─ 385 ─ ISSN: 2288-1115(Print), 2288-1123(Online)
ⓒ The Korean Society of Limnology. All rights reserved.
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-Commercial License(http://creativecommons.org/licenses/by-nc/3.0/), which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provide the original work is properly cited.
INTRODUcTION
Chironomid, or non-biting midges, comprise more than 10,000 species worldwide (Ekrem and Willassen, 2004). Al- though several genera belonging to the family Chironomi- dae have a global distribution, others are endemic to tropical areas in Africa and South America (Ekrem and Willassen, 2004). The identification of chironomid species is mainly based on the morphological characteristics of larvae and
imagoes (Makarevich et al., 2000). Members of the family Chironomidae are frequently used by freshwater biologists to assess and monitor environmental conditions (Verneaux and Verneaux, 2002; Aagaard et al., 2004). Chironomid larvae are important indicator organisms for water pollution because of their ability to thrive under a wide variety of freshwater conditions (Wright, 1984; Armitage, 1995; Lin- dergaard, 1995; Aagaard et al., 2004), including exposure to heavy metals, organic pesticides, and other xenobiotics (Di Veroli et al., 2014; Matt et al., 2017). However, it is gener- ally difficult to distinguish between the larvae and females of closely related species based on morphology, and species identification frequently depends on the association of these
Classification and Distribution of Chironomidae (Diptera) using DNA Barcoding at Urban Streams in Gwangju, South Korea
Sang-Hoon Yoon *
(0000-0001-6294-2077), Jeong-Wook Park
(0000-0003-2906-9709), Ji-Young Park
(0000-0003-3390-0083)
, Jin-Jong Seo
(0000-0001-8834-8287), Suk-Kyung Jeong
(0000-0003-4159-7360), Jae-Keun chung
(0000-0002-2230-3438)and Seok-Jin Bae
(0000-0002-7522-7694)Health & Environment Research Institute of Gwangju, Gwangju 61986, Republic of Korea
Abstract Chironomid communities are indicators of water pollution because of their ability to thrive under
freshwater conditions. However, it is difficult to distinguish between chironomid larvae based on morphology.
DNA barcoding, based on nucleotide sequences of marker genes, can be used to identify chironomid larvae.
Samples of chironomid larvae were collected from Gwangju Stream and Pungyeongjeong Stream, tributaries of the Yeongsan River in South Korea. We identified 3 subfamilies, 13 genera, 16 species, and 1 cryptic species.
There were 7 genera and 10 species from the subfamily Chironominae, 5 genera and 5 species from subfamily Orthocladiinae, 1 genus and 1 species from subfamily Tanipodinae, and the cryptic chironomid species of the family Chironomidae. There were 21 individuals from, 7 species and 1 cryptic species from the Gwangju Stream and 24 individuals, belonging to 10 species from the Pungyeongjeong Stream. The only species detected in both streams was Cricotopus bicinctus. The relationship between water quality and the species detected was difficult to explain, but the number of species showed a tendency to increase at sites where water quality was poor.
Additional investigations and studies are needed to understand the relationship between water quality and the chironomid species occurring in these two streams.
Key words: benthic macroinvertebrate index, Chironomidae, Cricotopus bicinctus, DNA barcoding, ecosystem
health
Manuscript received 15 October 2019, revised 10 December 2019, revision accepted 11 December 2019
* Corresponding author: Tel: +82-62-613-7584, Fax: +82-62-613-7620, E-mail: [email protected]
Sang-Hoon Yoon, Jeong-Wook Park, Ji-Young Park, Jin-Jong Seo, Suk-Kyung Jeong, Jae-Keun Chung and Seok-Jin Bae 386
life stages with identified pupal exuviae of adult males, which tend to possess more species-specific characteristics (Ekrem et al., 2007).
