for Marine Environmental Engineering Vol. 15, No. 2. pp. 133-141, May 2012
133
남해연안해역에서 분리한 유독와편모조류 Alexandrium tamarense와 Alexandrium catenella의 개체군 변화에 관한 생리
·생태학적 연구
I. 수온과 염분의 변화에 따른 성장 특성
오석진1·박지아1·권형규1·양한섭1,†·임월애2
1부경대학교 해양학과
2국립수산과학원 수산해양종합정보과
Ecophysiological Studies on the Population Dynamics of Two Toxic Dinoflagellates Alexandrium tamarense and Alexandrium catenella
Isolated from the Southern Coast of Korea
I. Effects of Temperature and Salinity on the Growth
Oh Seok Jin1, Park Ji A1, Kwon Hyeong Kyu1, Yang Han Soeb1,† and Lim Weol Ae2
1Department of Oceanography, Pukyong National University, Daeyeon-dong, Nam-gu, Busan 608-737, Korea
2Fishery and Ocean Information Division, NFRDI, Busan 619-705, Korea
요 약
한국 남해연안해역에서 분리한 유독와편모조류 Alexandrium tamarense와 Alexandrium catenella의 성장에 영향을 미치는 수온과 염분의 특성을 실내 배양실험을 통해 살펴보았다. 수온은 10, 15, 20, 25, 30oC의 5단계, 염분은 10, 15, 20, 25, 30, 35 psu의 6단계를 조합한 총 30단계의 조건에서 성장속도 변화를 관찰하였다. 최대성장속도를 보 이는 수온과 염분은 A. tamarense에서 15oC와 30 psu, A. catenella에서 25oC와 30 psu로 나타났다. 또한 최적 성장조건은 A. tamarense에서 10-20oC, 25-35 psu이고, A. catenella에서 20-30oC, 25-35 psu였다. 얻어진 수온 과 염분의 조건을 이용하여 중회귀분석에 의해 예측 모델식을 계산한 결과, A. tamarense는 µ=0.04+0.0193T- 0.0339S- 0.0005T2+0.0021S2+0.00073TS-0.000022T3-0.000038S3+0.00000086TS2-0.0000255T2S이며, A. catenella 는 µ=1.01-0.1288T-0.0778S+0.0067T2+0.0038S2+0.00204TS-0.0001T3-0.000059S3-0.0000131TS2-0.0000392T2S로 나타났으며, 예측값과 실험값 사이에는 높은 상관관계를 보였다. 본 연구결과는 수온과 염분의 현장관측 값을 이 용하여 성장속도를 추정할 수 있기 때문에, A. tamarense와 A. catenella의 개체군 확대를 이해하는데 도움이 될 것으로 기대된다.
Abstract − Effects of temperature and salinity on the growth of the toxic dinoflagellate Alexandrium tamarense and A. catenella isolated from the southern coast of Korea were examined in the laboratory. Growth experiment was conducted under the following combinations of temperature and salinity; 10, 15, 20, 25 and 30oC, 10, 15, 20, 25, 30 and 35 psu at a constant irradiance of 300 µmol photons m-2 s-1. Temperature and salinity conditions for maximum growth rate were indicated as follows: temperature 15oC and salinity 30 psu (0.31 d-1) in A. tam- arense, temperature 25oC and salinity 30 psu (0.36 d-1) in A. catenella. Temperature and salinity ranges for opti- mum growth condition of two species were 10~20oC, 25~35 psu and 20~30oC, 25~35 psu, respectively. The result of two-factor ANOVA indicated significant effects (P<0.001) of temperature and salinity on the growth rate, and two species were more effected by a temperature than a salinity on the growth. In addition, prediction equations were obtained through the multiple regressions of the specific growth rates as µ=0.04+0.0193T- 0.0339S-0.0005T2+0.0021S2+0.00073TS-0.000022T3-0.000038S3+0.00000086TS2-0.0000255T2S in A. tamarense
†Corresponding author: [email protected]
and µ=1.01-0.1288T-0.0778S+0.0067T2+0.0038S2+0.00204TS-0.0001T3-0.000059S3-0.0000131TS2-0.0000392T2S in A. catenella. Correlation coefficient between experimental values and simulated values was highly indicated.
These results seem to provide information for understanding the spreading mechanism of A. tamarense and A.
catenella.
Keywords: Toxic dinoflagellate(유독와편모조류), Alexandrium tamarense, Alexandrium catenella, Maximum specific growth rate(최대성장속도), Optimum growth condition(최적성장조건)
1. 서 론
마비성 패독(paralytic shellfish poisoning, PSP)의 문제는 1970 년대에 온대해역으로 국한되었던 것이, 1990년대 이후부터 점차 광 역화 되었으며(Hallegraeff[1993]), 대부분의 연안역에서 양식되는 이매패류에 발생하게 되었다(Shumway et al.[1994]). 이 패독은 삭 시톡신(saxitoxin)이라 불리는 신경성 독(neurotoxins)의 일종이며, 연령 및 신체조건에 따라 다르지만, 약 14 mg(5,000~20,000 MU) 정도를 섭취하게 되면, 사망에 이를 수 있다(Genenah and Shimizu [1981]). 우리나라에서도 PSP에 의한 사망사고가 84년, 86년과 96 년에 있었으며(Chang et al.[1987]; Lee et al.[1997]), 그 이후 이매 패류에 의한 독화현상을 사전에 방지하는 차원에서 매년 4~5월에 굴 과 홍합과 같은 이매패류의 채취 및 출하를 자제시켰고, 출하 직전 독성을 검사하여 생물의 생체내 독성이 기준치(80 µg/100 g)이상으 로 검출되면 출하를 금지시켰다. Han et al.[1992]와 Lim[2006]에 의하면, 최근 우리나라 남해연안 일대에 PSP 원인생물의 개체수가 증가하고 있으며, 발생해역은 점차 광역화되고 있는 양상을 보고 하였다.
