책임저자: 이삼준, 부산 남구 신선로 428번지
608-711, 동명대학교 보건복지대학 체육학과 Tel: 051-629-2106, E-mail: [email protected] 접수: 2012년 2월 27일, 심사: 2012년 2월 28일 게재승인: 2012년 3월 10일
이 논문은 2009년 정부(교육과학기술부)의 재원으로 한국연구재 단의 지원을 받아 수행된 연구임(NRF-2009-332-G00094).
노화 쥐의 성별에 따른 트레드밀 운동이 공간기억력에 미치는 영향
*경희대학교 의과대학 생리학교실, †동명대학교 보건복지교육대학 체육학과
신 말 순 *ㆍ김 보 균 *ㆍ이 수 신 *ㆍ허 유 미 *ㆍ김 창 주 *ㆍ이 삼 준
†
Effect of Treadmill Exercise According to Gender on Spatial Memory in Old-Aged Rats Mal-Soon Shin*, Bo-Kyun Kim*, Su-Shin Lee*, Yu-Mi Heo*, Chang-Ju Kim*, Sam-Jun Lee†
*Department of Physiology, College of Medicine, Kyung Hee University, Seoul, †Department of Physical Education, College of Health, Social Welfare and Education, Tong Myong University, Busan, Korea
We investigated the effects of treadmill exercise according to gender on spatial memory in old aged rats. Two month-old female rats and male rats were used as the young-aged groups, meanwhile 16-month-old female rats and male rats were used as the old-aged groups. The rats were divided into 8 groups (N=6 in each group): young-aged female control group, young-aged female exercise group, young-aged male control group, young-aged male exercise group, old-aged female control group, old-aged female exercise group, old-aged male control group, old-aged male exercise group. The rats in the exercise groups were forced to run on a motorized treadmill for 30 min once a day for 4 weeks. In old-aged rats, spatial memory was disturbed, in contrast treadmill exercise improved spatial memory in the old-aged rats. Neurogenesis in the hippocampus was decreased and apoptosis was increased in the old-aged rats. In contrast, treadmill exercise increased neurogensis and decreased apoptosis in the old-aged rats. Myostatin and follistatin expressions in the hippocampus were increased in the old-aged rats. Treadmill exercise suppressed myostatin and follistatin expressions in the old-aged rats. There was no gender difference in these effects of treadmill exercise. We showed that treadmill exercise is a very useful strategy for ameliorating age-induced memory loss by increasing neurogenesis through suppressing myostatin and follistatin expressions. (Korean J Str Res 2012;20:51∼60)
Key Words: Aging, Treadmill exercise, Spatial memory, Neurogenesis, Apoptosis
서 론
노화는 뇌에서 학습과 기억을 담당하는 해마에서의 기
능적인 변화를 초래하여, 학습능력 및 기억력 손실을 초래 한다(Jarrard, 1995). 노화에 따른 학습 및 기억력 손실은 신 경세포 생성과 밀접한 관련이 있는데, 신경세포 생성 (neurogenesis)은 사람을 포함한 포유동물의 특정 뇌 부위에 는 성장이 끝난 후에도 신경 간세포(neural stem cell)가 존재 하여 새로운 신경세포 생성이 지속적으로 일어난다(Eriksson et al., 1998). 특히, 새로운 신경세포의 생성이 이루어진다고 보고된 부위 중 해마(hippocampus)는 학습과 기억의 뇌 중 추로서, 해마 치상회(hippocampal dentate gyrus)에서 일어나 는 신경세포 생성은 학습, N-methyl-D-aspartate (NMDA) 수
용체 길항제, 세로토닌, 에스트로겐 및 운동 등으로 향상되 는 반면, 부신피질 호르몬, 스트레스와 노화로 인하여 억제 된다(Gould et al., 1999; Fuchs et al., 2000). 특히 노화는 해마 치상회에서 새로운 신경세포 생성을 감소시킬 뿐 아니라 세포사멸도 증가시킨다(Kuhn et al., 1996; Kim YP et al., 2004).
세포사멸(apoptosis)은 외부적 환경의 변화에 의해 급격히 일어나는 세포의 수동적 죽음인 괴사(necrosis)와는 달리 세 포 내부에 존재하는 죽음의 프로그램에 의해 여러 유전자 들이 발현됨으로써 일어나는 세포의 능동적인 죽음이며, 세포예정사(programmed cell death)라고도 한다(Wyllie, 1993).