DNA barcoding is a molecular method based on nucle- otide sequences of marker genes and is among the most informative techniques in phylogenetic analysis (Matsuhashi et al., 1999; Makarevich et al., 2000). In order to use DNA- based techniques, an appropriate molecular marker is critical (Kuncham et al., 2016). Mitochondrial DNA (mtDNA) is used in most molecular studies, and often only one or several genes can act as effective markers. The most commonly used gene is the cytochrome c oxidase I gene (COI). This gene has been widely used in evolutionary studies and, population ge- netics, as well as in species identification due to its relatively high degree of variation (Carew et al., 2003; Sharley et al., 2004). Use of COI has two advantages. First, the universal primers for this gene are robust, enabling recovery of its 5ʹ end from representatives of most, if not all, animal phyla (Folmer et al., 1994; Zhang and Hewitt, 1997). Second, COI appears to possess a greater range of phylogenetic signal than any other mitochondrial gene (Hebert et al., 2003).
River and stream ecosystem health assessment started as a Korean national project in 2016 (MOE, 2016). Algae, fish, benthic macroinvertebrates, and others are used in biotic water quality assessment. For the purposes of this ecosys- tem health assessment, the family chironomidae is divided into two types, based on the color of the larval body (Red or white).
Both Gwangju Stream and Pungyeongjeong Stream are approximately 20-km-long tributaries of the Yeongsan River. They are typical urban streams with anthropogenic disturbances such as rapid urbanization since the 1970̓s water pollution, purification projects, and public park proj- ects. The two streams have had insufficient maintenance flow and several kinds of water sources have been used to supplement the maintenance flow. There were three main- tenance flows discharged into the Gwangju Stream: water processed from a sewage treatment plant (STP), the lake water of the Juam Reservoir, and the water of the Yeongsan River. Meanwhile only the water from the Yeongsan River had been discharged into the Pungyeongjeong Stream.
Studies on the benthic macroinvertebrates, especially Chi- ronomidae, of small-scale urban streams such as Gwangju Stream and Pungyeongjeong Stream have been almost non- existent. The aims of this study were to identify species of chironomid larvae using the DNA barcoding method and to determine the characteristics of the distribution and abun-
dance of chironomid larvae along the two urban streams.
We also investigated the applicability of members of the family Chironomidae, as identified here, as a water quality indicator.
MATERIALS AND METHODS
1. Study area and sampling
Chironomid larvae were collected in 2016 from 10 sites in two urban streams: four sites (St.1-1 to St.1-4) in the Gwangju Stream and six sites (St.2-1 to St.2-6) in the Pungyeongjeong Stream; each site was sampled once. Sam- pling was performed at St.1-1 on January 12, at St.1-2 to St.1-4 on July 13~15, and at St.2-1 to St.2-6 on July 29.
Benthic macroinvertebrates were also collected and iden- tified along with the chironomid larvae. Stream ecosystem health was assessed using the Benthic Macroinvertebrate Index (BMI) of Korea, incorporating the species and individ- uals of benthic macroinvertebrates (MOE, 2016). Chemical water quality of each sampling site was analyzed. The mea- sured parameters included biological oxygen demand (BOD), chemical oxygen demand (COD), suspended solid (SS), total nitrogen (TN), and total phosphorus (TP) (MOE, 2015).
St.1-1 was a branch of the Gwangju Stream and was not affected by any maintenance flow additions. St.1-2 to St.1- 4 were affected by the STP discharge water and the water of the Yeongsan River. St.1-2 was also affected by the lake water of the Juam Reservoir during summer. St.2-1 to St.2- 6 were affected by the water of the Yeongsan River. St.2-1 was located in an agricultural area, St.2-2 and St.2-3 were located in industrial areas, and St.2-4 to St.2-6 were located in residential areas.
Chironomid larvae were collected using a Surber net (50 ×50 cm, mesh 1.0 mm). The larvae were washed through a 1-mm sieve, preserved in water for 5 days to excrete gut contents and fecal materials for preventing con- tamination, and stored in 80% ethanol (Pfenninger et al., 2007). The head capsule and the tail, with anal setae, were stored in 80% ethanol for future morphological examination (Yoon et al., 2011). The larval thorax and abdomen were used for DNA sequencing.
