IEG 환경지질연구정보센터
19
0
0
전체 글
(2) 82. Gerd Geyer. seemed to appear even later because the lowermost Middle Cambrian there appeared to be unfossiliferous. Paradoxides was recognized in Siberia in the 1960s, and the strata in which the earliest occur were not readily correlatable into Scandinavia. However, the earliest Paradoxides in Spain are distinctly older than the oldest species of Scandinavia (Sdzuy, 1967, 1971, 1972, 1995), and the earliest Paradoxides in Morocco occur in even older strata (Geyer, 1983, 1990a, 1990b, 1993, 1998, Geyer and Palmer 1995). “Olenellus” as used by Walcott and his early epigones is more-or-less a synonym of the Olenellina in its traditional concept. It thus included the genera and species of the present-day Family Olenellidae characteristic of Laurentia as well as the Holmiidae and the obviously distantly related Fallotaspidoidea. Ironically, holmiids are the traditional index fossils for the Lower Cambrian in Scandinavia and do not overlap there with Paradoxides so that the traditional concept did not fail a priori. It was even assumed that Olenellus (s.s.) had a longer range and thus occurred in Laurentia at a time when the first Paradoxides individuals turned up in other regions. However, it was difficult, if not impossible to prove this assumption. Direct evidence finally came from the holmiids. Fragments of holmiids occurred in Middle Cambrian (=Paradoxides-bearing) strata of Morocco, and the excavation of Acadoparadoxides specimens in the eastern Anti-Atlas for the international fossil market finally delivered perfect specimens of Cambropallas telesto Geyer 1993 as the youngest known holmiid together with Acadoparadoxides briareus Geyer 1993 well within the range of Paradoxides (s.l.). This permitted a re-evaluation of the stratigraphic significance of the Oleneloidea in general (Geyer and Palmer, 1995). Nevertheless, it also showed that a combination of both traditional concepts of Brögger and Walcott is unsuitable for future subdivision. Several authors have dealt with the problem of a correlation of the Lower-Middle Cambrian boundary interval (e.g., Öpik, 1968; Sdzuy, 1972; Repina and Suvorova, 1983; Rozanov and Sokolov, 1984; Spizharskiy et al., 1983, 1984; Geyer, 1990b, 1998, 1999; Pillola, 1991; Astashkin et al., 1991; Liñán et al., 1992, 1993; Álvaro et al., 1993; Geyer and Palmer, 1995; Zhuravlev, 1995; Gozalo, 1995, Blaker and Peel, 1997; Sundberg and McCollum, 1997; Geyer and Shergold, 2000; Fletcher, 2001, 2003; Korovnikov, 2001; Gozalo et al., 2003; Zhu et al., 2004). In general, all of them showed that an easy solution for a globally recognizable Lower-Middle Cambrian boundary is difficult or impossible to reach when either of the traditional concepts is used as a guideline. But what are the alternatives? 1.1. Material Repositories of the figured specimens are: EM, collection Université Claude Bernard, Lyon; GGU, Grønlands Geologiska Undersøgelse, Copenhagen; IGR, Institut de Géol-. ogie, Université de Rennes; MGUH, Geological Museum, University of Copenhagen; MNHN, Muséum Nationale de l’Histoire Naturelle, Paris; PIW, Institut für Paläontologie, Bayerische Julius-Maximilians-Universität Würzburg; SMF, Naturmuseum Senckenberg, Frankfurt am Main; SUS, collection of Uwe Scheibe, Cottbus. 2. CORRELATION TOOLS The Lower-Middle Cambrian boundary is a long pending problem and has been the subject of discussion within the International Subcommission on Cambrian Stratigraphy (ISCS) at least since 1974. However, despite the recognition of the difficulties, the problem was not dealt with in much detail before the subdivision initiative of the ISCS started in 1999 (Geyer and Shergold, 2000). Discussions in the ISCS indicated that there is a general agreement in that historical perspectives should not necessarily be taken into account. A fresh approach considered Oryctocephalus indicus, which never before played a major role in the Lower-Middle Cambrian boundary discussions, as a suitable marker for the base of a Middle Cambrian series (Yuan et al., 1997; Sundberg et al., 1999; Peng et al., 2000; McCollum and Sundberg, 2002; Sundberg and McCollum, 2003). To date there are several different possibilities for a globally usable Lower-Middle Cambrian boundary. Because “non-traditional methods” (such as carbon isotopes or magnetostratigraphy) can be shown to have no key potential for this stratigraphic interval, a GSSP will need to be based on paleontological data. Carbon isotopes may be useful for the Ediacaran-Cambrian boundary interval and the earliest Cambrian period for which several positive carbon isotope excursions can be detected in various regions (Banerjee et al., 1994; Brasier, 1985, 1991, 1992, 1994; Brasier et al., 1990, 1992, 1993, 1994a, 1994b; Brasier and Sukhov, 1998; Derry et al., 1994). Nevertheless, such distinct excursions are largely absent (or not yet recovered) for the second half of the Early Cambrian and the early Middle Cambrian although regional studies seem to promise useful results (e.g., Montañez et al., 2002). Moreover, during the process of introducing carbon isotopes as a distinctive stratigraphic tool, it became clear that only a calibration with other time-calibrated scales permitted a reliable intercontinental correlation because “early” applications of these techniques were subsequently proved by geochronological and/or biostratigraphical data to be misleading or even erroneous (e.g., Kirschvink et al., 1991; Magaritz et al., 1991). Studies on the “microplankton” since ca. 1990 have shown that distinctive changes in assemblages of acritarchs allow a zonation which is recognizable on various Cambrian continents (e.g., Moczydlowska, 1991, 1998a, 1998b, 2001; Palacios and Moczydlowska, 1998; Yang and Yin, 2001). However, a biased fossil record (Vidal, 1994) as well as taxonomic inconsistencies a priori diminish the possibility of.
(3) The base of a revised Middle Cambrian. recognizing discrete biochrons, which reflect major biotic events. Even during earliest Phanerozoic such events were not only radiations but also declines in diversity and even extinctions (Moczydlowska, 1998a, fig. 11). Secular biodiversity changes in the microplankton (Knoll, 1994; Vidal and Moczydlowska, 1995, 1997; Vidal, 1998; Moczydlowska, 2001) were carefully analyzed for Upper Silesia (Moczydllowska, 1998a). These results indicate that the recorded biotic changes have been influenced by environmental perturbations. After a rapid radiation at the Ediacaran-Cambrian boundary, the acritarch diversity gradually incrased to reach a maximum diversity and abundance in the Baltic Holmia kjerulfi Biozone (Moczydlowska, 1998a), coinciding with the supposed maximum transgression not only on Baltica but on a global scale. The acritarch diversity declined markedly afterwards, reflecting an extinction event and the turnover from the Heliosphaeridium dissimilareSkiagia ciliosa Assemblage Zone (Holmia kjerulfi Biozone) to the Volkovia dentifera-Liepaina plana Assemblage Zone (“Protolenus interval” of the latest Lower Cambrian in traditional concept). A notable turnover in the microplankton at the base of the traditional Middle Cambrian of Baltica reflects a considerable radiation and led to the Eliasum-Cristallinium Assemblage Zone (Moczydlowska, 1998a, 1999) which could have served as a marker for the base of a Middle Cambrian series. However, the base of this assemblage zone is not sharp enough and unknown from some critical regions so that it is not suitable to define a GSSP. Most acritarch species have a very long range and only a regional occurrence. By contrast, some acritarch species of the boundary interval are short-ranging and semi-cosmopolitan but they are very rare. Others may have a distinctive acme in a certain stratigraphic interval but being rare below and above. One example is Heliosphaeridium notatum, which is rare in the “Protolenus zone/interval” but abundant in the Acadoparadoxides oelandicus Zone of Baltica and equivalents so that it indicates the traditional Lower-Middle Cambrian boundary (Moczydlowska, 1998a, 1998b, 1999). “Brachiopods” were long neglected as biostratigraphic tools in the Cambrian. Their stratigraphic value at least for intracontinental correlation has been demonstrated, and they are particularly helpful in rocks where trilobites are absent or rare because of environmental constraints (e.g., Geyer and Mergl, 1995; Mergl et al., 1998). Nevertheless, brachiopods tend to show a strongly regional distribution. Apart from trilobites, frequent skeletal fossils (such as mollusks, chancelloriids, paleoscolecids, or early echinoderms) are either too sparsely occurring, and/or too poorly known in terms of their stratigraphic ranges to be helpful for a precise stratigraphic assignment. Trilobites appear to be the only reliable index fossils capable of defining a GSSP although particularly in the Lower-Middle Cambrian interval trilobites are also affected by major problems that lessen the criteria for a selection of. 83. a GSSP: 1. From the beginning of the fossil record, trilobite faunas of the Cambrian continents show a fair degree of endemicity; 2. intercontinental correlations suffer by a very different state of knowledge; and 3. often different lithofacies create different biofacies which are not yet sufficiently understood according to the stratigraphic ranges of its fossil taxa and lateral extent. Particularly diachronous facies distribution may create problems for exact biostratigraphic transregional schemes. The problem of the Lower-Middle Cambrian boundary has been addressed by Robison et al. (1977), by Geyer (1988, 1990b), Geyer and Palmer (1995), Fletcher (2001, 2003), Sundberg and McCollum (2003), and others. The ISCS developed a Lower-Middle Cambrian Working Group which is progressively initiative (see <http://www2.ewu.edu/WORKINGGROUP/index.htm>). There are five possibilities for global correlation at the Lower-Middle Cambrian boundary interval, which merit scrutiny: 1. FAD of Oryctocephalus indicus; 2. FAD of Ovatoryctocara granulata and/or Kiskinella cristata; 3. FAD of Arthricocephalus chauveaui; 4. Base of the Acidiscus-Cephalopyge assemblage zone; 5. Base of the STH “band”. These five possibilities are characterized briefly below, with a short critical evaluation of the present potential for global GSSP to define the base of a series and stage. (1), (2), and (3) rely on species of Oryctocephalidae – a group which underwent a remarkable radiation after the initial stage of a global transgression, which followed the complex terminal “Lower Cambrian” regressions, known as the “Hawke Bay Event” (Palmer and James, 1980). 3. TRILOBITE CORRELATION LEVELS 3.1. FAD of Oryctocephalus indicus Most ISCS Members momentarily seem to favor the FAD of Oryctocephalus indicus for defining the base of a global Middle Cambrian series (e.g., Sundberg et al., 1999; Zhao et al., 2001; McCollum and Sundberg, 2002; Zhao et al., 2004). Oryctocephalus indicus (Reed 1910) is a trilobite which was reported from the Tethyan Himalaya and South China as well as from western Laurentia. It thus appears to bracket three continents with a high precision. Problems lie in a perfect identification of even slightly distorted specimens. The type material was described by Reed (1910) as Zacanthoides indicus and comes from the Spiti area of the Tethyan Himalaya, northern India, and suffers from conspicuous tectonic deformation (see Jell and Hughes, 1997) so that in strict sense the species cannot be identified from other areas without certain reservations..