세포사멸이 일어나는 세포의 형태적 특징으로는 세포체 수축(cell shrinkage), 세포막의 수포 생성(membrane blebbing), 세포사멸체(apoptotic body)의 형성, 핵에서의 염색사 응축 (chromatin condensation), DNA의 분절(DNA fragmentation) 등 이 보고되고 있다(Kerr et al., 1972). Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) 양성 세포는 DNA 가닥이 조각날 때 free 3'-OH 그룹을 감지하는 것으로 세포 사멸 동안 발생하는 DNA 분절 절단으로 TUNEL assay를 통 하여 관찰할 수 있다(Gavrieli et al., 1992). Caspases는 cystein 계열의 단백질 분해효소로서 세포사멸의 신호를 받으면 단백질내의 특정부위를 절단하는 효소이다. Caspase-3는 세 포사멸의 신호를 받아 세포사멸을 일으키는 실행자 역할 을 한다(Reed, 2000). 세포사멸의 신호에 의해 방출된 cyto- chrome c는 Apaf-1, caspase-9, dATP와 결합하여 caspase-9를 활성화시키고, 이렇게 활성화된 caspase-9은 caspase-3를 활 성화시켜 세포사멸을 유도하는 것으로 알려져 있다(Li et al., 1997). 이와 같은 세포사멸은 노화의 중요한 기전 중 하 나로 알려져 있다(Pollack et al., 2002; Mattson et al., 2006).
Myostatin은 transforming growth factor-β (TGF-β)군에 속 하며, 배아 및 성인 골격근에서 발현되어 근육세포 생성을 조절하는데 관여를 하는 것으로 알려져 있다(Siriett et al., 2006). 그리고 myostatin은 근육세포 생성과 관련되어 많이 알려져 있으나, 뇌의 전 지역에 분포되어 발현되는 것에 비하여 뇌세포 생성과의 관계에 대하여는 잘 알려져 있지 않다. 그러나 myostatin과 유사한 TGFβ1의 증가는 노화한 생쥐의 해마 신경세포 생성을 강력하게 억제시키는 것으 로 보고되었다(Buckwalter et al., 2006; Wachs et al., 2006).
Follistatin은 과립막 세포, 영양막 세포, 성선성장 세포들 을 포함한 다양한 세포들의 성장과 분화를 조절하며, 난포 의 생성, 배아의 성장, 임신에 관여하는 것으로 알려져 있
다(Bilezikjian et al., 1996). Follistatin은 생후 전 기간에 걸쳐 서 거의 모든 조직에서 발현이 되며, 또한 태아의 발달에 매우 중요한 역할을 하는 것으로 알려져 있다(Patel, 1998).
이전의 연구에서 follistatin의 과발현 유도는 근육의 증대를 가져오는 반면에, follistatin을 knockout시킨 생쥐에서는 근 육의 소실이 현저히 나타났다(Lee SJ et al., 2001). 그러나 follistatin의 뇌에서의 작용은 장기강화(long-term potentiation, LTP) 및 신경세포 생성을 억제시킨다고 보고되었다(And- reasson et al., 1995; Inokuchi et al., 1996).
운동은 노화를 비롯한 여러 가지 질환에 의한 근육의 위 축과 뇌의 신경세포 감소를 개선하고 기능적 회복을 촉진 하므로, 실제 임상에서도 고령자들의 근 위축의 억제 및 인지력을 회복시키기 위한 치료의 한 방법으로 많이 사용 되고 있다(Kambadur et al., 1997; Cotman et al., 2002). 많은 연구에서 운동은 노화된 뇌에서의 신경세포생성을 증가시 키고 노화에 의한 학습 능력의 감퇴를 방지한다고 알려져 있다(van Praag et al., 2005; Drapeau et al., 2007; Fabel et al., 2008).
노화에 의한 기억력의 감퇴에 대한 운동의 효과는 신경 세포 생성과의 관련성에 대해서는 많이 알려져 있지만, 이 러한 운동의 효과에 대하여 myostatin 및 follistatin의 작용과 의 관련성에 대해서는 보고되지 않았다. 나아가 남녀 성별 에 따른 운동의 효과를 규명한 연구는 아직 없다. 이에 본 연구에서는 젊은 흰쥐와 노화 흰쥐를 이용하여 성별에 따 른 트레드밀 운동이 공간기억력, 뇌 해마 치상회의 세포 사멸과 신경세포생성, 그리고 myostatin 및 follistatin의 발현 에 미치는 영향을 규명하였다.
재료 및 방법
1. 실험 동물
본 연구의 실험동물은 대한 오리엔트 바이오(Orient Co., Seoul, Korea)에서 분양 받은 2개월의 Sprague-Dawley 계열 암컷 흰쥐(N=12)와 수컷 흰쥐(N=12), 16개월의 노화된 Sprague-Dawley 계열 암컷 흰쥐(N=12)와 수컷 흰쥐(N=12) 를 사용하였다. 실험동물은 실험 전 일주일 동안 실험실 환경에 적응시킨 후 실험을 하였고, 실험기간을 통하여 고 형 사료와 물을 자유스럽게 섭취하도록 하였다. 온도는 22
∼24oC, 습도는 50±1%가 유지되도록 하였으며, 조명은 실 험동물의 야행성을 고려하여 밤과 낮(12시간 주/야)이 조절 되는 실험실 환경에서 사육하였다. 실험동물은 젊은 암컷
대조군(N=6), 젊은 암컷 운동군(N=6), 젊은 수컷 대조군 (N=6), 젊은 수컷 운동군(N=6), 노화 암컷 대조군(N=6), 노화 암컷 운동군(N=6), 노화 수컷 대조군(N=6), 노화 수 컷 운동군(N=6)의 8군으로 나누었다.