2. DNA extraction, PCR amplification
Genomic DNA was extracted larvae using a DNeasy
Blood and Tissue Kit (Qiagen, Germany) following the
manufacturer’s instructions. A 658 bp fragment of the mi- tochondrial COI gene was amplified using the universal primers LCO1490 (5ʹ-GGT CAA CAA ATC ATA AAG ATA TTG G-3ʹ) and HCO2198 (5ʹ-TAA ACT TCA GGG TGA CCA AAA AAT CA-3ʹ) (Folmer et al., 1994). The first PCR was performed with 2 μL of DNA extract and 18 μL distilled water treated with diethyl pyrocarbonate (DEPC; Gendpot, USA) in the first PCR premix tube (Intron, Korea). PCR conditions were as follows: initial denaturation at 94℃ for 5 min, 40 cycles of 94℃ for 30 sec, 58℃ for 30 sec, 72℃
for 40 sec, and final elongation at 72℃ for 5 min, using a Geneamp 9700 biosystem (ABI, USA). PCR products were confirmed by electrophoresis on a 1.5% agarose gel with ethidium bromide.
3. Sequence alignment and phylogenetic analysis
The amplified PCR products were sent to Cosmogenetech (Daejeon, Korea) for sequencing using the ABI 3730XL DNA Analyzer (Applied Biosystems, Foster City CA). The nucleotide sequence downloaded from the National Center
for Biotechnology Information database, and phylogenetic analyses were conducted via the MEGA6 program. Genetic distances were calculated using the Kimura-2-parameter (K2P) distance model (Kimura, 1980). Intraspecific and interspecific sequence divergences based on K2P distances were calculated for all species, and mean intraspecific and interspecific K2P divergences were calculated from pair- wise comparisons within each species. Neighbor-joining (NJ) and maximum likelihood (ML) analyses were per- formed using MEGA v6.0 with 1,000 bootstrap replicates.
RESULTS
In total, 46 individuals were sequenced from 40 speci- mens from the Gwangju Stream and 30 specimens from the Pungyeongjeong Stream (Table 1). We identified 3 subfam- ilies, 13 genera, 16 species, and 1 cryptic species. These were classified as 7 genera and 10 species of the subfamily Chironominae, 5 genera and 5 species of the subfamily Or- thocladiinae, 1 genus and 1 species of the subfamily Tanipo-
Fig. 1. Map of sampling sites. There were 4 sites(St.1-1 to St.1-4) in the Gwangju Stream and 6 sites(St.2-1 to St.2-6) in the Pungyeong-jeong Stream.
Sang-Hoon Yoon, Jeong-Wook Park, Ji-Young Park, Jin-Jong Seo, Suk-Kyung Jeong, Jae-Keun Chung and Seok-Jin Bae 388
Table 1. Summary of the dataset: collection location, number of sequences, divergences, and GenBank accession numbers. SubfamilyGenusSpeciesCollection locationNo. of sequencesDivergence(%) GenBank accession
number(COI)Identity (%)
Gwangju Stream
Pungyeongjeong StreamTotalSample IDIntraspecificInterspecific ChironominaeChironomusChironomus flaviplumus
St.1-2, St.1-4
13GJ-11, 12, 13, 14, 15, 20, 29, 30, 31, 3316.435.3JF41207599 Chironomus kiiensisSt.2-41PY-1631.4JF41208699 DicrotendipesDicrotendipes nervosus
St.2-1, St.2-2, St.2-5
5PY-1, 2, 8, 22, 2412.032.9JF41212897 HarnishiaHarnishia sp.St.2-31PY-1529.2KP90277898 MicrotendipesMicrotendipes pedellusSt.1-31GJ-2329.4KR74879992 PolydedilumPolydedilum nubiferSt.2-4, St.2-52PY-17, 233.832.8JF41214799 Polypedilum cultellatumSt.2-42PY-19, 204.530.9JF41215699 Polypedilum japonicumSt.2-11PY-328.9KU49707199 RheotanytarsusRheotanytarsus sp.St.2-61PY-3031.2KR65351289 TanytarsusTanytarsus formosanus
St.2-3, St.2-5, St.2-6
3PY-14, 25, 268.231.5KU72013399 OrthocladiinaeCricotopusCricotopus bicinctusSt.1-1
St.2-1, St.2-2, St.2-6
9
GJ-1, 3, 9 PY13.732.1KP90280499 -4, 7, 9, 10, 27, 29 OrthocladiusOrthocladius rivulorumSt.2-2, St.2-32PY-6, 132.434.4KR74911588 ParacricotopusParacricotopus sp.St.1-21GJ-1846.7HQ10523291 Paratrichocladius
Paratrichocladius rufiventris
St.1-21GJ-1686.8JN88709099 TveteniaTvetenia paucuncaSt.1-41GJ-3257.4KR29109388 TanipodinaeConchapelopiaConchapelopia sp.St.1-11GJ-231.4KR42733193 --Chironomidae sp.St.1-41GJ-3590.9KM92749789
dinae, and the cryptic species of the Chironomidae family.