우리나라에서 대표적인 PSP 원인종은 Alexandrium tamarense (Lebor) Balech, A. catenella(Whedon et Kofoid) Balech 그리고 Gymnodinium catenatum(Graham) 이지만, G. catenatum에 의한 패류 독화는 현재까지 보고가 없으며, 대부분이 A. tamarense와 A.
catenella가 원인종으로 보고되었다. 지금까지 1997년 4월 진동만 에 한차례 A. tamarense 적조가 발생하였지만, 이 들 종은 우리나 라 서해와 남해에 광범위하게 분포 하고 있는 것으로 확인되었다 (Kim et al.[2002]; Kim and Kim[2004]). 이들 종은 적조에 준하는 밀도로 성장하는 예는 상당히 드문 현상이다. 하지만 이 종은 낮은 개체군 밀도에서도 이매패류를 독화시킬 수 있어 세심한 주의가 요 구된다(Sako et al.[1995]).
Alexandrium 속의 분류는 세포갑판(thecal plate)의 세부적인 관 찰에 따르지만(Fukuyo[1985]), A. tamarense와 A. catenella는 형 태학적으로 Alexandrium 속의 다른 종보다 유사하여 분류하기 매우 어렵다(Taylor et al.[1995]; Kim et al.[2002]). 하지만, A. tamarense는 PSP 독성군 중 gonyautoxin 5+6(GTX 5+6)의 부재로 A. catenella 와 독 조성에 차이가 있으며, 고성능 액체크로마토그래프 분석에 따라 어느 정도 분류가 가능하다(Sako et al.[1995]). 최근에는 분 자생물학적 측면에서 두 종의 분류가 시도 되고 있다(Adachi and Sako[1996]; Kim and Kim[2004]).
A. tamarense와 A. catenella의 개체군 변동을 파악하기 위한 두 종의 생리학적 특성에 관한 연구는 박 등[2004]이 여러 가지 환경 조건 하에서 A. tamarense 휴먼포자 발아율을 측정한 것과, 이 등 [2005], 이 등[2006] 그리고 Lee et al.[2006]이 실내배양실험에서 A. tamarense 또는 A. catenella의 수온, 염분, 빛 등의 환경요인에 대한 성장 특성을 파악한 것이 전부이다. 더욱이 우리나라 PSP에 따른 수산업 피해에 반하여, 두 종의 생리적인 특성을 서로 비교하 여 출현·예측 등을 고찰한 사례는 전무한 실정이다.
패독발생의 방지 및 예찰을 위해서라도 우리나라 해역에서 분리한 A. tamarense와 A. catenella의 여러 가지 환경변수에 대한 생리적 인 특성을 파악하는 것은 매우 중요하다. 따라서 본 연구는 그 첫 번째 단계로 수온과 염분의 변화에 따른 A. tamarense와 A. catenella 의 성장의 변화를 서로 살펴보고, 우점 전략을 고찰하였다. 또한 실 내실험을 통하여 도출된 자료를 바탕으로 발생 시기를 예측할 수 있는 모델식도 제안하여 향후 생태계 모델에 응용될 수 있도록 하 였다.
2. 재료 및 방법 마산만에서 Alexandrium 속의 월별 변화
2011년 3월부터 12월까지 마산만 한 정점(Stn. 2)에서 월별로 반 돈 채수기를 사용하여 표층 해수 1 L를 채수하고, Lugol 용액으로 최종농도 2%가 되게 고정한 후, 실험실로 옮겨 침전법에 의하여 농축하였다(Fig. 1). 시료를 균일하게 혼합 후 1 ml를 Sedwick- Rafter counting chamber에 첨가하여 검경하였으며, 세포수가 많은 경우 25% 또는 50% 희석하여 광학도립현미경(TE2000, Nikon, Japan)에서 계수하였다. 수온과 염분은 동 정점의 표층, 중층, 저층(B- 1 m; 퇴적물로부터 상부 1 m 수층)에서 채수한 해수(반돈 채수기 이 용)를 수온염분계(YSI 30, YSI, USA)로 측정하였다.
Alexandrium tamarense와 Alexandrium catenella의 분리 A. tamarense는 2011년 3월에 진해만(Stn. 1), A. catenella는 2011년 6월 마산만(Stn. 2)에서 표층해수를 채수하여(Fig. 1), pasteur pipette(Ø 50~100µm)으로 분리하였다(Guillard[1995]). 분리된 세 포는 여과해수(0.22 µm pore size, Millipore GSWP, USA)를 이용 하여 3~4회 반복 세척하여 분리주를 확립하였다. 사용된 배지는 동해 에서 채수한 외양수(N36°24'938'', E130°42'055'')를 바탕으로 한 개량 형 f/2 배지(Guillard and Ryther[1962])로, 배양튜브(DU.23175215,
SCHOTT/DURAN, Germany)에 10 ml 첨가 후, A. tamarense와 A.
catenella를 이식하였다. 전 배양시 온도, 염분 그리고 광조건은 채 수한 당시의 현장조건과 유사하게 두 종 모두 20oC, 30 psu 그리 고 300 µmol photons m-2 s-1(12L: 12D; coolwhite fluorescent lamp; FHF32SS, Kumho, Korea)로 하여 계대 배양하였다. A. tamarense 와 A. catenella에 대한 무균화 처리는 하지 않았지만, 2차 생물학적 오염을 막기 위해서 모든 실험기구는 고압멸균(202 kpa, 20 분) 및 건조멸균(120oC, 3 hr)시켜 사용하였다.