2. 트레드밀 달리기 프로토콜
운동은 소형동물용 트레드밀에서 하루 일회 30분간 4주 동안 실시하였다. 운동은 트레드밀 경사도 0%에서 초기 5 분간은 2 m/min 속도로 부하하다가 3 m/min 속도에서 25분 간 실시하였다.
3. BrdU의 주입
실험 시작 후 5일 동안, 일일 일회 50 mg/kg의 5-bromo- 2'-deoxyuridine (BrdU, Sigma Chemical Co., St. Louis, MO, USA)를 운동 시작 1시간 전에 모든 쥐에게 복강 주사하였 다.
4. Radial 8-arm maze 검사
Radial 8-arm maze 검사는 공간인지능력을 측정하기 위한 검사로(Ko IG et al., 2011; Yoon JS et al., 2011), 8개의 통로 (8×35 cm)가 있으며, 각 통로는 25 cm 높이의 투명한 벽이 둘러싸여 있다. 미로의 중심부는 지름 30 cm의 8각형 형태 를 이루고 있으며, 지면에서 1 m 높이에 위치해 있다. 각 arm의 끝부분에 지름 3 cm의 물그릇이 놓여진다. 검사에 사용될 쥐들은 조형시작 24시간 전부터 물 박탈에 들어가 며, 한 시기를 마친 후 1시간의 자유로운 물마시기를 허용 한 후 다시 24시간동안 물 박탈을 반복하였다. 실험용 쥐 는 미로의 중심부분에 놓이며, 정면으로 응시하는 방향이 무선(random)적이 되도록 하였다. 실험은 쥐가 물이 담긴 8 개의 모든 통로의 끝부분에 접근해 모든 물을 마시면 끝나 며, 모든 통로에 다다르지 못하더라도 10분이 경과하면 종 료되었다. 이 작업을 완수하기 위해서 쥐는 자신이 들어갔 던 통로와 들어가지 않은 통로를 기억하고 있어야 한다.
따라서 매 시행에서 자신이 들어간 통로와 관련된 정보를 보유하는 기억이 요구되는 과제이다. 행동측정 변인은 8개 의 모든 물통에 도착하는데 소요된 시간과 물을 마신 통로 에 다시 들어가는 오류회수를 측정하였다.
5. 뇌의 적출 및 조직처리
실험이 끝난 동물은 Zoletil 50Ⓡ (10 mg/kg, i.m.; Vibac, Carros, France)으로 마취시킨 후 흉강을 열고 우심방 귀를
절개함과 동시에 좌심실을 통하여 50 mM phosphate buffe- red saline(PBS)를 5분간 관류시켜 조직의 혈액성분을 제거 하고, 100 mM의 phosphate buffer (PB)에 녹인 4%의 parafor- maldehydye (PFA) 용액을 4oC에서 10분간 관류하였다. 안구 앞쪽의 전두골(frontal bone)을 절단한 후 두개골을 열고 뇌 를 적출하여 4% PFA에 담가 4oC에서 24시간 후 고정하였 다. 고정된 뇌 조직을 30% sucrose 용액에서 일주일간 침전 시켜 동결 방지한 후 freezing microtome (Leica, Nussloch, Germany)으로 냉동 절편을 40 μm의 두께로 연속관상 절 편을 제작하여 저온실에 보관하였다.
6. TUNEL 염색
신경 세포사멸을 나타내는 DNA 분절을 측정하기 위하 여 terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) 염색을 시행하였다(Ko IG et al., 2011). 염색방법은 In Situ Cell Death Detection KitⓇ (Roche, Mannhein, Germany) 를 사용하였다. 조직을 슬라이드에 부착하여 1일 동안 건 조 후 PBS 로 3회 세척한 후 ethanol-acetic acid (2:1) 용액으 로 −20oC에서 5분간 반응시켰다. 다음 PBS로 5분간 3회 세척하였고 0.1% sodium citrate에 0.5% Triton X-100이 함유 된 용액으로 얼음에서 2분 동안 반응시켰다. PBS로 5분간 3회 세척 후 3% H2O2 용액으로 실온에서 5분간 반응시키 고 PBS로 3회 세척한 후 조직을 슬라이드에 부착시켰다.
각 조직 위에 100 μg/ml proteinase K를 점적 후 1시간동안 37oC에서 반응시키고 PBS로 2분간 5회 세척 후 슬라이드 주변의 용액을 제거하고 20 μl의 효소 용액을 점적 후 습 도를 유지하면서 37oC에서 1시간 동안 반응시켰다. PBS로 2분간 5회 세척 후 주위 물기를 제거한 후 converter-POD를 각 조직당 30 μl 점적 후 37oC에서 습도를 유지하며 30분 간 반응시키고 PBS로 2분간 5회 세척하였다. 발색제로는 3,3'-diaminobenzidine tetrahydrochloride (DAB)을 50 mM Tris 완충액(pH 7.6)에 0.02% 농도로, H2O2는 0.03%로 사용하였 다. 발색반응은 상온에서 10분간 시켰으며, 반응이 끝난 후 조직을 PBS로 10분씩 3회 세척하였다. 발색이 끝난 조직은 gelatin-coated slide에 얹어서 2시간 동안 실온에서 건조시킨 후 에탄올의 농도를 높여가며 탈수시키고 xylene으로 투명 화시켜 polymount로 봉입한 후 광학 현미경으로 관찰하여 TUNEL-양성 세포수를 측정하였다.