There were 21 individuals from 7 species and 1 cryptic species from the Gwangju Stream and 24 individuals from 10 species from the Pungyeongjeong Stream. The only species detected in both streams was Cricotopus bicinctus.
The dominant and subdominant species were Chironomus flaviplumus (13 individuals) and Cricotopus bicinctus (3 in- dividuals) in the Gwangju Stream, and Cricotopus bicinctus (6 individuals) and Dicrotendipes nervosus (5 individuals) in the Pungyeongjeong Stream.
Interspecific divergences of sequenced samples were higher than intraspecific divergences. Mean intraspecific divergences of species with more than two individuals were 2.4~16.4%, and that of Cricotopus bicinctus was 13.7%.
Mean interspecific divergences were 28.9~90.9%. The species with the minimum intraspecific divergence was Or- thocladius rivulorum, and that with maximum interspecific divergence was Chironomus flaviplumus.
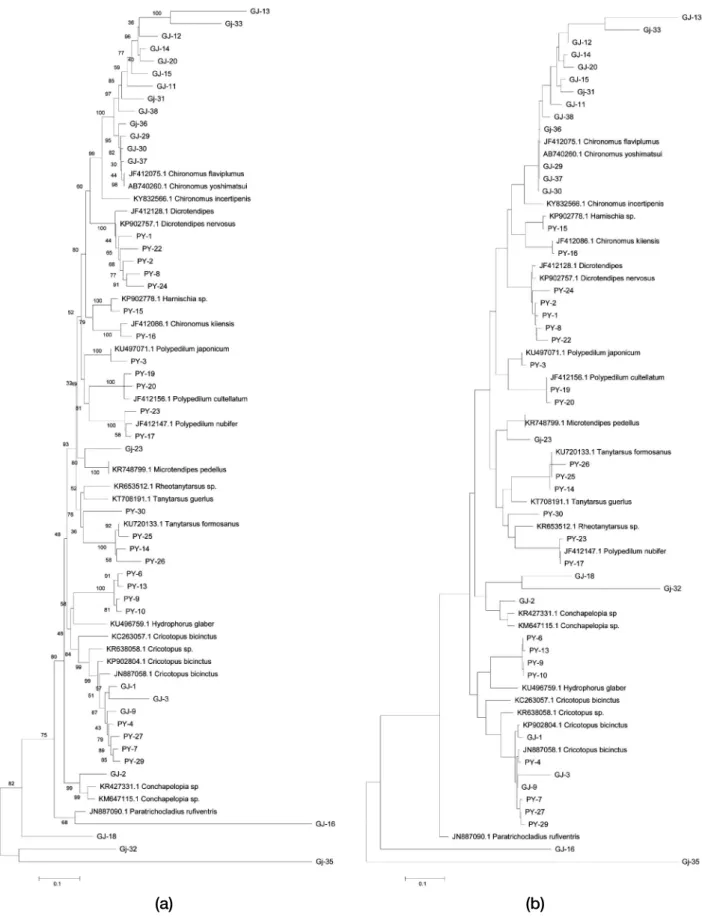
The result of the neighbor-joining (NJ) analysis calculated by genetic distances was similar to that of the maximum likelihood (ML) analysis calculated by base locations of genetic characters (Fig. 2). Both NJ and ML trees showed clear groupings at the species level. In the NJ tree, subfamily Chironominae, including Chironomus flaviplumus, Chi- ronomus kiiensis, Dicrotendipes nervosus, Harnishia sp., Microtendipes pedellus, Polydedilum nubifer, Polypedilum cultellatum, Polypedilum japonicum, Rheotanytarsus sp., and Tanytarsus formosanus produced monophyletic groups.