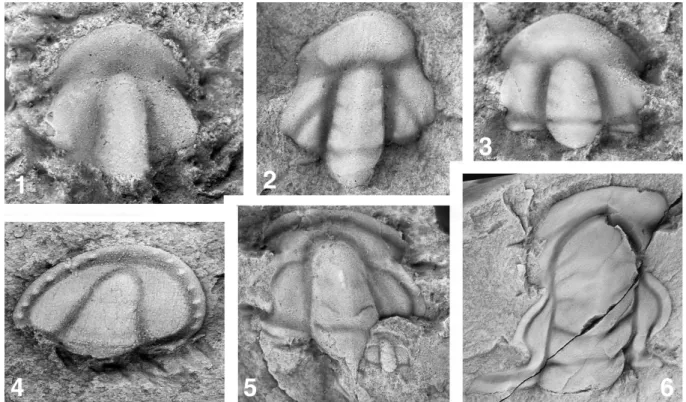
(4) 84. Gerd Geyer. Fig. 1. Trilobites of the Oryctocephalus indicus level. (1, 2) Oryctocephalus orientalis Saito 1901. (1) Internal mold of complete dorsal exoskeleton, GTB 24-5-6, ×6. (2) Internal mold of complete dorsal exoskeleton, GTB 20-5-1554, ×5. Oryctocephalus indicus level. From the Kaili Formation, Guizhou, South China. Specimens from the collections of Yuan et al. (2002), refigured with permission. (3, 4) Oryctocephalus indicus (Reed 1910). (3) Latex cast of external mold of cranidium, USNM 488953, ×5. (4) Latex cast of external mold of thorax and pygidium, USNM 488954, ×5.2. Specimens from the collections of Sundberg and McCollum (1997), Monola Formation, east side of Saline Valley, Nevada (by courtesy of F. Sundberg).. Reed (1910) described in the same article in which he introduced Zacanthoides indicus an “Oryctocephalus cf. reynoldsi” which was placed into synonymy with O. indicus by Jell and Hughes (1997) and almost certainly represents the same species. However, it illustrates the difficulties with determinations of the material so that the precise correlation from and into northern India a priori remains difficult. The best preserved material assigned to O. indicus comes from the Kaili Formation in southeastern Guizhou (Yuan et al., 2002) (Fig. 1). These excellent specimens from the Yangtze (or South China) Platform appear to differ slightly not only from the specimens from the Himalaya but also from the material recovered from the Emigrant Formation of the Split Mountain section and coeval strata of other sections in western Nevada (Sundberg and McCollum, 1997, 2003). As the key section in the Kaili Formation of Guizhou, the Split Mountain section was carefully studied, and very detailed range charts allow to reconstruct true (local!) ranges of the moderately diverse trilobites over four zones from the top of the Olenellus Biozone (traditional Bonnia-Olenellus Zone) to the Oryctocephalus indicus Zone (Sundberg and McCollum, 2003). In addition, the Split Mountain section includes other trilobites which might be helpful for interregional and even intercontinental correlation, such as Paraantagmus latus or Onchocephalites claytonensis. Paraantagmus latus was earlier described from the Paragraulos Zone of Huainan in the Anhui Province, China (Yuan and Li, 1999). Unfortunately, Paraantagmus latus is known from the Split Mountain section from only a single spot sample close to. the base of the newly introduced Amecephalus arrojosensis Biozone (just below the Oryctocephalus indicus Biozone). However, it is also known from the Monola Formation of the Saline Range, California. Microryctocara nevadensis ranges in Nevada from the Amecephalus arrojosensis Zone into the Oryctocephalus indicus Zone (Sundberg and McCollum, 2003) and is found in the Danzhaiaspis-Xingrenaspis Zone of the Kaili Formation in Guizhou (Yuan et al., 1997). Korovnikov (2001) suggested that Oryctocephalus reticulatus (Lermontova, 1940) as described from the YudomaOlenek facies region of the Siberian Platform is a junior synonym of O. indicus. At least if we follow the concept of a relatively large morphological plasticity of O. indicus, this interpretation would be correct and would enable a correlation with the Siberian Platform. Oryctocephalus reticulatus occurs in the Kounamkites Zone of the early (but not earliest!) Middle Cambrian Amgan Stage of the Siberian stratigraphic scheme. In addition, some Siberian specimens assigned to O. reticulatus by Yegorova et al. (1976, pl. 49, figs. 14–16) appear to represent O. americanus Sundberg and McCollum 2003, described from the Amecephalus arrojosensis Biozone in Nevada (F. Sundberg, pers. comm., March 2005). The morphologic plasticity of Oryctocephalus indicus needs to be clarified. Yuan et al. (2002) distinguished several subspecies of O. indicus in Guizhou, and Yuan and Zhao (2004) suggested that the material determined as O. indicus from Nevada represents another subspecies, which would slightly diminish its value for precise correlation. As.
(5) The base of a revised Middle Cambrian. suggested by F. Sundberg (communication on Lower-Middle Cambrian GSSP Working Group website, http://www.geology.ewu.edu/mccollum/workinggroup/correspondence.htm), the problems morphologic plasticity/local populations vs. distinction of subspecies needs to be clarified. In addition, Oryctocephalus indicus is known in Nevada only from a relatively thin pile of strata (ca. 6–8 meters; Sundberg and McCollum, 2003), whereas this species is described from 40–80 m of rocks in Guizhou. Thus, it needs to be solved (as suggested by F. Sundberg, communication on Lower-Middle Cambrian GSSP Working Group website, http://www.geology.ewu.edu/mccollum/workinggroup/correspondence.htm), preferable by means of eustatic sea-level changes, whether the strata in the two regions and whether these depict the true range of O. indicus. A most critical point is the subsequent appearances of the various species of Oryctocephalus. In the Split Mountain section, five species of Oryctocephalus have been identified. Oryctocephalus americanus occurs from the base of the Amecephalus arrojosensis Zone to about the base of the O. indicus Zone. Oryctocephalus orientalis is only known from a single horizon in the upper A. arrojosensis Zone as well as an unidentified species of Oryctocephalus. The first appearance of Orytocephalus indicus marks the base of the eponymous zone. Oryctocephalus orientalis was reported from South China and South Korea as well. However, in South Korea, O. orientalis occurs together with O. indicus, whereas in Guizhou O. orientalis occurs above O. indicus. It is yet unclear whether the species tend to occur at different times in the various regions, whether the ordinary regional biostratigraphies are unable to reveal the true ranges of the two species, or whether misidentifications lead to the different ranges. A further complication is the occurrence of Onchocephalites claytonensis Sundberg and McCollum 2003 in Nevada and Guizhou. This species occurs mainly in the Amecephalus arrojosensis Zone of western Nevada but ranges up into the O. indicus Zone (Sundberg and McCollum, 2003). Onchocephalites claytonensis was recently identified in limestones of the Tongren section, northeastern Guizhou, where it occurs only together with Oryctocephalus indicus (Yuan and Zhao, 2004). Its identified occurrence in Guizhou suggested that only the upper part of the O. indicus Biozone is represented in Nevada. However, F. Sundberg (“Comparison of the Onchocephalites claytonensis from China and those from Nevada”, communication on Lower-Middle Cambrian GSSP Working Group website, http://www.geology.ewu.edu/mccollum/workinggroup/correspondence.htm) provided a careful analysis of the material and suggested that the specimens from Guizhou represent a species different from O. claytonensis. It will be necessary to clarify these problems before the Oryctocephalus indicus level in any of the sections can be accepted as a GSSP. Summarized, the Oryctocephalus indicus level is possibly able to tie regions which are otherwise extremely difficult. 85. to correlate. However, the validity of the correlation needs to be confirmed by careful taxonomic analyses. Moreover, many other regions cannot be tied precisely by correlation from this level. It appears to be impossible to confidently identify the Oryctocephalus indicus level in such regions as western Gondwana, Avalonia, and Baltica. 3.2. FADs of Ovatoryctocara granulata and/or Kiskinella cristata Ovatoryctocara granulata was first described as Oryctocara (Ovatoryctocara) granulata from the Amgan of northern Siberia (Chernysheva, 1962, pl. V, figs. 6–8). It is a fairly common trilobite in the lower Amgan of the YudomaOlenek facies region where it defines the lowermost Amgan biozone and thus the base of the Middle Cambrian series. (It should be noted that the original concept of the Siberian stratigraphy had the Elankan as the lower part of the Amgan s. l., which was later transferred to the terminal Early Cambrian following a resolution of the All-Union Stratigraphic Workshop in 1983.) Well preserved and instructive material of O. granulata comes from the Nekekit River area (Yegorova et al., 1976), where the eponymous Nekekit River section is now the stratotype for the open-marine YudomaOlenek facies belt of the northern and northeastern Siberian Platform. Additional material of Ovatoryctocara granulata was described from South China (lowermost Kaili Formation of Guizhou, assigned to the Ovatoryctocara granulata-Bathynotus holopygus Zone of the Duyunian; Yuan et al., 2002) and the Avalonian southeastern Newfoundland (Fletcher, 2003). Material of Ovatoryctocara similar to Ovatoryctocara granulata is known from North Greenland (Ovatoryctocara sp. A of Blaker and Peel, 1997) (Fig. 2.1–2.3). Although this should not be a key point in the discussion, the FAD of Ovatoryctocara granulata has the advantage to lie at a level which agrees quite well with traditional boundary concepts so that there would be little change in many areas after selection of a GSSP for the (new) series boundary. It is even the index fossil for the basal zone of the Amgan and thus defines the base of the Middle Cambrian in the Siberian standard. As with Oryctocephalus indicus, the small individuals of Ovatoryctocara granulata are easily affected by tectonic distortion. Although such a tectonic deformation is most likely in southeastern Newfoundland, the specimens of Ovatoryctocara granulata from the uppermost Brigus Formation in the Easter Cove near Branch, St. Mary’s Bay, are quite well determinable because they were preserved in calcareous concretions (Fletcher, 1972, 1999, 2003). However, the material is very sparse (only three specimens identified to date) and only known from a single horizon at the very top of the Branch Cove Member so that its true range cannot be determined, and even the original horizon in this sec-.