7. Caspase-3, myostatin, follistatin 면역조직화학 법
Caspase-3, myostatin, follistatin 발현을 측정하기 위하여 면 역조직화학법을 시행하였다(Ko IG et al., 2011). 각 그룹에 서 해마 절편을 선택하여 먼저 조직 내에 존재하는 내재성 peroxidase를 비활성화시키기 위해서 50 mM PBS에 1%로 희 석된 H2O2에 30분간 반응시킨 후, 3회 50 mM PBS로 세척 한 후, 1% BSA와 10%의 horse serum으로 1시간 동안 반응 시킨 후 각각 anti-caspase-3 (1:500; Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-myostatin (1:500; Millipore Corpora- tion in Billerica, MA, USA), anti-follistatin (1:500; Santa Cruz Biotechnology) 항체를 0.05% BSA와 0.3% Triton X-100이 들 어있는 일차 항체용액에서 4oC에서 24시간 동안 실온에서 진탕하면서 반응시켰다. 다음 50 mM PBS로 세척한 후, biotinylated 2차 항체(Vector Laboratories, Burlingame, CA, USA)를 50 mM PBS에 0.3% Triton X-100을 함유한 이차 항 체용액에 1:200의 비율로 희석한 후 1시간 동안 반응시켰 다. 50 mM PBS로 세척한 후, avidin-biotin complex (Vector Laboratories)에서 1시간 동안 반응시켰다. 50 mM PBS로 세 척한 후, 50 mM Tris 완충액에 0.03% H2O2와 0.03% DAB을 함유하는 발색제로 발색반응을 실시한 다음 세척한 후 에 탄올의 농도를 높여가며 탈수시키고 xylene으로 투명화 시 켜 polymount로 봉입하였다.
8. BrdU 면역조직화학염색법
신경세포의 생성을 BrdU 면역조직화학법으로 검증하였 다(Yoon JS et al., 2011). 조직절편을 50 mM PBS로 세척한 후 0.5% Triton X-100에 반응시킨다. Triton X-100을 세척한 후 65oC에서 50% formamide와 2×SSC 용액에서 2시간 반응 시켰다. 반응 완료 후 2×SSC로 세척한 후 37oC에서 2N HCl 에 반응시켰으며, 반응이 끝나면 바로 100 mM sodium borate, pH 8.5로 세척한 후 50 mM PBS로 다시 세척하였다.
이 과정이 끝난 조직은 자유부유(free-floating)법을 사용하여 면역조직화학 염색을 시행하였다. 항체와 반응하기 전에 50 mM PBS에 10% horse serum과 1% bovine serum albumin (BSA)을 함유한 차단용액으로 한 시간 동안 반응시켰다.
BrdU 항체(Boehringer Mannheim, Germany)를 50 mM PBS에 0.5% BSA와 0.5% sodium azide를 함유한 일차 항체 용액에 1:600의 비율로 희석한 후 24시간 동안 반응시켰다. 50 mM PBS로 세척한 후, biotinylated 2차 항체(Vector Laboratories)를
50 mM PBS에 0.3% Triton X-100을 함유한 이차 항체용액에 1:200의 비율로 희석한 후 1시간 동안 반응시켰다. 50 mM PBS로 세척한 후, avidin-biotin complex(Vector Laboratories)에 서 1시간 동안 반응시켰다. 50 mM PBS로 세척한 후, 50 mM Tris 완충액에 40 mg/ml nickel, 0.03% H2O2와 0.03%
DAB를 함유하는 발색제로 발색반응을 실시한 다음 세척 하였다. 세척 후 50 mM PBS에 10% horse serum과 1% bovine serum albumin (BSA)를 함유한 차단용액으로 한 시간 동안 반응시켰다. NeuN 항체(1:500, Chemicon International, Teme- cula, CA, USA)를 50 mM PBS에 0.5% BSA와 0.5% sodium azide를 함유한 일차 항체 용액에 24시간 동안 반응시켰다.
50 mM PBS로 세척한 후, biotinylated anti-mouse IgG를 50 mM PBS에 0.3% Triton X-100을 함유한 이차 항체용액에 1:200의 비율로 희석한 후 1시간 동안 반응시켰다. 50 mM PBS로 세척한 후, avidin-biotin complex (Vector Laboratories)에 서 1시간 동안 반응시켰다. 50 mM PBS로 세척한 후, 50 mM Tris 완충액에 H2O2와 DAB를 함유하는 발색제로 발색 반응을 실시한 다음 세척하였다.