형광염색법을 통한 Alexandrium tamarense 와 Alexandrium catenella 배양주 확인
분리한 Alexandrium속이 A. tamarense 또는 A. catenella 배양주 인지 확인하기 위해, 대수성장기 세포와 배양액을 15 ml 튜브(FA 2096, BD Bioscience, USA)에 옮겨 담은 후, 5% glutaraldehyde를 최종농도가 0.5%가 되도록 첨가하여 고정하였다. 그 후 상등액을 제거하고, 99% 메탄올을 첨가하여 세포를 재현탁 시켜 24시간 동 안 냉장 보관하였다. Calcofluor white M2R 용액은 최종농도가 1 mg ml-1되도록 염색한 후, 형광현미경으로 검경하여 Taylor et al.
[1995]에 따른 갑판배열법으로 2종을 명확하게 동정하였다.
수온과 염분 단계별 성장속도 변화
각각의 세포를 현장수온과 염분에 상응하는 조건(20oC, 30 psu) 으로 광량은 약 300 µmol photons m-2 s-1에서 대수성장기가 될 때 까지 배양한 후, 여러 가지 세포밀도로 조제하여 형광광도계(Model 10AU005, Turner Designs, USA)로 in vivo chlorophyll 형광값을 측정하였다. 세포밀도와 형광값 사이에는 Fig. 2와 같은 관계식이 성립되었다. 수온과 염분에 따른 A. tamarense와 A. catenella의 성 장속도는 현미경을 이용한 직접 검경이 아닌 in vivo chlorophyll 형광값을 이용하여 Fig. 2에 제시한 관계식으로 간접적으로 산출 하였다.
수온과 염분에 따른 성장속도 실험은 수온 10, 15, 20, 25, 30oC 의 5단계, 염분 10, 15, 20, 25, 30, 35 psu의 6단계를 조합하여 총 30단계의 조건하에서 성장속도를 관찰하였다. 실험에 사용된 해수 는 동해 외양수로 염분농도가 33 psu이기 때문에 10~30 psu의 염 분은 초순수를 사용하여 희석하였으며, 35 psu는 자연 증발시켜 조 절하였다. 수온 20oC, 염분 30 psu 그리고 300 µmol photons m-2 s-1에 전배양(preculture)한 대수성장기 세포를 25 psu와 35 psu의 배지에 이식하여 배양하였다. 그리고 25 psu 배지의 세포가 대수 성장기 후기에 있을 때 20 psu로 이식하여 염분변화에 따른 영향 을 최대한 줄였다. 수온은 하루에 1oC씩 증감시켜 각각의 온도단 계에 도달하도록 하였다. 각각의 단계에 적응된 세포를 배양튜브 (DH. TT10100, DAIHAN, Korea)에 f/2 배지를 5 ml 주입한 후 세 포수가 약 100 cells ml-1가 되도록 첨가하였다. 그 후 이틀간격으 로 같은 시간에 in vivo chlorophyll 형광값을 측정하였다. 세포의 성장속도(growth rate; µ)는 대수성장(exponential growth)을 보이 는 기간 동안의 형광값을 이용하여 다음 식 (1)에 의해 계산하였다.
(1)
N0,Nt: 대수성장기에서 초기와 t시간 후의 세포수(cells ml-1)
∆t: 대수성장기의 기간(d-1) µ 1∆t--- Nt
N0
--- ln
= Fig. 1. Sampling station of the vegetative cell of Alexandrium tamarense
and Alexandrium catenella in the Masan and Jinhae Bay, Korea.
Fig. 2. Relationship between cell density and in vivo chlorophyll fluorescence of Alexandrium tamarense and Alexandrium catenella.
또한 A. tamarense와 A. catenella의 성장에 영향을 미치는 수온 과 염분의 관계를 알아보기 위해 식 (2)와 같은 다항식으로 표현하 여 성장예측모델을 작성하였으며, 3차항까지 변수증감법에 따른 중 회귀분석을 실시하였다.
µ=β00+β10·T+β20·T2+β30·T3+β01·S+β02·S2+β03·S3+β11·T·S
+β12·T·S2β21·T2·S+ ...βnn·Tn·Sn+ε (2) µ: 성장속도 (growth rate; d-1)
T: 수온(temperature; oC) S: 염분(salinity; psu)
βnn: 회귀계수(regression coefficient) ε: 오차항(error term)
3. 결과 및 고찰 마산만에서 Alexandrium 속의 월별 변화
조사기간 동안 마산만의 수온은 6.3~28.3oC의 범위로, 12월에 가장 낮은 수온을, 하계로 갈수록 수온이 증가하여 7월에 가장 높 은 수온을 보였으며, 6월부터 8월까지 성층이 형성되는 전형적인 온대해역의 수온분포 양상을 보였다. 염분은 13.5~33.6 psu의 범위 로, 4월에 가장 높았으며, 강우의 영향이 가장 크게 미쳤던 7월에 가장 낮은 염분과 강한 염분 약층이 형성되었다(Fig. 3A). 식물플 랑크톤 현존량은 7월과 8월에 Pseudonitzschia spp.와 Skeletonema costatum이 우점 하였으며, 특히 8월은 8×103 cells ml-1 이상으로 두 규조류의 혼합 적조가 발생하였다. 대체적으로 추계에 규조류와 와편모조류 모두 낮은 세포 농도를 보였다(Fig. 3B). Alexandrium spp.는 4월부터 7월까지 출현하였으며, 5월과 6월에 8 cells ml-1와 35 cells ml-1로 와편모조류의 약 60% 이상을 차지하는 우점종이었다.