Subfamily Orthocladiinae including Cricotopus bicinctus, Paratrichocladius rufiventris and subfamily Tanipodinae, in- cluding Conchapelopia sp. were also monophyletic groups.
Some specimens were not shown in both trees because their homologies were below 90%. PY-6 and PY-13 was similar
to Orthocladius rivulorum (homology 88%), GJ-18 were Paracricotopus sp. (homology 88%), GJ-32 was similar to Tvetenia paucunca (homology 88%), and GJ-35 was similar to Chironomidae sp. (homology 89%) (Table 1, Fig. 1).
The environmental variables at the sampling sites in the Pungyeongjeong Stream (St.2-1 to St.2-6) did not show large differences, but those in the Gwangju Stream (St.1-1 to St.1-4) did show some differences (Table 2). The mean concentration of BOD was 2.1 mg L
-1in the Gwangju Stream and 1.9 mg L
-1in the Pungyeongjeong Stream.
Notably, the concentrations of BOD, TN, and TP were the highest at St.1-1 because of the inflow of domestic sewage.
The concentrations of SS and TP in the Pungyeongjeong Stream were higher than those in Gwangju Stream. The mean concentration of SS was 5.5 mg L
-1in the Gwangju Stream and 10.0 mg L
-1in the Pungyeongjeong Stream. The mean concentration of TP was 0.083 mg L
-1in the Gwangju Stream and 0.132 mg L
-1in the Pungyeongjeong Stream.
The biotic water quality, as assessed using the BMI, was class A (very good) at St.1-2, class B (good) at St.1-1 and St.1-3 in the Gwangju Stream, and class C (neutral) at most sites of the Pungyeongjeong Stream. In particular, the BMI of St.2-5 was class E (very bad) because the bed structure of St.2-5 was mostly composed of sand and the majority of the benthic macroinvertebrates collected were chironomid larvae.
DIScUSSION
Chironomid larvae are indicator organisms used in the assessment of water quality and toxicity in rivers and lakes (Kim et al., 2001; Kim et al., 2009). However, community
Table 2. Environmental variables and Benthic Macroinvertebrate Index(BMI) assessment.Site BOD(mg L-1) COD(mg L-1) SS(mg L-1) TN(mg L-1) TP(mg L-1) BMI
St.1-1 3.3 6.4 2.0 4.255 0.178 B
St.1-2 1.0 4.0 6.0 1.447 0.040 A
St.1-3 1.9 10.4 5.5 3.283 0.048 B
St.1-4 2.0 5.2 8.5 2.491 0.067 C
St.2-1 1.2 6.0 7.0 1.604 0.097 C
St.2-2 1.7 6.4 13.5 1.275 0.113 C
St.2-3 1.9 6.6 12.5 1.388 0.158 C
St.2-4 2.1 7.0 8.5 1.456 0.162 C
St.2-5 2.6 6.6 11.5 1.472 0.125 E
St.2-6 2.1 7.8 7.0 1.569 0.135 C
BOD: Biological Oxygen Demand; COD: Chemical Oxygen Demand; SS: Suspended Solid; TN: Total Nitrogen; TP: Total Phosphorus
Sang-Hoon Yoon, Jeong-Wook Park, Ji-Young Park, Jin-Jong Seo, Suk-Kyung Jeong, Jae-Keun Chung and Seok-Jin Bae 390
Fig. 2. (a) Neighbor-joining(NJ) tree and (b) Maximum likelihood(ML) tree of K2P distances based on mtDNA COI(1,000 bootstrap rep- licates). Bootstrap values are on branches; Scale=K2P distance.