9. 양성반응 세포수 측정
각각의 항체들과 면역 반응이 일어난 세포들은 광학현 미경(BX-51, Olympus, Japan)을 이용하여 관찰하였다. BrdU- 양성 세포 수, TUNEL-양성 세포 수, caspase-3-양성 세포 수, myostatin-양성 세포 수, 그리고 follistatin-양성 세포 수를 치 상회의 과립세포층 mm2당으로 정량적인 분석을 하였다.
10. 자료처리
측정된 결과들은 SPSS 통계 program (PASW statistics version 18.0, IBM, Armonk, NY, USA)을 이용하여 각 항목에 대한 평균(mean) 및 표준오차(standard error of the mean;
SEM)를 산출하였다. 집단 간의 차이 검정은 일원분산분석 (one-way analysis of variance; one-way ANOVA)을 이용하였으 며, 구체적인 사후검정은 Duncan의 방법을 이용하였다. 그 리고 유의수준은 p<0.05로 설정하였다.
결 과
1. 공간기억력의 변화
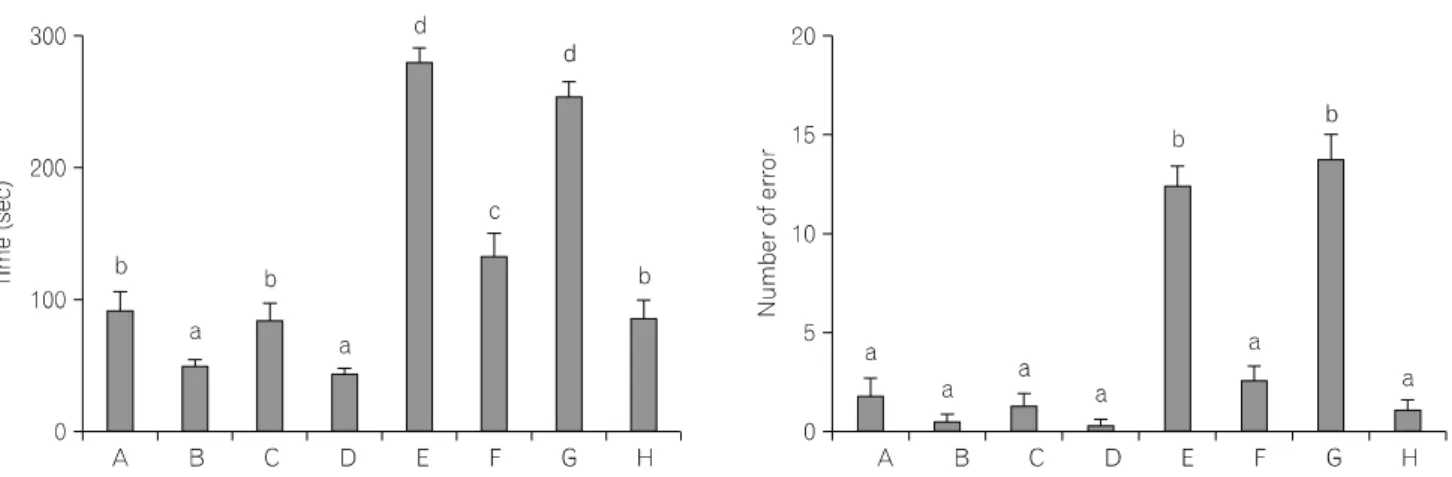
공간기억력을 측정하기 위한 검사 중 학습수행 시간과 오답 수에 대한 평균값은 Fig. 1에서 보는 바와 같다. 연구 결과, 젊은 쥐와 비교해 노화 쥐의 공간 기억력은 감소되
Fig. 1. Effects of treadmill exercise on spatial memory. Left: the time for accomplishing eight successful performances. Right: the number of errors made
before eight successful performances. The data are presented as the mean±SEM. (A) Young-aged female control group, (B) young-aged female and exercise group, (C) young-aged male control group, (D) young-aged male and exercise group, (E) old-aged female control group, (F) old-aged female and exercise group, (G) old-aged male control group, (H) old-aged male and exercise group. Different letters (a∼d) denote statistically significant differences (p<
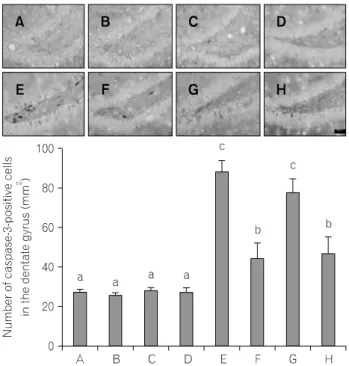
0.05) after Duncan post-hoc. For example, groups marked with different letters differ statistically.Fig. 2. Effect of treadmill exercise on caspase-3 expression in the
hippocampus. Above: photomicrographs of caspase-3-positive cells in the hippocampal dentate gyrus. The scale bar represents 50 μm. Below: (A) young-aged female group, (B) young-aged female and exercise group, (C) young-aged male group, (D) young-aged male and exercise group, (E) old-aged female group, (F) old-aged female and exercise group, (G) old-aged male group, (H) old-aged male and exercise group. The data are presented as the mean±SEM. Different letters (a∼c) denote statistically significant differences (p<
0.05) after Duncan post-hoc. For example, groups marked with different letters differ statistically.었으며, 트레드밀 달리기에 의하여 공간 기억력이 향상되 었다. 그러나 젊은 쥐와 노화 쥐 모두에서 암컷과 수컷 간 의 공간 기억력에는 유의한 차이는 나타나지 않았다. 공간 기억력에 대한 운동의 효과도 암컷과 수컷 사이에는 유의 한 차이가 없었다.