형광염색법을 통한 Alexandrium tamarense와 A. catenella 배양주 확인
서론에서 언급한 것과 같이 A. tamarense와 A. catenella은 출현 시기 뿐만 아니라 형태학적으로 매우 유사하여, 두 종의 분류에 있 어 많은 어려움이 있다(Balech[1995]; Taylor et al.[1995]). 이들 종 의 동정은 세포갑판(thecal plate)의 세부적인 관찰을 필요로 하며, 특히 Talyor et al.[1995]는 Alexandrium속을 분류하는데 3가지 key diagnostic plates(apical pore complex; a.p.c., first apical plate; f.a.p., posterior sulcal plate; p.s.p.)의 중요성을 강조했다. A. tamarense와 A. catenella는 a.p.c.와 p.s.p.의 유사하지만, f.a.p.에 ventral pore(v.p.) 가 A. tamarense의 경우 확인가능하며, A. catenella는 v.p.가 확인 되지 않는다(Talyor et. al.[1995]). 하지만 A. tamarense의 v.p.의 경 우 계대배양이 오래 진행될수록, 배양주에 따라 v.p.가 사라지는 변 형이 발생하는 경우가 있기 때문에, 세포벽 판을 통한 동정을 위해 서는 분리 직후 배양된 배양주에 대해 확인 작업을 수행하여야 할 것이다.
본 연구에서 실내배양실험을 위해 분리한 Alexandrium 배양주를
calcofluor white M2R을 통한 형광염색법으로 세포갑판을 살펴본 결과, 상각에 존재하는 a.p.c.와 하각의 중앙에 존재하는 p.s.p.는 두 분리주 모두 유사하였다(Fig. 4). 하지만 3월에 분리된 세포에는 f.a.p.에 v.p.가 존재하여 A. tamarense로 판단하였으며(Fig. 4B; E), 6월에 분리된 세포에는 v.p.가 존재하지 않아 A. catenella로 판단 하였다(Fig. 4H; J).
수온과 염분 단계별 성장속도 변화
본 연구에서 수행한 수온과 염분 조건 중, A. tamarense는 수온 30oC를 제외한 모든 염분 조건에서 성장하였다. 성장하지 않은 조 Fig. 3. Monthly variation of temperature (A), salinity (B) and com- position of phytoplankton (C) in the Masan Bay, Korea.
건을 제외하고 염분조건에 따른 수온별 성장속도를 살펴보면, 10 psu에서 0.03~0.12 d-1, 15 psu에서 0.06~0.19 d-1, 20 psu에서 0.12~0.24 d-1, 25 psu에서 0.21~0.25 d-1, 30 psu에서 0.19~0.31 d-1, 35 psu에서 0.08~0.22 d-1의 범위였다(Figs. 5 and 7). A. catenella는 A.
tamarense와 다르게 수온 10~15oC 중 10~20 psu 염분에서 성장하 지 않았으며, 30oC의 모든 염분조건에서 성장하였다. 성장하지 않은 조건을 제외하고 염분조건에 따른 수온별 성장속도를 살펴보면, 10 psu에서 0.08~0.12 d-1, 15 psu에서 0.12~0.24 d-1, 20 psu에서 0.19~
0.26 d-1, 25 psu에서 0.17~0.29 d-1, 30 psu에서 0.19~0.36 d-1, 35 psu에서 0.09~0.28 d-1로 나타났다(Figs. 6 and 7).
A. tamarense와 A. catenella의 수온과 염분에 대한 성장속도를 Fig. 4. Thecal plates structures of Alexandrium tamarense and Alex- andrium catenella by calcofluor white M2R staining. A, G: optical photomicrograph of vegetative cells, B: ventral view, C: dorsal view, D: epitheca, E: apical pore complex (a.p.c.) and first apical plate with ventral pore (v.p.), F: posterior sulcal plate (p.s.p.) with posterior attachment pore (p.a.p.), H: ventral view, I: dorsal view, J: epitheca, K: apical pore complex (a.p.c.) and first apical plate, L: posterior sulcal plate (p.s.p.) with posterior attachment pore (p.a.p.).
Fig. 5. Growth curves of Alexandrium tamarense grown at various water temperature and salinity combinations. Each symbol repre- sents the average of triplicate data.
Fig. 6. Growth curves of Alexandrium catenella grown at various water temperature and salinity combinations. Each symbol repre- sents the average of triplicate data.
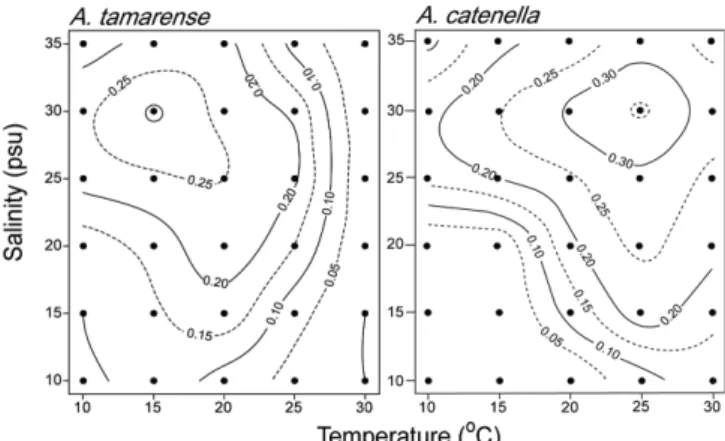
contour plotting으로 표현한 결과, A. tamarense는 수온이 낮고, 염 분이 높을수록 성장속도가 증가하여, 수온 15oC, 염분 30 psu에서 0.31 d-1로 최대 성장속도가 나타났으며(Fig. 7) 점차 감소하였다.
A. catenella는 수온과 염분이 높을수록 높은 성장속도를 보였으며, A. tamarense보다 10oC 높은 수온인 25oC와 염분 30 psu에서 0.36 d-1로 최대 성장속도가 나타나다가(Fig. 7) 점차 감소하였다. A.
catenella와 A. tamarense의 최적 성장조건(optimum growth condition, 최대성장속도의 80%이내 구간)을 알아보면, A. tamarense는 10- 20oC, 25-32 psu이고, A. catenella는 20-30oC, 25-35 psu였다.