(a) (b)
investigation of the family Chironomidae and water quality assessment using the species of this family poses difficulties because chironomid larvae cannot be easily distinguished based on their morphology. The method of DNA barcoding uses different sequences and not morphological informa- tion. It can be used to identify Chironomidae and investigate their community structure by using continuously accumu- lated sequence data (Kim et al., 2012; Hebert et al., 2016).
The BMI, using species and individuals, showed that the biological water quality of the Gwangju Stream was better than that of the Pungyeongjeong Stream (Table 2). The number of chironomid species was three at class A sites, three at class B sites, eleven at the class C sites, and three at the class E sites. Although the relationship between water quality and the species detected is difficult to explain be- cause of the small number of samples from each study site, the number of chironomid species showed a tendency to increase at the sites with a quality below class C. The water quality of class C was moderate, but sites below class C had poor water quality. The survey time for each site varied from January to July. This might also lead to differences in distribution of Chironomid due to water temperature and environmental differences at the time of investigation.
Cricotopus bicinctus was detected at many class B and C sites. The concentrations of BOD, COD, TN, and TP were the highest at class B St.1-1, indicating that Cricotopus bicinctus can inhabit a wider range of environmental con- ditions than can other chironomid species. Surber (2011) found that Cricotopus bicinctus was particularly resistant to electroplating wastes containing hexavalent chromium, cyanides, and copper, and to low oxygen levels.
Intraspecific divergences of Cricotopus bicinctus, detect- ed in both Gwangju Stream and Pungyeongjeong Stream, were 4.2~28.2% (Table 3). Intraspecific divergences were the lowest for samples PY-4 and PY-7 and the highest for GJ-3 and PY-10. Mean intraspecific divergence was 16.4%
for the three samples from the Gwangju Stream (GJ-1,3,9) and 16.9% for the four samples from the Pungyeongjeong Stream (PY-4,7,9,10). Mean intraspecific divergence among the three samples from Gwangju Stream and the four sam- ples from Pungyeongjeong Stream was 17.9%; no major difference was observed between the mean intraspecific di- vergences of the two streams. It was concluded that the nine specimens of Cricotopus bicinctus detected in Gwangju Stream and Pungyeongjeong Stream were the same species genetically.
cONcLUSIONS
Gwangju Stream and Pungyeongjeong Stream, the tribu- taries of the Yeongsan River, are urban streams with anthro- pogenic disturbances. The two streams have had insufficient maintenance flow and multiple kinds of water sources have been used to supplement maintenance flow. Although the two streams are just 3 km apart, the only chironomid species detected in both streams was Cricotopus bicinctus.
We could identify the species of chironomid larvae in our samples, but could not analyze the differences between the species collected from the two streams or the relation- ship between the genetic sequences and water qualities.
Large quantities of waste water processed at the STP are discharged into the Gwangju Stream, but in the Pungyeong- jeong Stream, only water from the Yeongsan River is dis- charged. Cricotopus bicinctus was detected at St.1-1, St.2- 1, St.2-2, and St.2-6, where there was no discharge of waste water processed at the STP. We believe that the waste water processed at the STP influences the presence of Cricotopus bicinctus. However, the number of species and individuals of benthic macroinvertebrates including Chironomidae, can change according to water quality, streambed structure, and other factors; thus, additional investigations and studies are required to elucidate this situation.
Author information Sang-Hoon Yoon (Health & Envi- ron ment Research Institute of Gwangju (HERI), Res- earcher), Jeong-Wook Park (HERI, Researcher), Ji-Young Park (HERI, Researcher), Jin-Jong Seo (HERI, Section chief), Suk-Kyung Jeong (HERI, Section chief), Jae-Keun Chung (HERI, Head of department), Seok-Jin Bae (HERI, Head of department)
Author contribution statement Conceptualization,
Table 3. Intraspecific divergences of Cricotopus bicinctus detectedin Gwangju Stream and Pungyeongjeong Stream.