(6) 86. Gerd Geyer. Fig. 2. Trilobites of the Ovatoryctocara granulata level. All from Henson Gletscher Formation, North Greenland. Specimens from Blaker and Peel, 1997, reproduced with permission. (1–3) Ovatoryctocara granulata Chernysheva 1962 (described as Ovatoryctocara sp. A by Blaker and Peel, 1997). All from GGU collection 218610, Locality 5, section northeast of the snout of Henson Gletscher. (1) Pygidium, internal mold, dorsal view, MGUH 23480, ×24. (2) Cranidium, internal mold, dorsal view, MGUH 23478, ×23. (3) Cranidium, internal mold, dorsal view, MGUH 23479, ×24. (4–6) Pagetides elegans Rasetti 1945. All from GGU collection 298553, Loc. 1 in southern Freuchen Land. (4) Cranidium, partially exfoliated, dorsal view, MGUH 23174, ×12. (5) Pygidium, partially exfoliated, dorsal view, MGUH 23185, ×15. (6) Pygidium, internal mold, dorsal view, MGUH 23186, ×15.. tion appears to be exploited now. Even more than Oryctocephalus indicus, Ovatoryctocara granulata can be put into a stratigraphic context with auxiliary key taxa such as Kiskinella cristata Romanenko and Romanenko 1962 and Peronopsis roddyi Resser and Howell 1938. Kiskinella cristata, first described from the Kiska River area in the Dolgy Mys Mountains of the High Altay (Romanenko and Romanenko, 1962; Chernysheva, 1971), also occurs in southeastern Newfoundland (Fletcher, 2003) with well preserved specimens that enable a confident determination. Kiskinella cristata was found immediately above O. granulata in the Easter Cove section (see Fletcher, 2003). Unlike Ovatoryctocara granulata, is known from a number of samples in the lower Chamberlain’s Brook Formation. It remains uncertain whether these occurrences portray its true stratigraphic range but its lowest occurrence is identical with that of O. granulata. A similar species, Kiskinella decora Yegorova 1967, occurs in the lower Amgan Kounamkites Zone of the Daldyn River basin in of the northern Prianabar region on the Siberian Platform (Yegorova and Savitskiy, 1969). Peronopsis roddyi Resser and Howell 1938, also known as Eoagnostus roddyi and originally described from the Lower Cambrian Kinzers Shale of Pennsylvania, would be an extremely helpful index fossil. Peronopsis roddyi is not only known from the Kinzers Shale (see Rasetti and Theokritoff, 1967) but was identified from North Greenland. (together with material identified as Oryctocephalus vicinus and Arthricocephalus chauveaui; Blaker and Peel, 1997) (Fig. 4.1–4.3) and recently from southeastern Newfoundland (Fletcher, 2003) so that it would permit a correlation from Laurentia into western Avalonia. In Newfoundland, its first occurrence in the Easter Cove Section coincides with that of Ovatoryctocara granulata but P. roddyi is also present in the overlying Easter Cove Member of the lower Chamberlain’s Brook Formation (Fletcher, 2003). Logically, P. roddyi would better portray a sincere biozone in southeastern Newfoundland than does O. granulata. Nevertheless, distinct differences in the figured material of P. roddyi from different areas indicate that it requires careful restudy before the identification can be accepted. Unfortunately, the many species which co-occur with Ovatoryctocara granulata in the Kuonamka Formation of the Nekekit River area are usually of little help for precise intercontinental correlation although Oryctocara ovata, O. angusta, Pagetia horrida and Pagetides spinosus are noteworthy. Pagetia horrida, however, appears to indicate the severe problems that arise from different facies: It is known to occur in the Anabaraspis Zone of the upper Toyonian in the Yudoma-Olenek Facies region, whereas it occurs in the lower Amgan in the Anabar-Sinyaya Facies region (see Astashkin et al., 1991; Korovnikov, 2001). In addition, the O. granulata Zone yields “Paradoxides” eopinus, the oldest Paradoxides species of the Siberian Platform..