한편, 본 연구에서 도출된 결과를 우리나라와 가까운 일본해역의 분리주와 비교하면, 일본 Mikawa 만에서 분리한 A. tamarense 배 양주는 최대성장속도를 보였던 수온과 염분이 15oC와 32 psu (Yamamoto et al.[1995]), 일본의 Hiroshima만 배양주의 경우는 수온 17oC, 염분 25 psu(Hamasaki et al.[2001]), 동 만에서 Yamamoto et al.[1997]는 15oC와 30 psu였다. 또한 우리나라 남해안 분리주의 경우 16~22oC, 30~35 psu(이 등[2005]) 그리고 진해만 분리주는 15oC, 30 psu(Lee et al.[2006])로 본 연구와 유사하였다. 하지만 Taiwan 분리주는 28oC(Su et al.[1993]), Canada의 동부연안 분리 주는 15-23 psu(Prakash[1967])로 분리해역에 따라 동일종이라도 생리적인 특징이 차이가 있다는 것을 의미한다(Gallagher[1982];
Lee and Shim[1995]). 적어도 우리나라와 일본 A. tamarense 분리 주의 경우 수온과 염분에 대해서는 유사한 생리학적 특성을 보이는 것으로 생각된다. 실제 1997년 진동만에서 일어났던 A. tamarense bloom의 경우, 수온이 12.3~12.7oC, 염분이 32.4~32.8 psu일때 발 생하였다(유와 Fukuyo[2000]). 경쟁종 배제와 다른 환경조건이 충 족한다면, 본 연구에서 밝혀진 최대성장을 보인 수온과 염분 조건 의 80%이내에 구간은 이 종이 적조를 일으킬 충분한 수온과 염분 범위임을 알 수 있다. 또한 매년 Alexandrium bloom이 일어나는 Hiroshima 만의 경우도 10.2~20.2oC 범위인 4월중에 발생하고 있 다(Itakura et al.[2002]).
A. catenella의 경우, A. tamarense에 비하여 연구사례가 적다. 홍 콩 Tai Tam 만 분리주의 최적성장조건은 수온 20~25oC, 염분
30~35 psu로 진해만 분리주와 유사하게 나타났다(Gavin et al.[1997]). 또한, 지중해의 분리주는 27oC와 30 psu 이상(Laabir et al.[2011]) 그리고 일본 Tokuyama만 분리주는 20~25oC, 25~35 psu였다. 하지만, 일본 Iwata현 인근해역의 분리주는 지금까지 보 고와 다르게 10~15oC로 A. tamarense의 최적성장을 위한 수온범 위와 유사하였으며, 염분은 32 psu로 본 실험과 유사하였다 (Iwasaki[1992]). 이와 같이 Iwasaki[1992]의 보고를 제외한다면, 다 른 지역의 분리주는 본 실험과 유사한 수온과 염분에 대한 생리적 인 특성을 가진 것으로 보인다.
마산만에서 관찰한 Alexandrium spp.의 경우(Fig. 3), 판(thecal plate)의 세부적인 관찰 및 분자생물학적 동정을 하지 않아 정확히 어떠한 종이 분포하는지 알 수는 없었다. 만약 현장에서 관찰한 Alexandrium spp.를 A. tamresne와 A. catenella로 한정시킨다면, 실내실험에서 도출한 결과를 이용하여 추론이 가능할 것으로 보인 다. 비교적 저수온기인 4월과 5월의 경우 A. tamarense가, 6월은 A.
tamarense와 A. catenella의 혼합, 7월은 A. catenella가 우점 할 가 능성이 높다고 판단된다. 특히 7월은 마산만 표층염분농도가 낮았 는데, 실내배양실험에서 고수온, 저염분에서는 Alexandrium 2종의 성장속도가 현저하게 떨어지는 양상(0.1 d-1)을 보인 것으로 보아, 실제 현장에서 이들 2종은 고수온기인 7월과 8월 이후 수주내(water column) 유영세포(vegetative cell)의 개체군이 완전히 소멸되어, 그 이후 출현하지 못하는 궁극적 원인이라 생각된다.
배양실험으로부터 얻어진 성장속도에 따라 수온과 염분조건을 이용한 이원배치분산분석(two-factor ANOVA)을 수행한 결과, 0.001% 유의수준 내에서 두 종의 성장속도에 대해 수온과 염분은 유의한 영향을 나타내었다. A. tamarense의 경우, 수온과 염분 및 그들의 상호작용의 제곱 합(sum of squares)은 총 제곱 합(total sum of squares)의 62.7%, 23.7% 및 11.9%를 차지하였으며(Table 1), A. catenella는 각각 53.2%, 39.8% 및 0.6%를 차지하여 두 종 모 두 성장은 수온의 영향을 더 많이 받는 것으로 나타났다(Table 2).
우리나라 주요 적조 종인 C. polykrikoides도 본 연구와 유사한 실 험을 수행한 결과 각각 30.5%, 65.8%, 96.3%을 보고 하여, 성장은 수온과 염분의 상호작용이 중요하지만, 그 중에서도 염분에 대하여 크게 영향을 받는 것으로 나타났다(Kim et al.[2004]; 오 등[2010]).
최근 서해안에서 적조를 일으키는 Chattonella marina의 경우 본 연구종과 유사하게 90.6%, 2.3%, 그리고 5.5%로 수온에 크게 의 존하여 성장하는 것을 알 수 있다(노 등[2006]). 이와 같이 비교적 Fig. 7. Contour plotting of growth rate in Alexandrium tamarense
and Alexandrium catenella.