2. Caspase-3-양성 세포 수의 변화
해마 치상회 부위에서의 caspase-3-양성 세포 수의 평균값 은 Fig. 2에서 보는 바와 같다. 연구결과, 노화 쥐에서 caspase-3 양성 세포 수가 증가되었으며, 트레드밀 달리기에 의하여 억제되었다. 그러나 젊은 쥐와 노화 쥐 모두에서 암컷과 수컷 간의 caspase-3-양성 세포 수에는 유의한 차이 는 나타나지 않았다. Caspase-3-양성 세포 수에 대한 운동의 효과도 암컷과 수컷 사이에는 유의한 차이가 없었다.
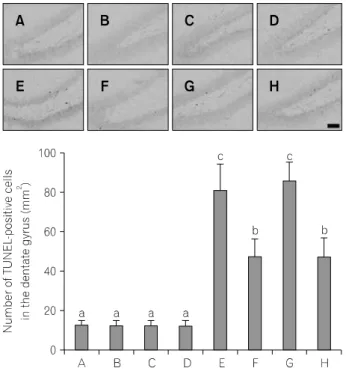
3. TUNEL-양성 세포 수의 변화
해마 치상회 부위에서의 TUNEL-양성 세포 수의 평균값 은 Fig. 3에서 보는 바와 같다. 연구결과, 노화 쥐에서 TUNEL-양성 세포 수가 증가되었으며, 트레드밀 달리기에 의하여 억제되었다. 그러나 젊은 쥐와 노화 쥐 모두에서 암컷과 수컷 간의 TUNEL-양성 세포 수에는 유의한 차이는 나타나지 않았다. TUNEL-양성 세포 수에 대한 운동의 효 과도 암컷과 수컷 사이에는 유의한 차이가 없었다.
4. BrdU-양성 세포 수의 변화
해마 치상회 부위에서의 BrdU-양성 세포 수의 평균값은
Fig. 3. Effect of treadmill exercise on DNA fragmentation in the hippo-
campus. Above: photomicrographs of terminal deoxynucleotidyl transfe- rase-mediated dUTP nick end labeling (TUNEL)-positive cells in the hippocampal dentate gyrus. The scale bar represents 50 μm. Below:number of TUNEL-positive cells in each group. The data are presented as the mean±SEM. (A) Young-aged female group, (B) young-aged female and exercise group, (C) young-aged male group, (D) young-aged male and exercise group, (E) old-aged female group, (F) old-aged female and exercise group, (G) old-aged male group, (H) old-aged male and exercise group. Different letters (a∼c) denote statistically significant differences (p
<
0.05) after Duncan post-hoc. For example, groups marked with different letters differ statistically.Fig. 4. Effect of treadmill exercise on neurpgenesis in the hippocampus.
in the hippocampus. Above: photomicrographs of BrdU-positive cells in the hippocampal dentate gyrus. The scale bar represents 50 μm. Below:
number of BrdU-positive cells in each group. The data are presented as the mean±SEM. (A) Young-aged female group, (B) young-aged female and exercise group, (C) young-aged male group, (D) young-aged male and exercise group, (E) old-aged female group, (F) old-aged female and exercise group, (G) old-aged male group, (H) old-aged male and exercise group. Different letters (a∼c) denote statistically significant differences (p
<
0.05) after Duncan post-hoc. For example, groups marked with different letters differ statistically.Fig. 4에서 보는 바와 같다. 연구결과, 노화 쥐에서 BrdU-양 성 세포 수가 감소되었으며, 트레드밀 달리기에 의하여 증 가하였다. 그러나 젊은 쥐와 노화 쥐 모두에서 암컷과 수 컷 간의 BrdU-양성 세포 수에는 유의한 차이는 나타나지 않았다. BrdU-양성 세포 수에 대한 운동의 효과도 암컷과 수컷 사이에는 유의한 차이가 없었다.
5. Myostatin-양성 세포 수의 변화
해마 치상회 부위에서의 myostatin-양성 세포 수의 평균 값은 Fig. 5에서 보는 바와 같다. 연구결과, 노화 쥐에서 myostatin-양성 세포 수가 증가되었으며, 트레드밀 달리기 에 의하여 감소하였다. 그러나 젊은 쥐와 노화 쥐 모두에 서 암컷과 수컷 간의 myostatin-양성 세포 수에는 유의한 차 이는 나타나지 않았다. Myostatin-양성 세포 수에 대한 운동 의 효과도 암컷과 수컷 사이에는 유의한 차이가 없었다.