(7) The base of a revised Middle Cambrian. Pagetides rjonsnitzkii (Lermontova 1940) was first described from the (at that time) terminal Lower Cambrian of Dzhagdi, eastern Siberian Platform, and found to occur together with the Ovatoryctocara material and with Oryctocephalus vicinus Chernysheva 1962 in the Henson Gletscher Formation of Peary Land, North Greenland. The Siberian material of Oryctocephalus vicinus occurs in the lower Amgan Ovatoryctocara Zone of the Kuonamka Formation on the Olenek Uplift, but the species has a rather long range in the Henson Gletscher Formation, with a lowest stratigraphic occurrence below(!) Arthricocephalus (see below) and ranging up to the top of the formation where it occurs together with Pagetides elegans. Again, there are minor differences in morphology between the Siberian specimens of Pagetides rjonsnitzkii and the material from Greenland (see discussion in Blaker and Peel, 1997, p. 43). However, the correlation between the Ovatoryctocara level in Greenland and the lowermost Amgan in Siberia is largely reconfirmed by the occurrence of Pagetides elegans and Peronopsis roddyi above the strata with Ovatoryctocara and those with Arthricocephalus in Peary Land (see below). Pagetides elegans characterizes the upper part of the “Pagetides Fauna” of the Taconic sequence where it occurs with the youngest species of Olenellus (Rasetti, 1967) as it does in North Greenland (Fig. 2.4–2.6). Summarized, a correlation from Siberia into western Avalonia is possible by means of Ovatoryctocara granulata, with a subsequent possibility to correlate from there into western Gondwana. However, southwestern Newfoundland appears to be unsuitable for a GSSP because of a relatively strong structural deformation. In addition, the occurrences in Siberian and southeastern Newfoundland most probably do not depict true ranges but are triggered by changes in facies. A correlation into Laurentia at this level is only possible if the occurrences of Peronopsis roddyi in Newfoundland can be verified. 3.3. FAD of Arthricocephalus chauveaui Arthricocephalus chauveaui Bergeron, 1899, another oryctocephalid trilobite, was first described from the present-day Balang Formation of northeastern Guizhou (Fig. 3.7–3.9). It was subsequently found in other parts of the Yangtze (or South China) Platform, probably in North Greenland (Blaker and Peel, 1997) (Fig. 3.1–3.4), Avalonian Newfoundland (Fletcher 2001, 2003), and Siberia (described as Oryctocarella sibirica Tomashpolskaya and Karpinskiy 1962 and partly as Oryctocara spp.; see Blaker and Peel, 1997, for synonymy and discussion). Its occurrences have a high promise for relatively precise intercontinental, or even global correlation. However, the identity of Arthricocephalus chauveaui has been matter of recent debates, and its FAD is in a stratigraphic position that would be well in traditional Lower Cambrian strata in most regions.. 87. As it is the case for most oryctocephalid trilobites, even a slight deformation notably affects the morphology of Arthricocephalus chauveaui. Particularly in South China, small differences in morphological details led to the introduction of numerous different species under Arthricocephalus. It remains open, how far this taxonomic distinction is hypertrophied. Arthricocephalus duyunensis, A. horridus Qian and Lin 1980, and A. granulus Chien and Lin 1977 were already synonymized with A. chauveaui. However, it is unclear for reasons of preservation whether species such as A. pulchellus Zhang and Qian, 1980, A. xiaosaiensis Qian 1980, A. pectines Qian 1980, A. tongrenensis Yin 1978, A. intermedius Chien and Lin 1974, or A. xinzhaiheensis Chien and Lin 1974 (see Zhang et al., 1980 for complete list) are well defined or should be placed in synonymy, too, although they partly present distinct differences between them and the remaining species. A recent analysis of the ontogeny of A. chauveaui from the Balang Formation of Guizhou (McNamara et al., 2003) illustrates the morphologic plasticity of the species but also proves that certain character states seen in figured material obviously are not characteristic for this species. In this context, the material from the Henson Gletscher Formation of northern Greenland (Blaker and Peel, 1997) needs careful reinvestigation. Nevertheless, McNamara et al. (2003) confirmed certain evolutionary trends in the Chinese material of Arthricocephalus so that at least four species of them can be placed in an evolutionary sequence. This sequence with Arthricocephalus chauveaui – A. xinzhaiheensis (with its subjective synonym A. intermedius) – A. balangensis – A. pulchellus portrays their stratigraphic appearance and a series from more to less plesiomorphic characters. The later species shares features with the genus Tonkinella suggesting a close phylogenetic relationship. It should further be noted that A. xinzhaiheensis, A. balangensis, and A. pulchellus represent the further subgenus Arthricocephalites, which was raised to generic level. In South China, Arthricocephalus chauveaui occurs just below the latest species of Redlichia (see Zhao et al., 2001) which mark the top of the traditional Lower Cambrian in China. As noted by Fletcher (2003), the overlying lowermost Middle Cambrian Xystridura-bearing strata at Yaxian, Hainan Province, bear specimens of Kiskinella luoyacunnensis (Lin and Jago, 1993), a species close to K. cristata. Peronopsis roddyi is found to accompany Arthricocephalus chauveaui in the lower Henson Gletscher Formation in North Greenland (Blaker and Peel, 1997) (Fig. 4.1–4.3) which would suggest that its stratigraphic occurrence matches more-or-less that of Ovatoryctocara granulata. Other characteristic trilobite species of this association are Haliplanktos jishouensis (Zhou in Zhou et al., 1977), Ogygopsis typicalis (Resser 1939) and Lancastria plana (Tomashpolskaya 1960), whereas Pagetides elegans Rasetti 1945 first appears slightly above A. chauveaui. Haliplanktos jishouen-.
(8) 88. Gerd Geyer. Fig. 3. Trilobites of the Arthricocephalus chauveaui level. Specimens from Blaker and Peel, 1997, reproduced with permission. (1–4), (7–9), Arthricocephalus chauveaui Bergeron 1899. (1–4) From Henson Gletscher Formation, Loc. 1 in southern Freuchen Land, North Greenland. (1) Cranidium, internal mold, dorsal view, MGUH 23473 from GGU collection 315096, ×16. (2) Complete dorsal exoskeleton of immature individual, dorsal view, MGUH 18225 from GGU collection 315095, ×8. (3) Cranidium, dorsal view, MGUH 23472 from GGU collection 315095, ×19. (4) Meraspid pygidium, dorsal view, MGUH 23474 from GGU collection 315099, ×11. (7–9) Lectotype material of Bergeron (1899) (see Lane et al., 1988) from Balang Formation, north of Tongren, northeastern Guizhou, South China, EM 90001b, collection Université Claude Bernard, Lyon, all ×12. (7) Articulated thorax and pygidium (dorsal view) plus disarticulated cranidium (ventral view) and hypostome (ventral view) of supposedly the same individual, lectotype. (8) Two complete exoskeletons, dorsal view, paralectotypes. (9) articulated thorax and pygidium, distinctly flattened, dorsal view, paralectotype. (5, 6) Haliplanktos jishouensis (Zhou in Zhou et al., 1977). Both from Henson Gletscher Formation, North Greenland, Loc. 1 in southern Freuchen Land. (5) Articulated thorax and pygidium of fairly complete dorsal exoskeleton, dorsal view, MGUH 23475 from GGU collection 315096, ×9. (6) Complete dorsal exoskeleton, dorsal view, MGUH 23476 from GGU collection 315099, ×7..
(9) The base of a revised Middle Cambrian. 89. Fig. 4. Trilobites of the Arthricocephalus chauveaui level. All from Henson Gletscher Formation, North Greenland. Specimens from Blaker and Peel, 1997, reproduced with permission. (1), (2), (4), (5), (8), from Loc. 1 in southern Freuchen Land; (7), (9), from Loc. 2 at western Henson Gletscher; (3), (6), from Loc. 6 in Løndal. (1–3) Peronopsis roddyi (Resser and Howell 1938). (1) Immature cephalon with particularly well developed dorsal furrows, dorsal view, MGIH 23137 from GGU collection 315107, ×24. (2) Pygidium, internal mold, dorsal view, MGIH 23143 from GGU collection 315107, ×20. (3) Incomplete pygidium, dorsal view, MGIH 23139 from GGU collection 271750, ×17. (4–6) Ogygopsis virgata (E. Romanenko 1962). (6) Incomplete cranidium, dorsal view, MGIH 23321 from GGU collection 315097, ×7.5. (5,6) Pygidium, partly exfoliated, dorsal and right lateral views, MGIH 23348 from GGU collection 301343, ×6. (7–9) Lancastria plana (Tomashpolskaya 1960). (7) Immature cranidium, partly exfoliated, dorsal view, MGIH 23507 from GGU collection 271485, ×9. (8) Cranidium, dorsal view, MGIH 23510 from GGU collection 301341, ×4. (9) incomplete cranidium, dorsal view, MGIH 23506 from GGU collection 271485, ×8.. sis (Fig. 3.5, 3.6) was first described as Arthricocephalus (Arthricocephalites) jishouensis from South China (Zhou et al., 1977), where it also co-occurs with Arthricocephalus chauveaui. Specimens of this species were even erroneously determined as Arthricocephalus chauveaui by Yegorova et al. (1963) and Lu et al. (1965). Ogygopsis typicalis was originally described as Taxioura typicalis from the socalled “Ptarmigania strata” or “Ptarmigania fauna” of the basal Langston Formation of northeastern Utah and southeastern Idaho, where it was believed to belong to the early Middle Cambrian Albertella Zone whereas it occurs in the upper Olenellus Zone of Greenland. A special case as a stratigraphic tool in this level is Lan-. castria plana, first described as Paraoryctocephalops plana from the Dolgiy Mys Range, Kuznetsk Ridge, in the High Altay Mountains (Tomashpolskaya in Khalfin, 1960). It occurs in its type area together with Arthricocephalus chauveaui as it does in North Greenland (Fig. 4.7–4.9). However, in North Greenland its range extends into the level with Ovatoryctocara granulata. In this context, the sections of North Greenland strongly suggest that the chronometric difference between both levels is less dramatic than believed by some authors. A similar species was described under Goldfieldia kunshanensis Zhang and Zhou 1985 from the southern Jiangsu Province of South China, where it occurs above the species of Arthricocephalus (Zhang and Zhou,.