GJ-1 GJ-3 GJ-9 PY-10 PY-4 PY-7 PY-9 GJ-1 0.0
GJ-3 13.4 0.0 GJ-9 12.9 16.3 0.0 PY-10 22.3 28.2 26.1 0.0
PY-4 10.2 13.7 5.6 23.0 0.0
PY-7 10.8 15.4 6.3 24.0 4.2 0.0
PY-9 22.0 28.2 25.5 2.0 23.7 24.6 0.0
Sang-Hoon Yoon, Jeong-Wook Park, Ji-Young Park, Jin-Jong Seo, Suk-Kyung Jeong, Jae-Keun Chung and Seok-Jin Bae 392
Sang-Hoon Yoon; Methodology, Sang-Hoon Yoon & Jeong- Wook Park & Jae-Keun Chung; Formal Analysis, Sang- Hoon Yoon & Jeong-Wook Park; Investigation, Sang-Hoon Yoon & Ji-Young Park; Resources, Jeong-Wook Park &
Ji-Young Park; Data Curation, Jeong-Wook Park & Jin-Jong Seo; Original Draft Preparation & Writing, Sang-Hoon Yoon; Review & Editing, Jin-Jong Seo & Suk-Kyung Jeong;
Supervision, Seok-Jin Bae; Funding Acquisition, Suk-Kyung Jeong & Seok-Jin Bae.
conflict of interest The authors declare no conflict of interest.
Funding This study was financially supported by the
‘Nation al Project for International Suitability Foundation Establishment of Experiment Analysis’ of the Ministry of Environment and the National Institute of Environmental Research and ‘Project for Strengthening Research Capa- bility’ of the Health and Environment Research Institute of Gwangju (Korea).
Acknowledgements We thank Editage (www.editage.
co.kr) for the manuscript editing and proofreading in En- glish.
REFERENcES
Aagaard, K., J.O. Solem, T. Bongard and O. Hanssen. 2004. Stud- ies of aquatic insects in the Atna River 1987-2002. Hydro- biologia 521: 87-105.
Armitage, P.D. 1995. Chironomidae as food, p. 425-428. In:
The Chironomidae: the biology and ecology of non-biting midge(Armitage, P., P.S. Cranston and L.C.V. Pinder, eds.).
Chapman and Hall, London, UK.
Carew, M.E., V. Pettigrove and A.A. Hoffmann. 2003. Identify- ing Chironomids(Diptera: Chironomidae) for biological monitoring with PCR-RFLP. Bulletin of Entomological Research 93: 483-490.
Di Veroli, A., F. Santoro, M. Pallottini, R. Selvaggi, F. Scardazza, D. Cappelletti and E. Goretti. 2014. Deformities of chiron- omid larvae and heavy metal pollution: From laboratory to field studies. Chemosphere 112: 9-17.
Ekrem, T. and E. Willassen. 2004. Exploring Tanytarsini relation- ships(Diptera: Chrionomidae) using mitochondrial COII gene sequences. Insect Systematics and Evolution 35: 263- Ekrem, T., E. Willassen and E. Stru. 2007. A comprehensive DNA 276.
sequence library is essential for identification with DNA barcodes. Molecular Phylogenetics and Evolution 43: 530- 542.
Folmer, O., M. Black, W. Hoeh, R. Lutz and R. Vrijenjoek. 1994.
DNA primers for amplication of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates.
Molecular Marine Biology and Biotechnology 3: 294-299.
Hebert, P.D.M., S. Ratnasingham, E.V. Zakharov, A.C. Telfer, V.
Levesque-Beaudin, M.A. Milton, S. Pedersen, P. Jannetta and J.R. deWaard. 2016. Counting animal species with DNA barcodes: Canadian insects. Philosophical Transac- tions of the Royal Society B 371(1702): 20150333.
Kim, J.Y., J.H. Lee and H.I. Ree. 2001. Seasonal population dy- namics of chironomids midges(Diptera: Chironomidae) emerging from reclaimed rice fields in Seosan, Korea in 1997-1999. Korean Journal of the Entomology 31(4): 225- Kim, B.S., Y.K. Park, S.S. Hong, Y.J. Yang, K.H. Park, M.H. 232.