6. Follistatin-양성 세포수의 변화
해마 치상회 부위에서의 follistatin-양성 세포 수의 평균 값은 Fig. 6에서 보는 바와 같다. 연구결과, 노화 쥐에서 follistatin-양성 세포 수가 증가되었으며, 트레드밀 달리기에 의하여 감소하였다. 그러나 젊은 쥐와 노화 쥐 모두에서 암컷과 수컷 간의 follistatin-양성 세포 수에는 유의한 차이 는 나타나지 않았다. Follistatin-양성 세포 수에 대한 운동의 효과도 암컷과 수컷 사이에는 유의한 차이가 없었다.
고 찰
운동은 뇌성마비, 뇌졸중 등의 뇌 손상이나 Alzheimer병, 파킨슨병과 같은 퇴행성 뇌 질환 환자에서 기능회복을 촉 진시키며 인지기능을 향상시킨다고 알려져 있다(Fordyce et
Fig. 5. Effect of treadmill exercise on myostatin expression in the
hippocampus. Above: photomicrographs of myostatin-positive cells in the hippocampal dentate gyrus. The scale bar represents 50 μm. Below:number of myostatin-positive cells in each group. The data are presented as the mean±SEM. (A) Young-aged female group, (B) young-aged female and exercise group, (C) young-aged male group, (D) young-aged male and exercise group, (E) old-aged female group, (F) old-aged female and exercise group, (G) old-aged male group, (H) old-aged male and exercise group. Different letters (a∼c) denote statistically significant differences (p
<
0.05) after Duncan post-hoc. For example, groups marked with different letters differ statistically.Fig. 6. Effect of treadmill exercise on follistatin expression in the
hippocampus. Above: photomicrographs of follistatin-positive cells in the hippocampal dentate gyrus. The scale bar represents 50 μm. Below:number of follistatin-positive cells in each group. The data are presented as the mean±SEM. (A) Young-aged female group, (B) young-aged female and exercise group, (C) young-aged male group, (D) young-aged male and exercise group, (E) old-aged female group, (F) old-aged female and exercise group, (G) old-aged male group, (H) old-aged male and exercise group. Different letters (a∼c) denote statistically significant differences (p
<
0.05) after Duncan post-hoc. For example, groups marked with different letters differ statistically.al., 1993; Johansson et al., 1996). 본 연구에서는 젊은 흰쥐와 노화 흰쥐의 공간기억력에 대한 운동의 효과를 성별에 따 라 비교하기 위하여 트레드밀 운동 4주 후에 8-arm maze test를 실시하였다. 실험 결과, 노화 쥐는 젊은 쥐와 비교해 공간학습 수행시간이 증가되었고 오류가 높았으나 트레드 밀 운동을 시행한 결과 노화 쥐의 성별에 관계없이 공간학 습 수행시간과 오류가 감소되었다. 그리고 젊은 쥐에서도 트레드밀 운동은 성별에 관계없이 공간학습 수행시간과 오류를 감소시켰다. 본 연구결과 노화에 의해 유발되는 공 간기억력 감퇴를 트레드밀 운동이 향상 시킬 수 있다는 것 을 의미하며, 젊은 쥐에서도 트레드밀 운동은 공간기억력 의 향상에 도움을 줄 수 있다는 것을 의미한다.
노화는 인간과 동물의 인지력 감소와 밀접한 관계가 있 다(Patrylo et al., 2007; Is et al., 2008). 사람을 대상으로 한 연 구에서, 노화는 해마의 위축을 유발하지만(West, 1993;
Small et al., 2002), 운동을 한 고령자들은 운동을 하지 않은 고령자들에 비해서 뇌조직의 손실이 적은 것으로 나타났 다(Colcombe et al., 2003). 운동에 의한 인지력 및 기억력의 향상은 신경세포생성의 증가, 시냄스 가소성의 증가, 신경 전도 및 영양인자 발현의 증가에서 비롯된 결과로 해석되 고 있다(Lemaire et al., 2000; Cotman et al., 2002).
노화과정 또는 퇴행성 뇌질환에 의한 신경세포의 소실 에서 세포사멸은 중요한 역할을 담당하고 있다. 해마에서 의 신경세포 세포사멸은 학습능력과 기억력의 장애를 일 으키며(Sun et al., 2009), 노화는 해마의 신경세포의 사멸을 증가시킨다고 알려져 있다(Pollack et al., 2002; Mattson et al., 2006). 본 연구에서는 세포사멸 특징을 확인할 수 있는 DNA 분절을 확인하는 TUNEL 방법과 세포사멸 과정 중 실행자 역할을 담당하는 caspase-3에 대한 면역조직화학법 을 실시하여 확인하였다(Reed, 2000). 실험 결과, 노화 쥐의
해마에서는 TUNEL-양성 세포 수와 caspase-3-양성 세포 수 가 젊은 쥐와 비교해 증가하였으며, 트레드밀 운동에 의해 이들 수의 유의한 감소가 나타났으나 성별에 따른 차이는 나타나지 않았다. 그리고 젊은 쥐에서 트레드밀 운동은 TUNEL-양성 세포 수와 caspase-3-양성 세포 수에는 성별에 관계없이 유의한 변화를 유발시키지 않았다. 이러한 결과 는 노화에 의해 세포사멸이 증가하는 것을 의미하며, 트레 드밀 운동이 노화에 유도된 세포사멸을 억제하는 효과가 있다는 것을 의미한다. 운동은 노화에 의해 유발되는 세포 사멸을 억제하여 신경학적 장애를 개선하고 뇌의 신경세 포 생성을 촉진하여 기능적 회복을 촉진하는 것으로 알려 져 있다(Albeck et al., 2006; Arida et al., 2007).
