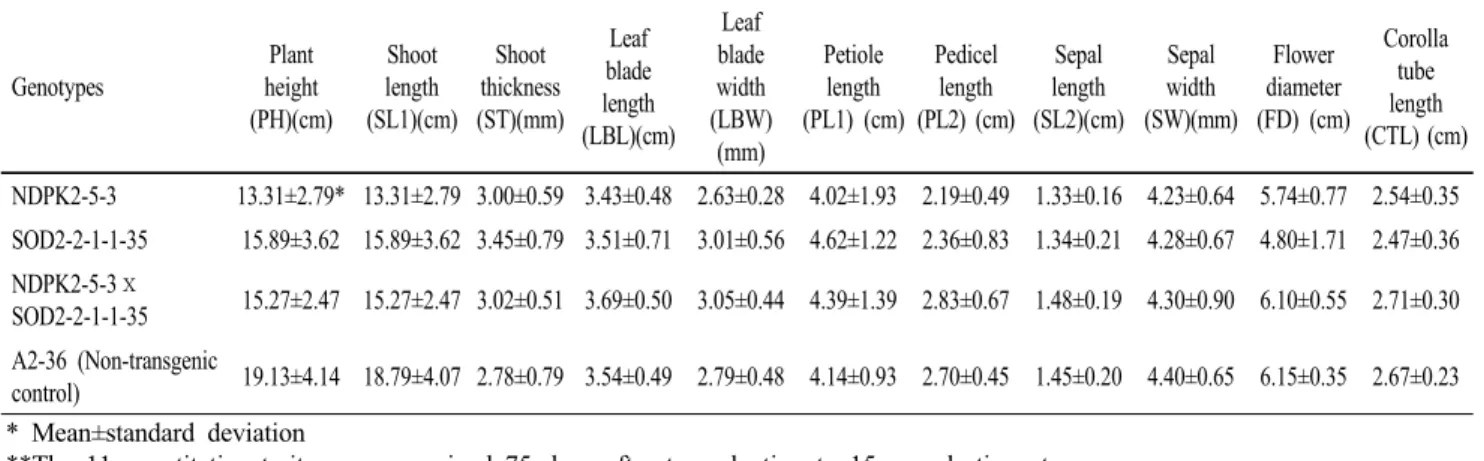
Table 4 Mean for 11 quantitative traits expressed in progeny from a cross of transgenic petunia lines, NDPK2-5-3 (T2) and SOD2-2-1-1-35 (T4)
Genotypes
Plant height (PH)(cm)
Shoot length (SL1)(cm)
Shoot thickness (ST)(mm)
Leaf blade length (LBL)(cm)
Leaf blade width (LBW)
(mm)
Petiole length (PL1) (cm)
Pedicel length (PL2) (cm)
Sepal length (SL2)(cm)
Sepal width (SW)(mm)
Flower diameter (FD) (cm)
Corolla tube length (CTL) (cm) NDPK2-5-3 13.31±2.79* 13.31±2.79 3.00±0.59 3.43±0.48 2.63±0.28 4.02±1.93 2.19±0.49 1.33±0.16 4.23±0.64 5.74±0.77 2.54±0.35 SOD2-2-1-1-35 15.89±3.62 15.89±3.62 3.45±0.79 3.51±0.71 3.01±0.56 4.62±1.22 2.36±0.83 1.34±0.21 4.28±0.67 4.80±1.71 2.47±0.36 NDPK2-5-3ⅹ
SOD2-2-1-1-35 15.27±2.47 15.27±2.47 3.02±0.51 3.69±0.50 3.05±0.44 4.39±1.39 2.83±0.67 1.48±0.19 4.30±0.90 6.10±0.55 2.71±0.30 A2-36 (Non-transgenic
control) 19.13±4.14 18.79±4.07 2.78±0.79 3.54±0.49 2.79±0.48 4.14±0.93 2.70±0.45 1.45±0.20 4.40±0.65 6.15±0.35 2.67±0.23
* Mean±standard deviation
**The 11 quantitative traits were examined 75 days after transplanting to 15 cm plastic pot
Table 5 MAnalysis of 11 quantitative traits expressed in progeny from a cross of transgenic petunia lines, NDPK2-5-3 (T2) and SOD2-2-1-1-35 (T4)
Genotypes
>only non-transgenic control but<
their parents
>both non-transgenic control and their maternal parent
>both non-transgenic control and their paternal parent
>both non-transgenic control and their parents
<only non-transgenic control but >
their parents
<both non-transgenic control and their maternal parent
<both non-transgenic control and their paternal parent
<both non-transgenic control and their parents NDPK2-5-3ⅹ
SOD2-2-1-1-35 - ST*, PL1 - LBL, LBW,
PL2, SL2, CTL SW, FD - PH, SL1 -
*Abbreviated word is described at Table 4
al. (2008) reported that a transgenic chinese cabbage with both MnSOD and catalase genes of Escherichia coli by co-transformation exhibited tolerance to sulfur dioxide com- pare to the transgenic cabbage with the MnSOD gene and the catalase one. Chan et al. (2005) reported that a trans- genic Phalenopsis with both CymMV coat protein cDNA (CP) gene and sweet pepper ferredoxin-like protein cDNA (Pflp) gene by double transformation enhanced resistance to viral and bacterial phytopathogens compare to the trans- genic Phalenopsis with the CymMV coat protein cDNA (CP) gene and the sweet pepper ferredoxin-like protein cDNA (Pflp) gene.