Jeong, S.R. Kim, K.H. Park, W.H. Yeh, D.H. Kim, M.K.
Hong, Y.J. Ahn and J.S. Shin. 2009. Comparison of acute toxicity of molinate on two aquatic insects, Chironomus riparius and Cloeon dipterum in different larval stages. The Korean Journal of Pesticide Science 13(4): 256-261.
Kim, S.M., K.H. Song, H.I. Ree and W. Kim. 2012. A DNA bar- code library for Korean Chironomidae(Insecta: Diptera) and indexes for defining barcode gap. Molecules and Cells 33: 9-17.
Kimura, M. 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution 16:
111-120.
Kuncham, R., T. Thayumanavan and G.V.S. Reddy. 2016. Phy- logenetic relationship of some Indian chironomids based on mitochondrial DNA cytochrome oxidase I. Advances in Bioresearch 7(4): 46-51.
Lindergaard, C. 1995. Classification of water-bodies and pollu- tion, p. 385-404. In: The Chironomidae: the biology and ecology of non-biting midge(Armitage, P., P.S. Cranston, and L.C.V. Pinder, eds.). Chapman and Hall, London, UK.
Makarevich, I.F., E.V. Berezikov, V.P. Guryev and A.G. Blinov.
2000. Molecular phylogeny of the Chironomus genus de- duced from nucleotide sequences of two muclear genes, ssp160 and the globin 2b gene. Molecular Biology 34: 606- Matt, N.K., L.M. Bryant and S. Vink. 2017. Differential gene 612.
expression of Australian Cricotopus draysoni(Diptera:
Chironomiae) populations reveals seasonal association in detoxification gene regulation. Nature 7: 14263.
Matsuhashi, T., R. Matsuda, T. Mano and M.C. Yoshida. 1999.
Microevolution of the mitochondrial DNA control region in the Hapanese brown bear(Ursus arctos) population. Mo- lecular Biology and Evolution Society 16: 676-684.
MOE(Ministry of Environment). 2015. The standard method on water pollution, National Institute of Environmental Re- search. Incheon, Korea(in Korean).
MOE(Ministry of Environment). 2016. Biomonitoring survey
and assessment manual, National Institute of Environmen- tal Research. Incheon, Korea(in Korean).
Pfenninger, M., C. Nowak, C. Kley, D. Steinke and B. Streit.
2007. Utility of DNA taxonomy and barcoding for the in- ference of larval community structure in morphologically cryptic Chironomus(Diptera) species. Molecular Ecology 16: 1957-1968.
Sharley, D.J., V. Pettigrove and Y.M. Parson. 2004. Molecular Identification of Chironomus spp.(Diptera) for biomonitor- ing of aquatic ecosystems. Australian Journal of Entomol- ogy 43(4): 359-365.
Surber, E.W., 2011. Cricotopus bicinctus, a midgefly resistant to electroplating wastes. Transactions of the American Fish- eries Society 88: 111-116.
Verneaux, V. and J. Verneaux. 2002. Assessing lake functioning
using the macrobenthic community with special reference to Chironomidae(Diptera). A subalpine lake(Lake Annecy) as an example. Archiv fuer Hydrobiologie 154: 61-78.
Wright, J.F. 1984. The chironomid larvae of a small chalk stream in Berkshire, England. Ecological Entomology 9: 231-238.
Yoon, T.J., M.J. Baek, J.M. Hwang, H.J. Kang, W.Y. Choi, J.M.
Hur and Y.J. Bae. 2011. Analysis of molecular operational taxonomic unit(MOTU) in Chironomids(Chironomidae, Diptera) for use in indicator organisms for water quality assessment. Korean Journal of Nature Conservation 5(2):
77-82.
Zhang, D.X. and G.M. Hewitt. 1997. Assessment of the universal- ity and utility of a set of conserved mitochondrial primers in insects. Insect Molecular Biology 6: 143-150.