Table 1. Summary of two-way analysis of variance of the population growth rate of Alexandrium tamarense as a function of temperature, salinity and their interaction
Source of variation Sum of squares d.f. Mean square F Temperature 0.511 4 0.128 2.041
Salinity 0.193 5 0.038 7.946
Interaction 0.097 20 0.005 1.543
Error 0.012 60 2.06E-04
Total 0.814 89
*p<0.001
외양에서 적조를 형성하는 종은 염분에, 연안에서 적조를 형성하는 종은 수온이 성장을 조절하는 주요 요인임을 알 수 있다. 추후에도 우리나라에서 발생하는 적조 원인종에 대하여 실내실험을 통하여 자료를 수집한다면, 최근 남해안의 일부 만에서 발생하는 댐의 수 문 개방에 따른 저염화 및 이상 기후에 따른 종천이 과정을 어느 정도 예측할 수 있을 것으로 생각된다.
중회귀 분석에 의한 A. tamarense와 A. catenella 출현 예측 모 델식은 다음과 같이 계산되었다.
Alexandrium tamarense: µ=0.04+0.0193T-0.0339S-0.0005T2 +0.0021S2+0.00073TS-0.000022T3-0.000038S3+0.00000086TS2
-0.0000255T2S(r2=0.869) (3)
Alexandrium catenellae: µ=1.01-0.1288T-0.0778S+0.0067T2 + 0.0038S2+0.00204TS-0.0001T3-0.000059S3-0.0000131TS2 -0.0000392T2S (r2=0.849) (4)
예측 모델식에 대해 실내실험에 사용된 수온과 염분 구간을 대 입하여 성장속도를 계산한 결과, 배양실험을 통해 얻은 실측값과 계산값 사이에 높은 상관관계가 나타났다(Fig. 8). 여러 가지 환경 변수가 식물플랑크톤의 성장에 관여하나, 수온과 염분 간의 두 함 수만 고려한다면 상기 (3)과 (4)의 모델식은 훌륭히 두 종의 성장
속도를 예측할 수 있을 것으로 생각된다.
한편, 적조생물의 이상발생과 적조 발생 해역의 광역화는 생활 사와 밀접한 관련이 있다(Anderson and Wall[1978]; Anderson et al.[1983]). 특히 유성생식을 통한 cyst는 유영세포의 성장에 적합 하지 않는 환경을 보낼 수 있기 때문에, 개체군 대발생의 씨앗종 (seed population)으로 중요한 역할을 한다(Anderson and Morel [1979]). Cyst의 발아에는 여러 가지 환경요인이 영향을 주지만, 그 중 수온이 가장 큰 영향을 준다(Anderson[1980]; Dale[1983]). Anderson [1998]에 따르면 A. tamarense cyst는 5~21oC에서, Park[1999]는 10~15oC에서 가장 높은 발아율을 보이며, 20~25oC에서는 발아가 관찰되지 않는다고 보고하였다. 이러한 수온은 A. tamarense 유영 세포의 최적성장조건과 유사하였다. 따라서 동계 또는 춘계에 cyst 발아 후에 최대성장속도에 도달할 수 있기 때문에, 타 종과 경쟁에 있어 유리한 위치를 차지할 것으로 보인다. A. catenella의 경우, 남 아프리카에서 sediment trap으로 채집한 cyst는 10oC일 때 최대 발 아율을 보여 유영세포의 최대성장속도를 보인 수온과 일치하지 않 았다(Joyce and Pitcher[2006]). 하지만 일본 Tanabe 만과 일본 Seto 내해의 퇴적물에서 분리한 cyst는 각각 20oC와 17.5oC에서 높은 발아율을 보였다(Takeuchi[1994]; Itakura and Yamagichi [2005]). A. tamarense와 같이 최대성장속도와 최대발아율을 보인 수온조건이 일치하지는 않지만, 본 실험의 최적성장속도를 보인 수 온범위로 춘계에 발아를 시작하여 하계에 우점할 수 있는 생리적 인 조건을 가진 것으로 판단된다.
2010년에 마산만, 진해만 및 진동만 일대에 Alexandrium속에 의 한 패류독소 발생으로 패류채취 금지 조취가 취해졌으나, 2011년 에는 Alexandrium spp. 출현이 확인되었음에도 불구하고 마산만에 서 패독이 발생하지 않았다. 이는 수온과 염분 이외에도 다른 환경 적 요인이 그들의 성장과 패독발생에 중요하게 기여하고 있는 것 을 의미한다. 추후 A. tamarense와 A. catenella의 성장에 영향을 주는 화학적 또는 생물학적 요인에 대한 생리특성에 대해 연구할 필요성이 대두되었다.
Table 2. Summary of two-way analysis of variance of the population growth rate of Alexandrium catenellae as a function of temperature, salinity and their interaction
Source of variation Sum of squares d.f. Mean square F Temperature 0.536 4 0.134 2.041
Salinity 0.401 5 0.080 1.946
Interaction 0.064 20 0.003 1.543
Error 0.006 60 1.03E-04
Total 1.008 89
*p<0.001
Fig. 8. Relationship between experimental data and simulated data of growth rate in Alexandrium tamarense and Alexandrium catenella.
사 사
본 논문은 부경대학교 환경·해양대학 간접비 2011년도 신진교수 연구력 강화 지원사업으로 수행된 연구로, 지원기관에 감사를 표합 니다.
참고문헌
[1]노일현, 윤양호, 김대일, 오석진, 2006, “가막만에서 분리한 유해성 침편모조류 Chattonella marina(Subrahamanyn) Hara et Chihara(Raphidophyceae)의 성장에 미치는 수온, 염분 및 빛의 영향”, 한국수산과학회지, Vol.39, 487-494.