Morphological traits expressed in progeny
Theoretically, only a specific characteristics controlled by the transfer of a gene would be changed in a transgenic
transferase (IPT) gene-transgenic petunia lines exhibited horticultural characteristics different from its wild type plant, whereas the other line were not greatly altered in any other horticultural traits in comparison to wild type plant.
This study investigated 17 morphological traits expressed in the progeny between two transgenic petunia lines with SOD2 or NDPK2 gene compared to the traits of wild type as well as its parental (SOD2- or NDPK2-transgenic) plants.
The examined data is shown in Table 4 and 6. In this study, also, the progeny remained similar to wild type plant in the expression of 11 quantitative traits, although it showed slightly longer or wider characteristics than either its paren- tal and/or wild type plants. The leaf blade, corolla tube, pedicel and sepal of the progeny were slightly longer and wider than that of both its parental and wild type plant, whereas the sepal and flower diameter of progeny was a bit wider and longer, respectively, than those of its parental
Table 6 Six qualitative traits expressed in progeny from a cross of transgenic petunia lines, NDPK2-5-3 (T2) and SOD2-2-1-1-35 (T4)
Genotypes
Green color of upper side
of leaf blade
Color of veins of flower
Main color of upper side of corolla lobe
Conspicuousness of veins on upper side of
corolla lobe
Undulation of margin of corolla lobe
Main color of inner side of corolla tube
NDPK2-5-3 medium red 45B weak weak 4B
SOD2-2-1-1-35 medium red 40A medium medium 6B
NDPK2-5-3ⅹ
SOD2-2-1-1-35 medium red 46B or 48C medium middle weak 157A
A2-36 (Non-transgenic
control) light red 46B or 52C medium weak 157A
*The above 6 qualitative traits were examined 75 days after transplanting to 15cm plastic pot
Fig. 3 Comparison of the color and shape of flowers between progeny plants from a cross of transgenic petunia lines, NDPK2-5-3 and SOD2-2-1-1-35, and its parents or non-transgenic control plants. A NDPK2-5-3, B SOD2-2-1-1-35, C Progeny from a cross of NDPK2-5-3 and SOD2-2-1-1-35, D Non-tansgenic control plants
but those of non-transgenic control plants (Table 5). In the expression of 6 qualitative traits, there is no significant dif- ference between the progeny and its parental or wild type plants (Table 6 and Fig. 3).
In conclusion, this study confirmed the resistance to oxida- tive stress was enhanced by stacking of SOD2 and NDPK2 genes in petunia. The results suggest that transgenic plants expressing two transgenes can be generated by a cross between independently transformed plants. These results also support François et al. (2002)’argument that introduc- tion of multiple transgenes or multiple-transgne-stacking is highly desired. Also, this study suggests that the transgenic progeny seeds with both SOD2 and NDPK2 genes would be valuable data and genetic materials for breeding new petunia cultivar resistant to environmental stress.