(10) 90. Gerd Geyer. Fig. 5. Trilobites of the Cephalopyge zone. All from Jbel Wawrmast Formation, southern Morocco. (1, 2) Cephalopyge notabilis Geyer 1988. (1) Complete dorsal exoskeleton, latex cast of external mold, Amouslek, western Anti-Atlas, SMF 41781, ×5. (2) Pygidium, dorsal view, Ourika Wawrmas, central Anti-Atlas, ×8. (3) Acanthomicmacca neltneri Hupé 1953, cranidium, dorsal view, Ourika Wawrmas, central Anti-Atlas, ×4. (4) Hamatolenus (Hamatolenus) marocanus (Neltner 1938), lectotype, incomplete dorsal exoskeleton, Wirgane, western High Atlas, MNHN R. 50831, ×1.3. (5) Schistocephalus ornatus Geyer 1998, holotype, partial cranidium, dorsal view, Itsiar, western High Atlas, PIW 97II146a, ×2.5. (6) Kingaspidoides brevifrons (Hupé 1953), holotype, cranidium, dorsal view, Ourika Wawrmas, central Anti-Atlas, MNHN R. 50829, ×3. (7) Protolenus (Protolenus) interscriptus Geyer 1990, holotype, latex cast of internal mold, dorsal view, Afourigh, Ounein area, western High Atlas, SMF 41584, ×2. (8) Cobboldites tichkaensis Geyer 1988, paratype, pygidium, largely exfoliated, Tizi n’Tichka, central High Atlas, SMF 41676, ×4.. 1985), but is associated with “Changaspis taihuensis”, which represents another Goldfieldia. Goldfieldia pacifica Palmer 1964, the type species of the genus, was originally described from the Saline Valley Formation of Esmeralda County. Ogygopsis batis appears to be identical with O. sibirica (E. Romanenko 1960), described from the upper Toyonian of the Suyarik River area of the High Altay Mountains (Chernysheva, 1971; Yegorova et al., 1976). In addition, Ogygopsis batis has also been identified from the upper Toyonian Anabaraspis splendens Zone of the Elanka Formation in the Elanka section of the Lena region (Yegorova et al., 1976). Ogygopsis virgata (E. Romanenko 1962), known from a spot sample just below Arthricocephalus chauveaui in the Henson Gletscher Formation of North Greenland (Fig. 4.4–4.6), was reported from the Harkless Formation of western Nevada and again from the lowermost Amgan Suyarik “Gorizon” of the Suyarik River area in the High Altay (Chernysheva, 1971) and is at least close to O. antiqua of the Hillard Limestone of Alaska (Palmer, 1968). Arthricocephalus chauveaui and key species in its assemblage may be able to tie regions which are otherwise difficult to correlate. However, confident identification of the species is often difficult so that the true ranges of the spe-. cies are probably not yet known. A major disadvantage of this level is that there are few (if any) further species to back or extend the correlations. In addition, the level lies in traditional “Lower Cambrian” strata. 3.4. Base of the Acidiscus-Cephalopyge Assemblage “Zone” Locally rich associations of trilobites are termed herein the Acidiscus-Cephalopyge assemblage “zone”. It unites essentially two faunal assemblages described from different regions. The first is the Acimetopus-Acidiscus assemblage of Warwickshire, central England (e.g., Purley Shales; Rushton, 1966) and Taconic Laurentia (Rasetti, 1966, 1967; Rasetti and Theokritoff, 1967). Its key species (in the so-called Acimetopus Fauna) include mostly eodiscoids such as Acidiscus theristes Rushton 1966, Acimetopus bilobatus Rasetti 1966 and other Acimetopus spp., Bathydiscus spp. (including Oodiscus; see Fletcher, 2003), and Tannudiscus (more than 25 eodiscoid species according to Rasetti 1966, 1967; and 9+ species of true olenellids). Unfortunately, the biostratigraphy of the immediately overlying strata is incompletely known from Warwickshire (where the sequence appears to be partly condensed; see Rushton, 1966), and the very productive samples of the Taconic sequences usually.
(11) The base of a revised Middle Cambrian. came from occasional small outcrops that mostly did not permit recognition of stratigraphic ranges of the fossils. The second assemblage is a relatively rich and variably developed association of protolenoid, ellipsocephaloid, and earliest solenopleurid trilobites with some eodiscoids, known from areas such as Morocco, Spain, the Holy Cross Mountains of southwestern Poland, the Welsh Borderlands, North Wales, and southeastern Newfoundland. It includes such key species as Condylopyge eli Geyer 1998, Cephalopyge notabilis Geyer 1988, Cobboldites spp., Hamatolenus spp., Latoucheia spp., Kingaspidoides spp., Schistocephalus spp., Acadoparadoxides spp. (A. nobilis, A. mureroensis, A. harlani). This second assemblage is best studied from the Jbel Wawrmast Formation of the Moroccan Atlas ranges (Geyer, 1983, 1998, Geyer and Landing, 1995, Geyer et al., 1995) (Fig. 5) where it can be investigated over a stretch of almost 400 kilometers with rich association and notable variations in lithofacies and biofacies that allows a confident determination of true biostratigraphic ranges for this regions. Both assemblages can be tied in southwestern Newfoundland. Fletcher (1972, 2003) identified a number of key species to occur together, such as Acidiscus theristes, Cephalopyge notabilis, Condylopyge eli, and Hamatolenus (Fletcher, 2003). These species have their acme in the upper Brigus Formation which Fletcher (2003) assigned to the Cephalopyge notabilis Zone so that this assemblage would be slightly older than the Ovatoryctocara granulata chron. The “Protolenus limestone” of Comley in the Welsh borderland shares Latoucheia and Cobboldites with the Cephalopyge notabilis Zone of southern Morocco. Difficulties exist with the identification of some specimens from that region. For example, some specimens of Fletcher [2003, pl. 2, figs. 23, 24, and 26 (only)] identified as Cephalopyge notabilis do not represent this species because they have marginal spines whereas the cephalic and pygidial margins of C. notabilis are smooth. This may result in the problems that the newly established zones for southwestern Newfoundland (Fletcher, 2003) are locally reversed (such as in the Redland section of Fletcher, 2003, fig. 5) by comparison to southern Morocco (Geyer, 1990a) although other species clearly parallel the “Moroccan-type” biostratigraphic scheme; one example being the occurrence of Orodes howleyi (Walcott 1889) in the Redland Cove Member of the Brigus Formation suggesting a lower Hupeolenus Zone as does Orodes schmitti Geyer 1990 in Morocco. Key species from these assemblages occur in other regions and permit a fairly reliable correlation so that the variable associations of mostly miomerid trilobites are not only known from the mentioned regions of eastern Laurentia, western and eastern Avalonia, West Gondwana, but also from the Tyan-Shan, the Sayan-Altay Fold Belt, the Primorye region of eastern Siberia, South China, Australia, and Antarctica. As an example, the assemblage allows a. 91. correlation via the Schistocephalus spp. into the AnabarSinyaya facies region, a transitional facies region that crosses the Siberian Platform from the northeast to the southwest. Schistocephalus antiquus characterizes the lowermost Middle Cambrian biozone of this Anabar-Sinyaya facies, together with S. juvenis. This coeval species Schistocephalus juvenis occurs in the Burj Formation of Jordan (Rushton and Powell, 1998) within an assemblage that most probably represents an equivalent to the upper Cephalopyge Zone (Geyer and Landing, 2000), a suggestion that is confirmed by Mypsolenites in equivalent strata of Israel and Morocco (Geyer and Landing, 2004). Due to numerous facies changes, sections of the Anabar-Sinyaya facies region usually are affected by multiple minor unconformities such as in the Elanka Formation of the Elanka section, the regional stratotype for the Lower-Middle Cambrian boundary in the Anabar-Sinyaya facies belt (e.g., Korovnikov, 2004). It should also be mentioned that undescribed material of Syspacephalus occurs in the Cephalopyge notabilis Zone of the Moroccan Atlas Ranges (G. Geyer, unpubl. data), whereas this genus has its acme in the Amecephalus arrojosensis Zone of western Laurentia (but a very long range if used in a broad generic concept as suggested earlier). The Acidiscus-Cephalopyge assemblage proves again that Olenellus persisted into strata which are coeval with the Cephalopyge notabilis bearing strata. Paradoxides (s.l.) was already a frequent faunal element in these strata in North Africa and southwestern Europe. However, Olenellus persisted even into strata coeval with those with Ovatoryctocara granulata and Kiskinella cristata, which bear the frequent large Acadoparadoxides harlani of southeastern Newfoundland (Fletcher, 2003). In addition to the relatively numerous and easily determinable species which permit a correlation into most other regions, the position of the Acidiscus-Cephalopyge assemblage ‘zone’ is at a supposedly global transgressive systems tract which would permit a corroboration of the level by sequence stratigraphic methods. However, the level also has some disadvantages. Its exact position is often difficult to pinpoint because there is not one single species of global distribution, and its correlation is often impeded by relatively scarce occurrences of the key taxa. Moreover, the base of the Acidiscus-Cephalopyge assemblage ‘zone’ often falls into clastic facies with a resulting sparse fossil content. It should be noted that this clastic facies resulting from a transgressive reworking is locally misinterpreted as the regressive facies which often sits below, sometimes separated only by a cryptic unconformity. The position of the Acidiscus-Cephalopyge assemblage ‘zone’ roughly coincides with that of Ovatoryctocara granulata and therefore agrees quite well with the base of the Middle Cambrian in Baltica and various other regions so that there would be little change in many areas after selection of a GSSP for the series boundary (Geyer and Landing,.