[2]박명환, 김영옥, 조수연, 한명수, 2004, “마산만에서 분리한 Alexandrium tamarense 휴먼시트의 발아와 환경요인의 영향”, 한국환경생물학회, Vol.22, 200-205.
[3]오석진, 김창훈, 권형규, 양한섭, 2010, “2008년 한국 남해 안에서 분리한 유해 와편모조류 Cocholodinium polykrikoides Margelef의 성장에 미치는 수온, 염분 그리고 광조건의 영 향”, 한국수산과학회지, Vol.43, 715-722.
[4]유종수, Fukuyo, Y., 2000, “국내에서 처음 발생한 유독 플 랑크톤 Alexandrium tamarense 적조에 관하여”, 한국해양환 경공학회 2000년도 추계학술대회 논문집, 61-68.
[5]이창규, 이옥회, 이삼근, 2005, “한국연안에서 분리한 적조 형성 미세조류 10종의 성장에 미치는 온도, 염분, 광도의 영향”, 바다, Vol.10, 79-91.
[6]이해옥, 이나운, Toshiya, K., 한명수, 2006, “진해만의 유독 플랑크톤 Alexandrium catenella의 성장특성”, 한국환경생 물학회지, Vol.27, 147-154.
[7] Adachi, M., Sako, Y. and Ishida, Y., 1996, “Analyses of Alex- andrium (Dinophyceae) species using sequences of the 5.8S ribosomal DNA and internal transcribed spacer regions”, J. Phy- col., Vol.32, 424-432.
[8] Anderson, D.M. and Wall, D., 1978, “Potential importance of benthic cysts of Gonyaulax tamarensis and G. excavata in ini- tiating toxic dinoflagellate blooms”, J. Phycol., Vol.14, 224-234.
[9] Anderson, D.M. and Morel, F.M.M., 1979, “The seeding of two red tide blooms by the germination of benthic Gonyaulax tam- arensis hypnocysts”, Estuar. Coas. Mar. Sci., Vol.8, 279-293.
[10] Anderson, D.M., 1980, “Effects of temperature conditioning on development and germination of Gonyaulax tamarensis (Dino- phyceae) hypnozygotes”, J. Phycol., Vol.16, 166-172.
[11] Anderson, D.M., Chisholm, S.W. and Watra, C.J., 1983, “Impor- tance of life-cycle events in the population dynamics of Gon- yaulax tamarensis”, Mar. Biol., Vol.76, 179-189.
[12] Anderson, D.M., 1998, “Physiology and bloom dynamics of toxic Alexandrium species, with emphasis on life cycle transition”, In:
Anderson, D.M., Cembella, A.D. and Hallegraeff, G.M. (eds), Physiological ecology of harmful algal blooms. Springer Verlag, Berlin, 29-48.
[13] Balech, E., 1995, “The Genus Alexandrium Halim (Dinoflagel-
late)”, Sherkin Island Marine Station, Sherkin Island, 151pp.
[14] Chang, D.S., Shin, I.S., Pyeun, J.H. and Park, Y.H, 1987, “A study on paralytic shellfish poison of sea mussel, Mytilus edu- lis”, Bull. Korean. Fish. Soc., Vol.20, 293-299.
[15] Lee, C.K. and Lim, W.A., 2006, “Variation of Harmful Algal Blooms in Masan-Chinhae Bay”, ScienceAsia, Vol.32, 51-56.
[16] Dale, B., 1983, “Dinoflagellate resting cysts”, In: Fryxell, G.A., Cambridge Univ. Press, Cambridge, 69-136.
[17] Fukuyo, Y., 1985, “Morphology of Protogonyaulax tamarensis (Lebour) Taylor and Protogonyaulax catenella (Whedon and Kofoid) Taylor form Japanese Coastal Waters”, Bull. Mar. Sci., Vol.37, 529-537.
[18] Gallagher, J.C., 1982, “Physiological variation and electrophoretic banding patterns of genetically different seasonal populations of Skeletonema costatum (Bacillariophyceae)”, J. Phycol., Vol.18, 148-162.
[19] Gavin, K.Y.S., Maria, L.C.Y. and Chan, D.K.O., 1997, “Envi- ronmental and nutritional factors which regulate population dynam- ics and toxin production in the dinoflagellate Alexandrium catenella”, Hydrobiologia, Vol.352, 117-140.
[20] Genenah, A.A. and Shimizu, Y., 1981, “Specific toxicity of paralytic shellfish poisons”, J. Agric. Food. Chem., Vol.29, 1289-1291.
[21] Guillard, R.R.L. and Ryther, J.H., 1962, “Studies of marine planktonic diatoms Cyclotella nana Hustedt and Detonula con- jervaces (Cleve) Gran”, Can. J. Mircobiol., Vol.8, 223-239.
[22] Guillard, R.R.L., 1995, “Culture methods”, In: Hallegraeff, G.M., Anderson, D.M. and Cembella, A.D. (eds.), Manual on Harmful Marine Microalgae, UNESCO, New York, 45-62.
[23] Hallegraeff, G.M., 1993, “A review of harmful algal blooms and their apparent global increase”, Phycologia, Vol.32, 7999.
[24] Han, M.S., Jeon, J.K. and Kim, Y.O., 1992, “Occurrence of dinoflagellate Alexandrium tamarense, a causative organism of paralytic shellfish poisoning in Chinhae Bay”, J. Plankton Res., Vol.14, 1581-1592.
[25] Itakura, S., Yamaguchi, M., Yoshida, M. and Fukuyo, Y., 2002,
“The seasonal occurrence of Alexandrium tamarense (Dino- phyceae) vegetative cells in Hiroshima Bay”, Jpn. Fish. Sci., Vol.68, 77-86.
[26] Itakura, S. and Yamaguchi, M., 2005, “Morphological and phys- iological differences between the cysts of Alexandrium catenella and A. tamarense (Dinophyceae) in the Seto Inland Sea, Japan”, Plankton Biol. Ecol., Vol.52, 85-91.