Literature Cited
Chan YL, Lin KH, Sanjaya, Liao LJ, Chen WH, Chan MT (2005) Gene stacking in Phalenopsis orchid enhances dual tolerance to pathogen attack. Trans Res 14:279-288
Chareonpornwattana S, Thara KV, Wang L, Data SK, Panbangred W, Muthukrishnan S (1999) Inheritance, expression, and silencing of a chitinase transgene in rice. Theor Appl Genet 98:371-378
Choi HW, Yu XH, Lemaux PG, Cho MJ (2009) Stability and inheri- tance of endosperm-specific expression of two transgenes in progeny from crossing independently transformed barley pl- ants. Plant Cell Rep 28:1265-1272
Clark DG, Dervinis C, Barrett JE, Klee H, Jones M (2004) Drought- induced leaf senescence and horticultural performance of trans- genic PSAG12-ipt petunias. J Amer Soc Hort SCI 129:93-99 Fang GC, Hanau RM, Vaillancourt LJ (2002) The SOD2 gene,
encoding a manganese-type superoxide dismutase, is up- regulated during conidiogenesis in the plant pathogenic fungus Colletotrichum graminicola. Fungal Genetics and Biology 36:155-165
François IEJA, Broekaert WF, Cammue BPA (2002) Different approaches for multi-transgene-stacking in plants. Plant Sci 163:281-295
Kim JS, Lee BH, Kwon SY, Kim YH, Kim SH, Cho KY (2005) Antioxidative responses of transgenic tobacco plants express- ing both superoxide dismutase and ascorbate peroxidase in chloroplasts to several herbicides. Kor J Plant Biotech 32:
97-103
Kim KU, Kim DU, Kwon ST (1986) Development of herbicide (paraquat) tolerant plant through tissue culture. Korea J Weed Sci 61:191-200
Kwon DY, Jeong YI, Lee HS, Kim JS, Cho KY, Allen RD, Kwak SS (2002) Enhanced tolerance of transgenic tobacco plants expressing both superoxide dismutase and ascorbate peroxi- dase in chloroplasts against methyl viologen-mediated oxida- tive stress. Plant, Cell and Environment 25:873-882
Lee SY, Han BH, Cho AY (2009a) Inheritance and expression of transgene in SOD2-transgenic petunia descendants and their
sweetpotato plants that express both CuZnSOD and APX in chloroplasts to methyl viologen-mediated oxidative stress and chilling. Mol Breeeding 19:227-239
Moon H, Lee B, Choi G, Shin D, Prasad DT, Lee O, Kwak SS, Kim DH, Nam J, Bahk J, Hong JC, Lee SY, Cho MJ, Lim CO, Yun DJ (2003) NDP kinase 2 interacts with two oxidative stress- activated MARKs to regulate cellular redox state and enhances multiple stress tolerance in transgenic plants. PNAS 100:358- 363
National Seed Management Office (1997) Standard for survey in application and evaluation of carnation new varieties. pp 32-40 (In Korean)
Rural Development Administration (1995) Standard for survey in agricultural experiment & research. pp 388-390 (In Korean) Sriskandarajah S, Mibus H, Serek M (2007) Transgenic Campanula
carpatica plants with reduced ethylene sensitivity. Plant Cell Rep 26:805-813
31:19-195
Tang L, Kwon SY, Kwak SS, Sung CK, Lee HS (2004b) Selection of transgenic potato plants expressing both CUZNSOD and APX in chloroplasts with enhanced tolerance to oxidative stress. Kor J Plant Biotech 31:109-113
Tang, L, Kim MD, Yang KS, Kwon SY, Kim SH, Kim JS, Yun DJ, Kwak SS, Lee HS (2008) Enhanced tolerance of transgenic potato plants overexpressing nucleoside diphosphate kinase 2 against multiple environmental stresses. Transgenic Res 17:
705-715
Tseng MJ, Liu CW, Yiu JC (2008) Tolerance to sulfur dioxide in Chinese cabbage transformed with both the superoxide dismu- tase containing manganese and catalase genes of Escherichia coli. Sci Hort 115:101-110
Zhang Y,Yin X, Yang A, Li G, Zhang J (2005) stability of inheri- tance of transgenes in maize (Zea mays L.) lines produced using different transformation methods. Euphytica 144:11-22