(12) 92. Gerd Geyer. 2004). It should be noted, however, that the Cephalopyge notabilis Zone in Morocco correlates with the traditional Lower Cambrian Protolenus limestone of Comley, England, and is not at the base of the West Gondwanan Celtiberian Series and Agdzian Stage because Acadoparadoxides is known from the older Hupeolenus Zone. 3.5. Base of the STH “Band” Associations with such trilobites as Serrodiscus bellimarginatus, Triangulaspis annio, T. schucherti and Hebediscus attleborensis are located in the traditional Lower Cambrian in regions such as western Avalonia (southeastern Newfoundland; e.g., Fletcher, 2003), eastern Avalonia (central England; Rushton, 1966), Taconic Laurentia (Lochman, 1956; Rasetti, 1967), West Gondwana (Spain, Morocco, Sardinia; e.g., Richter and Richter, 1941; Hupé, 1953; Sdzuy, 1962; Rasetti, 1972; Geyer, 1983, 1990a; Gil Cid, 1988, 1991; Pillola, 1991; Álvaro and Liñán, 1997; Geyer and Landing, 2002) (Fig. 6), the Altay-Sayan Fold Belt (e.g., Repina et al., 1964), various parts of the Siberian Platform (e.g., Demokidov et al., 1959; Demokidov and Lazarenko, 1959;. Datsenko et al., 1968; Yegorova and Savitskiy, 1969; Savitskiy et al., 1972; Repina et al., 1974) and the Russian Far East region (Okuneva and Repina, 1973); with restrictions even in Laurentian Greenland and South Australia. Nevertheless, this faunal band is generally strongly fossiliferous because its animals existed earlier than the globally detectable regression that masks the successions in most of the Cambrian continents by a huge input of clastic material. As a result, auxiliary trilobite taxa exist in this level and enable a correlation into other regions. In Western Gondwana, this fauna includes other species of Hebediscus, Triangulaspis (such as T. vigilans and T. zirarii), Mallagnostus llarenai, Serrodiscus coloi, Serrodiscus silesius (from eastern Germany), saukiandiids, and early ellipsocephalines such as Rinconia spp. (from Spain and Morocco) (Fig. 6). A correlation into the Avalonian Callavia Zone is confirmed by the occurrence of this genus just below and Atops cf. calanus within the STH “band” in the Moroccan Anti-Atlas (Geyer and Landing, 2002). In the “Taconic sequence” of the New York State and adjacent areas, the assemblage was earlier termed the “Elliptocephala asaphoides Fauna” (Lochman, 1956) and includes. Fig. 6. Trilobites of the STH band. Examples from Western Gondwana (1–3, 5, 6 from Morocco, 4 from Germany). (1) Hebediscus lemdadensis Geyer 1988, cranidium, dorsal view, Sectigena Zone, Lemdad Formation, Lemdad Syncline, Section Le I, SMF 41733, ×14. (2) Sectigena intecta Geyer 1990, holotype, cranidium, latex cast of external mold, dorsal view, Sectigena Zone, Issafen Formation, Issafen Syncline, Section Ida ou Drif/Jbel Tasousekht, SMF 52846, ×7. (3) Berabichia vertumnia Geyer 1990, cranidium, dorsal view, Sectigena Zone, Lemdad Formation, Lemdad Syncline, Section Le I, SMF 41865, ×6. (4) Serrodiscus silesius Richter and Richter 1941, cephalon, internal mold, dorsal view, Charlottenhof Formation, Lusatiops Member, Serrodiscus beds, Quarry no. 3, Niederludwigsdorf, eastern Germany, SUS 402, ×6. (5) Rinconia issafenensis Geyer 1990, two cranidia, dorsal view, Sectigena Zone, Issafen Formation, Issafen Syncline, Section Ida ou Drif/Jbel Tasousekht, IGR 19345a-b, ×7. (6) Longianda termieri Hupé 1953, distorted partial cranidium, dorsal view, Issafen Formation, Issafen Syncline, MNHN (unregistered), ×1.7..
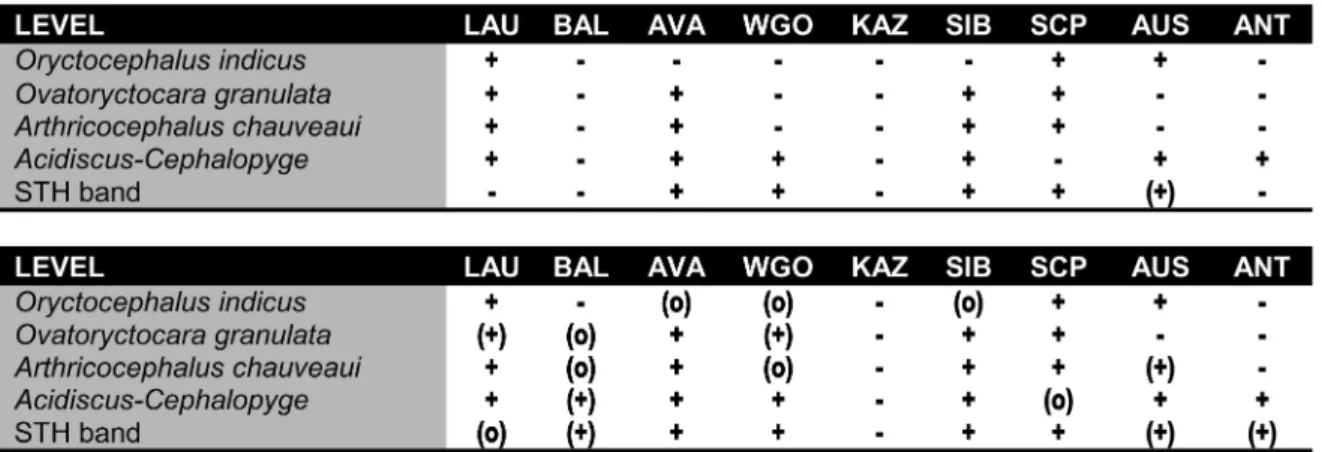
(13) The base of a revised Middle Cambrian. such species as Calodiscus meeki (Ford 1876), C. schucherti (Matthew 1896), and Serrodiscus speciosus (Ford 1873). Mallagnostus llarenai (Richter and Richter 1941), formerly dealt with as Ladadiscus llarenai, is not only known from the Sierra Nevada of southern Spain (Sdzuy, 1962) but also from the lower Purley Shales of Warwickshire, where it occurs together with Serrodiscus bellimarginatus (Rushton, 1966) and thus brackets Western Gondwana with Avalonia. Elements of this fauna occur in Siberia in the Bergeroniellus gurarii to B. asiaticus zones of the Botoman Stage (e.g., in the Buom Formation of the southwestern Prianabar region, in the Erkeket Formation of the Olenek Uplift, and in equivalent strata of the northwestern Siberian Platform), in the West Sayan Mountains (Obruchev “Gorizon”, e.g., Repina et al., 1964), and in the Far East region (Dmitriev strata; Okuneva and Repina, 1973); in Avalonian Newfoundland in the top of the Callavia Zone and the lower Hupeolenus Zone sensu Fletcher (2003); in Greenland in the middle Olenellus Zone (Kap Troedson Formation and lower Aftenstjernesø Formation; Blaker and Peel, 1997); and equivalent strata of Spitsbergen (Slaklidalen Formation; Major and Winsnes, 1955). A number of identifications require reconsideration (e.g., Hebediscus attleborensis as described from the lowermost Botoman of the Parfen-Yurakh and Kyunkyuy Rivers, Siberia; see Yegorova and Savitskiy, 1969). Other problems concerning the STH band and its stratigraphic precision are the clarification and calibration of various species of Serrodiscus and a re-evaluation of the Acanthomicmacca clade. Using this Serrodiscus-Triangulaspis-Hebediscus band (STH band) would allow a reasonably precise global correlation and offer the opportunity to calibrate it with physical methods. However, the key taxa are known from relatively few Cambrian areas; the associations are often paucispe-. 93. cific; and its key species tend to be difficult to recognize. The level often sits in a slightly to distinctly regressional development with a clastic input, often with hiatuses, and with associations which show little variation in biofacies. In addition, choosing the STH band as a global marker would force a departure from the traditional level(s) of the LowerMiddle Cambrian boundary so that new series names would be needed. The level is so much earlier than most of the other levels discussed above that it may be a candidate for a GSSP for the base of a terminal “Lower Cambrian” stage. 4. GLOBAL CORRELATION OF THE TRADITIONAL LOWER-MIDDLE CAMBRIAN BOUNDARY INTERVAL The five correlation levels characterized by trilobites or trilobite association all suffer from certain deficiencies. A common feature is that the stratigraphic ranges of key species are not sufficiently known to guarantee a precise recognition of the desired first occurrence in a studied section. Accordingly, numerous regions could only be tied tentatively to a potential GSSP (Fig. 7). In addition, problems of confident identification of the index species are particularly due for oryctocephalid species because of their small size (and resulting frequent deformation), their minimal differences between related species, and their morphologic plasticity. Most of the correlation levels discussed above are only recognizable with sufficient precision from a majority of Cambrian continents whereas key faunal elements are absent on the others. As a result, a fine-scale correlation of the Lower-Middle Cambrian boundary interval on a global scale is impossible and a sufficiently reliable and wellfounded GSSP for a series boundary is difficult in this moment.. Fig. 7. Momentary possibilities for fine-scale correlation of relevant levels for defining the base of a serious in the Lower-Middle Cambrian boundary interval on important Cambrian continents. Upper table indicates direct correlation by means of key taxa. Lower table shows possibilities of correlation when auxiliary taxa for interregional and intercontinental correlation are included. +, exact or apparently exact correlation possible; (+), more-or-less exact correlation may be possible; (o), approximate or tentative correlation may be possible; -, sufficiently exact correlation not possible momentarily. LAU, Laurentia; BAL, Baltica; AVA, Avalonia; WGO, West Gondwana; KAZ, Kazakhstania; SIB, Sibiria; SCP, South China Platform; AUS, Australia; ANT, Antarctica..