[27] Iwasaki, J., 1992, “Effect of temperature, salinity and pH on the growth of Alexandrium catenella, a paralytic shellfish poison producing dinoflagellate”, Bull. FIsh. Exp. Stn. Ibaraki-ken, Vol.30, 93-100 (in Japanese).
[28] Joyce, L.B. and Pitcher, G.C., 2006, “Cysts of Alexandrium catenella on the west coast of South Africa: distribution and characteristics of germination”, Afri. J. Mar. Sci., Vol.28, 295-298.
[29] Kim, D.I., Mastuyama, Y., Nagasoe, S., Yamaguchi, M., Yoon,
Y.H., Oshima, Y., Imada, N. and Honjo, T., 2004, “Effects of temperature, salinity and irradiance on the growth of the harm- ful red tide dinoflagellate Cochlodinium polykrikoides Margalef (Dinophyceae)”, J. Plankton Res., Vol.26, 61-66.
[30] Kim, K.Y. and Kim, C.H., 2004, “A molecular phylogenetic study on Korean Alexandrium catenella and A. tamarense iso- lates (Dinophyceae) based on the partial LSU rDNA sequence data”, J. Kor. Soc. Oceanogr., Vol.39, 163-171.
[31] Kim, Y.O., Park, M.H. and Han, M.S., 2002, “Role of cyst ger- mination in the bloom initiation of Alexandrium tamarense (Dino- phyceae) in Masan Bay, Korea”, Aquat. Microb. Ecol., Vol.29, 279-286.
[32] Hamasaki, K., Horie, M., Tokimrrsu, S., Toda, T. and Taguchi, S., 2001, “Variability in toxicity the dinoflagellate Alexandrium tamarense isolated from Hiroshima Bay, western Japan, as a reflection of changing environmental conditions”, J. Plankton Res., Vol.23, 271-278.
[33] Laabir, M., Jauzein, C., Genovesi, B., Masseret, E., Grzebyk, D., Cecchi, P., Vaquer, A., Perrin, Y. and Collos, Y., 2011,
“Influence of temperature, salinity and irradiance on the growth and cell yield of the harmful red tide dinoflagellate Alexandrium catenella colonizing Mediterrancean waters”, J. Plankton Res., Vol.10, 1550-1563.
[34] Lee, H.O., Ishimaru, T., Toshiya, K. and Han, M.S., 2006,
“Growth of the dinoflagellate Alexandrium tamarense isolated from Jinhae Bay, Korean in axenic cultures”, Kor. J. Environ.
Biol., Vol.24, 275-281.
[35] Lee, J.S., Shin, I.S. and Kim, Y.M., 1997, “Paralytic shellfish toxins on the mussel, Mytilus edulis, caused the shellfish poi- soning accident at Geoje, Korea, in 1996”, J. Kor. Fish. Soc., Vol.11, 158-160.
[36] Lee, W.H. and Shim, J.H., 1995, “Intraspecific variation in the light intensity niche component of the diatom Skeletonema cos- tatum from Korean coastal water”, J. Kor. Sci. Oceangr., Vol.30, 436-441.
[37] Park, M.H., 1999, “Population dynamics of Alexandrium tam- arense cysts and vegetative cells in Masan Bay, Korea”, MSc the-
sis, Hanyang University, Seoul (in Korean with English abstract).
[38] Prakash, A., 1967, “Growth and toxicity of a marine dinoflagel- late, Gonyaulax tamarensis”, J. Fish. Res. Bd. Can., Vol.24, 1589- 1606.
[39] Sako, Y., 1995, “Life cycle and spread of toxic dinoflgaellate A.
tamarense”, Monthly Ocean, Vol. 27, 628-635 (in Japanese).
[40] Shumway, S.E., Cembella, A.D. and Richard, L., 1994, “Seques- tering and putative biotransformation of paralytic shellfish tox- ins by the sea scallop Placopecten magellanicus: seasonal and spatial scales in natural populations”, J. Mar. Bio. Ecol. Vol.1, 1- 22.
[41] Su, H.M., Chiang, Y.M. and Liao, I.C., 1993, Role of temper- ature, salinity and ammonia on the occurrence of the Taiwanese strain of Alexandrium tamarense. In: Smayda, T.J. and Shimizu, Y. (eds.), Toxic phytoplankton Biomass in the Sea. Elsevier, Amsterdam. 837-842.
[42] Takeuchi, T., 1994, “The ecology of Alexandrium catenella, a toxic red tide dinoflagellate in Tanabe Bay, Wakyama Prefec- ture”, Fish. Exp. Stn. Vol.2, 1-88(in Japanese with English abstract).
[43] Taylor, F.J.R., Fukuyo, Y. and Larsen, J., 1995, “Taxonomy of harmful dinoflagellates”, In: Hallegraeff, G.M., Anderson, A.D.
and Cembella, A.D. (eds.), Manual on harmful marine microal- gae, UNESCO, Paris, 283-317.
[44] Yamamoto, T., Yoshizu, Y. and Tarutani. K., 1995, “Effect of Temperature, Salinity and Irradiance on the Growth of Toxic Dinoflagellate Alexandrium tamarense isolated from Mikawa Bay, Japan”, Jpn. J. Phycol., Vol.43, 91-98.
[45] Yamamoto, T. and Tarutani, K., 1997, “Effects of temperature, salinity and irradiance on the growth of toxic dinoflagellate Alexandrium tamarense isolated from Hiroshima Bay, Japan”, Jpn. J. Phycol., Vol.45, 95-101 (in Japanese with English abstract).
2012년 3월 8일 원고접수 2012년 4월 7일 심사수정일자 2012년 4월 30일 게재확정일자