(14) 94. Gerd Geyer. Fig. 8. Correlation table of the Lower-Middle Cambrian boundary interval for selected key areas. Grey-scale bands indicate (from top to bottom) stratigraphic positions of the Oryctocephalus indicus level, the Ovatoryctocara granulata and/or Kiskinella cristata level, the Arthricocephalus chauveaui level, the Acidiscus-Cephalopyge assemblage zone, and the STH “band”. Characters indicate occurrences of Arthricocephalus chauveaui (Ac), Acidiscus theristes (At), Cephalopyge notabilis (C), relevant Hebediscus spp. (H), Kiskinella cristata (Kc), Ladadiscus llarenai (Ll), Ovatoryctocara granulata (Og), Oryctocephalus indicus (Oi), relevant Schistocephalus spp. (Sc), relevant Serrodiscus spp. (S), Triangulaspis (T), and Xystridura spp. (X).. Although a precise correlation between the different faunal realms is exceedingly intricate at the present state of knowledge, a sufficiently precise correlation can be achieved by a two-step procedure. The first step is a detailed correlation between neighboring regions. None of the levels discussed above is readily identifiable in all of the important areas. However, scrutinized application of the available data will allow a fairly precise correlation of regional biostratigraphic and lithostratigraphic units. The second step is a consequence of the first and results in a combination of the trilobite levels discussed under 3.1 through 3.4. This procedure shows a promise for intercontinental correlation that can be used as a global framework with adequate accuracy to serve normal correlation purposes. Early attempts by Lochman-Balk and Wilson (1958) and successive accumulation of additional information from classical areas and new records from additional regions with detailed profiles lead to a number of correlation tables all with new information and important supplementary data (e.g., Geyer, 1983, 1990b; Rozanov and Sokolov, 1984; Geyer and Palmer, 1995; Zhuravlev, 1995; Blaker and Peel, 1997; Geyer, 1998; Yuan and Li, 1999; Geyer and Shergold, 2000; Fletcher, 2001, 2003, Shergold and Geyer, 2003) although most of them were restricted to major Cambrian areas or the relevant faunal realms. Not surprisingly, this system of modifying existing correlation tables not only lead to tremendous changes against the earliest concepts but also to an approximation of certain correlation levels so that theoretically the ideal correlation may be calculated even by geometric means. This is true at least for the major areas although randomly studied regions tend to vacillate in the positions of their chronostratigraphic boundaries depending. of certain schools of research. Improved insights into (1) the relationships between different bathymetric regimes and (2) biofacies resulting from differentiations of the temperatures of the sea-water led to enhanced correlation. Nevertheless, migrating facies belts are not yet sufficiently identified so that corrections will be necessary in future. Even the polar wander path of the later Early to Middle Cambrian is difficult to reconstruct and subject of debates (e.g., Kirschvink et al., 1997) resulting in different interpretation of cool-water, offshore vs. warm-water, nearshore assemblages. Nonetheless, not only has it become clear that the traditional Lower-Middle Cambrian boundaries in the different regions have different chronologies; regional faunal associations with obvious “Lower Cambrian” or “Middle Cambrian aspects” are now known to have been misleading. The “Protolenus fauna” of Avalonia has its counterpart in Protolenus- and Latoucheia-bearing strata of the Cephalopyge Zone of southern Morocco, which is rich in Acadoparadoxides and solenopleurids and thus of Middle Cambrian aspect. The Anabaraspis fauna of the upper Toyonian of Siberia was that to have a clear Lower Cambrian aspect, with Anabaraspis being the predecessor of Paradoxides (s.l.) which in Siberia first occurs in the overlying basal Amgan. However, it can now be shown that the first Paradoxides even predates the Siberian Anabaraspis. (In this context, it should be noted that Anabaraspis splendens ranges up well into the lower Amgan in the Kharaulakh Mountains.) On a global scale, archaeocyaths have a clear acme in the lower and middle part of the trilobite-bearing Early Cambrian. However, species such as Tegerocyathus greenlandicus and Archaeocyathus pearylandensis occur in Greenland.
(15) The base of a revised Middle Cambrian. in the Paralleldal Formation, which represents the upper part of the Olenellus Zone (Debrenne and Peel, 1986), and thus in a stratigraphic interval which is under discussion herein. In addition, other species of Tegerocyathus occur in the coeval Obruchev “Gorizon” of the Altay-Sayan Fold Belt and in the coeval Sanashtykgol “Gorizon” of the Russian Far East region as well as in the Elanka Formation of the Siberian Platform. The most severe difficulty is distinguishing between true, regional, or facies-controlled occurrences of key taxa. As an example, Oryctocephalus reticulatus (suggested as a junior synonym of O. indicus) appears to first occur in the middle Kounamkites Zone of the Siberian Amgan, whereas Ovatoryctocara granulata is already found at the base of the underlying Oryctocara Zone (e.g., Astashkin et al., 1991; Korovnikov, 2001). In South China, the FAD of Ovatoryctocara granulata appears to briefly predate that of O. indicus, and both species appear to coexist (e.g., Yuan et al., 2002). In western Laurentia, possible specimens of Ovatoryctocara granulata are described from the Amecephalus arrojosensis Zone of western Nevada, where they predate (briefly?) Oryctocephalus indicus of the O. indicus Zone above (e.g., Sundberg and McCollum, 2003). However, indirect correlation appears to indicate that the Nevadan O. indicus Zone starts earlier than the middle Kounamkites Zone of the Siberian Amgan. Until now, the situation appears to be largely under control, but it becomes more complicated if the correlation includes the occurrences of Arthricocephalus chauveaui and other species (such as Peronopsis roddyi or Kiskinella cristata) as mentioned above under 3.3. Whether we will accept a specific solution open-minded or insist on traditional concepts will affect the practicability of the boundary level which is the sole criterium to meet. History of stratigraphic research teaches us that alternative approaches – despite being well-settled and having advantages against others – will not develop a victorious concept simply by ordinary scientific processes. However, a Working Group consisting of scientists with different opinions and background will be able to channel considerations and to direct studies to a feasible solution. ACKNOWLEDGMENTS. Many thanks are given to John Peel (Uppsala) who generously provided specimens and figures of the trilobite material from North Greenland for this study, to Zhao Yuanlong (Guiyang) for photographs of relevant trilobite specimens from the Kaili Formation of Guizhou, to Fred Sundberg (Show Low, AZ) for photographs of trilobite specimens from Nevada and to Michael Streng (Uppsala) for technical support. In addition, Fred Sundberg provided most detailed information on the trilobites and stratigraphy of the Great Basin and corrected mistakes in the first version of the manuscript. Peter Jell (Brisbane) and Peng Shanchi (Nanjing) reviewed the submitted manuscript and very much enhanced the quality of the article providing critical remarks, which is greatly acknowledged. Thanks are also due to Duck Keun Choi (Seoul) who encouraged me to write this report. I also thank several colleagues for encouraging this study.. 95. REFERENCES Álvaro, J.J., Gozalo, R., Liñán, E. and Sdzuy, K., 1993, The palaeogeography of northern Iberia at the Lower-Middle Cambrian transition. Bulletin de la Société géologique, France, (VIII), 164, 843–850. Álvaro, J. and Liñán, E., 1997. Nuevos datos acerca del Bilbiliense (Cámbrico inferior terminal) en las Cadenas Ibéricas y su correlación con otras áreas. Revista Española de Paleontología, 12, 277–280. Astashkin, V.A., Pegel, T.V., Shabanov, Yu.Ya., Sukhov, S.S., Sundukov, V.M., Repina, L.N., Rozanov, A.Yu. and Zhuravlev, A.Yu., 1991, The Cambrian System on the Siberian Platform; correlation chart and explanatory notes. International Union of Geological Sciences, Publication, 27, 133 p. Banerjee, D.M., Schidlowski, M., Siebert, F. and Brasier, M.D., 1997, Geochemical changes across the Proterozoic-Cambrian transition in the Durmala phosphorite mine section, Mussoori Hills, Garhwal Himalaya, India. Palaeogeography, Palaeoclimatology, Palaeoecology, 132, 183–194. Bergeron, J.N., 1899, Étude de quelques trilobites du Chine. Bulletin de la Société Géologique du France, Series 3, 27, 181–207. Blaker, M.R. and Peel, J.S., 1997. Lower Cambrian trilobites from North Greenland. Meddelelser om Grønland, Geoscience, 35, 145 p. Brasier, M.D., 1985, Evolutionary and geological events across Precambrian-Cambrian boundary. Geology today, Sept–Oct, 141–146. Brasier, M.D., 1991, Nutrient flux and the evolutionary explosion across the Precambrian—Cambrian boundary interval. Historical Biology, 5, 85–93. Brasier, M.D., 1992, Paleoceanography and changes in the biological cycling of phosphorus across the Precambrian-Cambrian boundary. In: Lipps, J.H. and Signor, P.W. (eds.), Origin and Early Evolution of the Metazoa. Plenum Press, New York, p. 483–523. Brasier, M.D., 1994, Towards a carbon isotope stratigraphy of the Cambrian System: potential of the Great Basin succession. In: Hailwood, E.A. and Kidd, R.B. (eds.), High Resolution Stratigraphy. Geological Society Special Publication, 70, 341–350. Brasier, M.D. and Sukhov, S.S., 1998, The falling amplitude of carbon isotopic oscillations through the Lower to Middle Cambrian; northern Siberia data. Canadian Journal of Earth Sciences, 35, 353–373. Brasier, M.D., Anderson, M.M. and Corfield, R.M., 1992, Oxygen and carbon isotope stratigraphy of early Cambrian carbonates in southeastern Newfoundland and England. Geological Magazine, 129, 265–279. Brasier, M.D., Khomentovsky, V.V. and Corfield, R.M., 1993, Stable isotopic calibration of the earliest skeletal fossil assemblages in eastern Siberia (Precambrian-Cambrian boundary). Terra Nova, 5, 225–232. Brasier, M.D., Corfield, R.M., Derry, L.A., Rozanov, A.Yu. and Zhuravlev, A.Yu., 1994a, Multiple δ13C excursions spanning the Cambrian explosion to the Botomian [sic] crisis in Siberia. Geology, 22, 455–458. Brasier, M.D., Rozanov, A.Yu., Zhuravlev, A.Yu., Corfield, R.M. and Derry, L.A., 1994b, A carbon isotope reference scale for the Lower Cambrian succession in Siberia: report of IGCP Project 303. Geological Magazine, 131, 767–783. Brasier, M.D., Magaritz, M., Corfield, R., Luo, H., Wu, X., Ouyang, L., Jiang, Z., Hamdi, B., He, T. and Fraser, A.G., 1990, The carbon- and oxygen-isotope record of the Precambrian-Cambrian boundary interval in China and Iran and their correlation. Geo-.
(16) 96. Gerd Geyer. logical Magazine, 127, 319–332. Brøgger, W.C., 1878, Om Paradoxidesskifrene ved Krekling (About Paradoxides shales at Krekling). Nyt Magazin Naturvidenskap, 24, 18–88. Brøgger, W.C., 1882, Paradoxides Ölandicus-nivået ved Ringsaker i Norge. Geologiska Föreningens i Stockholm Förhandlingar, N:o 74, VI, 143–148. Brøgger, W.C., 1886, Om alderen af Olenelluszonen i Nordamerika. Geologiska Föreningens i Stockholm Förhandlingar, 101, VIII, 182–213. Chernysheva, N.E., 1962, Kembriyskie trilobity semeystva Oryctocephalidae (Cambrian trilobites of the family Oryctocephalidae). In: Shvedov, N.A. (ed.), Problemy neftegazonosnosti Sovetskoj Arktiki. Paleontologiya i biostratigrafiya, 3. Trudy nauchno-issledovatel'skiy institut geologii Arktiki (NIIGA), Gostoptekhizdat, Leningrad, 127, 3–52. Chernysheva, N.E., ed., 1971, Amginskiy yarus Altae-Sayanskoy oblasti. Trudy Sibirskoy nauchno-issledovatel’skiy institut geologii, geofiziki i mineral’nogo syr’ya (SNIIGGiMS), Serya paleontologiya i stratigrafiya, III, Zapadno-Sibirskoe nizhnoe izdat., Novosibirsk, 267 p. Datsenko, V.A., Zhuravleva, I.T., Lazarenko, N.P., Popov, Yu.N. and Chernysheva, N.E., 1968, Biostratigrafiya i fauna kembriyskikh otlozheniy severo-zapada Sibirskoy platformy (r. Sukharikha). Trudy nauchno-issledovatel’skiy institut geologii Arktiki (NIIGA), Nedra, Leningrad, 155, 212 p. Debrenne, F. and Peel, J.S., 1986, Archaeocyatha from the Lower Cambrian of Peary Land, central North Greenland. Rapport Grønlands geologiske Undersøgelse, 132, 39–50. Demokidov, K.K. and Lazarenko, N.P., 1959, Novye dannye po stratigrafiya kembriyskikh otlozheniy zapadnogo sklona severnogo Kharaulakha. In: Shvedov, N.A. (ed), Sbornik statey po paleontolologii i stratigrafii, 16, 11–12. Demokidov, K.K., Kaban’kov, V.Ya., Lazarenko, N.P., Savitskiy, V.E. and Sobolevskaya, P.F., 1959, Stratigrafiya siniyskikh i kembriyskikh otlozheniy severo-vostoka Sibirskoy platformy. Trudy nauchno-issledovatel’skiy institut geologii Arktiki (NIIGA), Gostoptekhizdat, Leningrad, 101, 210 p. Derry, L.A., Brasier, M.D., Corfield, R.M., Rozanov, A.Yu. and Zhuravlev, A.Yu., 1994, Sr and C isotopes in Lower Cambrian carbonates from the Siberian craton: A paleoenvironmental record during the ‘Cambrian explosion’. Earth and Planetary Science Letters, 128, 671–681. Fletcher, T.P., 1972, Geology and Lower to Middle Cambrian trilobite faunas of southwest Avalon, Newfoundland. Part Two. Unpubl. Ph. D. thesis, Univsity of Cambridge: 295 p. Fletcher, T.P., 1999, Significant trilobites in the basal beds of a Cambrian stage corresponding to the Amga Stage of Siberia. In: Palmer, A.R. (comp. and ed.), Laurentia 99. 5th Field Conference of the Cambrian Stage Subdivisian Working Group, International Subcommission on Cambrian Stratigraphy (Abstracts), Institute for Cambrian Studies, Boulder, CO, p. 55–56. Fletcher, T.P. 2001, Arthricocephalus chauveaui Bergeron – a key species for the correlation of a global stage boundary. In: Peng, S., Babcock L.E. and Zhu, M. (eds.), Cambrian System of South China. Palaeoworld, 13, 257–260. Fletcher, T.P. 2003, Ovatoryctocara granulata: the key to a global Cambrian stage boundary and the correlation of the olenellid, redlichiid and paradoxidid realms. Special Papers in Palaeontology, 70, 73–102. Geyer, G., 1983, Biostratigraphische und systematisch-paläontolo-. gische Untersuchungen im Grenzbereich Unter-/Mittelkambrium von Marokko. Unpubl. Ph. D. dissertation., Insitut für Paläontologie, Bayerische Julius-Maximilians-Universität Würzburg, 229 p. Geyer, G., 1988, Agnostida aus dem höheren Unterkambrium und dem Mittelkambrium von Marokko. Teil 2: Eodiscina. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 177, 93–133. Geyer, G., 1990a, Revised Lower to lower Middle Cambrian biostratigraphy of Morocco. Newsletters on Stratigraphy, 22, 53–70. Geyer, G., 1990b, Correlation along the Lower/Middle Cambrian boundary – a puzzling story with an elusory end? In: Repina, L.N. and Zhuravlev, A.Yu. (eds.), Tretiy mezhdunarodnyy simpozium po kembriyskoy sisteme. 1–9 avgusta 1990 g., SSSR, g. Novosibirsk. Tezisy dokladov, Novosibirsk, p. 100–102. Geyer, G., 1993, The giant Cambrian trilobites of Morocco. Beringeria, 8, 71–107. Geyer, G., 1998, Intercontinental, trilobite-based correlation of the Moroccan early Middle Cambrian. Canadian Journal of Earth Sciences, 35, 374–401. Geyer, G., 1999, The Lower-Middle Cambrian faunal turnover – reality or sampling artifact? In: Chen, J.-Y., Chien, P.K., Bottjer, D.J., Li G.-X. and Gao, F. (eds.), International Symposium “The Origins of Animal Body Plans and Their Fossil Records”. Early Life Research Center, Kunming, p. 20–21. Geyer, G. and Landing, E., 1995, The Cambrian of the Moroccan Atlas region. Beringeria Special Issue, 2, 7–46. Geyer, G. and Mergl, M., 1995, Mediterranean representatives of the obolellid Trematobolus (Brachiopoda) and a review of the genus. Paläontologische Zeitschrift, 69, 179–209. Geyer, G. and Palmer, A.R., 1995, Neltneriidae and Holmiidae (Trilobita) from Morocco and the problem of Early Cambrian intercontinental correlation. Journal of Paleontology, 69, 459–474. Geyer, G. and Landing, E., 2000, The Cambrian of Israel and Jordan – the feather edge of the Mediterranean Realm. In: Aceñolaza, G.F. and Peralta, S. (eds.), Cambrian from the southern edge. Instituto Superior de Correlacion Geológica (INSUGEO), Tucumán, Miscelánea, 6, p. 98–101. Geyer, G. and Landing, E., 2002, The Bani Stage and its significance for Cambrian intercontinental correlation. In: Álvaro, J.J. and Clausen, S. (eds.), VIII Conference of the Cambrian Stage Subdivision Working Group (Programme and Abstracts), CaunesMinervois, September 12–14, Lille, p. 17. Geyer, G. and Landing, E., 2004, A unified Lower-Middle Cambrian chronostratigraphy for West Gondwana. Acta Geologica Polonica, 54, 179–218. Geyer, G. and Shergold, J.H., 2000, The quest for internationally recognized divisions of Cambrian time. Episodes, 23, 188–195. Geyer, G., Landing, E. and Heldmaier, W., 1995, Faunas and depositional environments of the Cambrian of the Moroccan Atlas region. Beringeria Special Issue, 2, 47–119. Gil Cid, M.D., 1988, Nuevos datos sobre la fauna del Cámbrico Inferior (Marianense de Llerena (Badajoz). Boletín de Geología i Minero, 99-IV, 578–583 (108–113). Gil Cid, M.D., 1991, Caracterización paleontológica del Cámbrico de la zona de Ossa Morena (Z. O. M.) y sus paleorrelaciones con Marruecos y Centroeuropa. Boletín de Geología i Minero, 102, 3–18. Gozalo, R., 1995, El Cámbrico de las Cadenas Ibéricas. In: GámezVintaned, J.A. and Liñán, E. (eds.), Memorias de las IV Jornadas Aragonesas de Paleontología: “La expansión de la vida en el Cámbrico”. Libro homenaje al Prof. Klaus Sdzuy, Institución “Fernando el Católico”, Zaragoza, p. 137–167..
수치


관련 문서