노화에 의한 해마에서의 신경세포 생성의 감소는 기억 력의 감퇴와 밀접한 관련이 있다 (Bizon et al., 2004; Drapeau et al., 2007). 본 실험에서 해마 부위의 신경세포 생성은 노 화에 의해 감소되었으며, 젊은 쥐와 노화 쥐에 트레드밀 운동을 실시한 결과 운동에 의하여 신경세포 생성이 증가 되었다. 그러나 운동에 의한 신경세포 생성의 효과는 젊은 쥐와 노화 쥐 모두에서 성별에 따른 차이는 나타나지 않았 다. 이러한 결과는 젊은 쥐와 노화쥐에서의 운동은 성별에 관계없이 해마 신경세포생성을 증가시킨다는 것을 의미한 다. Galea et al.(2006)은 신경 세포생성이 성별에 의하여 영 향을 받는다고 하였으나, 이러한 성별의 차이에 따른 신경 세포 생성은 설치류에 한정되어 나타난다고 하였다. 예를 들어 젊은 수컷 쥐는 암컷 쥐 보다 신경세포 생성이 높았 으며, 노화된 암컷 쥐보다 노화된 수컷 쥐에서 신경세포 생성이 더욱 높았으나, 성인기 신경세포의 생존율은 성별 에 따른 차이가 나타나지 않았다(Perfilieva et al., 2001).
Myostatin은 지방생성과 심장근 성장과정에 영향을 미친 다고 알려져 있다(Artaza et al., 2005; McKoy et al., 2007).
Myostatin은 피질판(cortical plate)을 포함한 대뇌피질과 해마 에서 발현되는데 그 기능은 잘 알려져 있지 않다. 그러나 myostatin이 신경세포 생성을 억제할 수 있음이 여러 연구 에서 제시되었다(Buckwalter et al., 2006; Wachs et al., 2006).
본 연구에서 노화 쥐는 젊은 쥐와 비교해 해마에서 myostatin의 발현이 높았으며, 트레드밀 운동은 성별 관계 없이 젊은 쥐와 노화 쥐의 myostatin 발현을 억제시켰다. 이 러한 결과는 노화에 의해 유도된 해마 myostatin 발현의 증 가는 신경세포 생성을 억제시키는 역할을 했으며 트레드 밀 운동에 의하여 myostatin 발현이 억제됨으로 신경세포 생성이 다시 증가되어 공간기억력 향상에 도움을 준 것으
로 생각된다.
Follistatin은 근육에서는 근육량을 증가시키는 중요한 변 인으로 알려져 있으며 간세포의 생성을 촉진하는 인자로 서 작용한다는 것이 보고되어 주목을 받고 있다(Kogre et al., 1995). 이것은 activin이 간세포의 세포사멸을 유도하여 간세포 생성을 억제하는 상태를 follistatin이 해제하기 때문 이라고 알려져 있다(Kogre et al., 1995). Follistatin의 기능은 조직마다 변화양상이 매우 다르며 복잡한데, follistatin은 activin signaling pathway를 억제하는 물질로 세포들의 분화 증식과 수상돌기 가시(dendritic spine) 형태를 조절한다 (Inokuchi et al., 1996; Shoji-Kasai et al., 2007). 그러나 근육에 서의 기능과는 달리 뇌에서 follistatin은 long-term potentia- tion (LTP)과 기억력을 억제하는 역할을 하며, 또한 follistatin 의 과발현은 해마 신경세포생성을 억제하는 역할을 한다 고 알려져 있다(Andreasson et al., 1995; Inokuchi et al., 1996).
본 연구에서 노화 쥐는 젊은 쥐와 비교해 해마에서의 follistatin 발현이 높았으며, 트레드밀 운동은 성별에 관계없 이 노화 쥐의 follistatin 발현을 억제시켰다. 이러한 결과는 노화에 의해 유도된 해마에서의 follistatin 발현 증가는 신 경세포 생성을 억제시켰으며, 트레드밀 운동에 의하여 follistatin 발현이 억제됨으로써 신경세포 생성이 다시 증가 되어 공간기억력을 향상시켰음을 의미한다.
본 연구를 통하여 노화에 의해 해마에서 myostatin과 follistatin의 증가가 신경세포 생성을 억제하여 공간기억력 을 감퇴시킴을 알 수 있었고, 트레드밀 운동은 myostatin과 follistatin의 발현을 억제함으로써 신경세포 생성을 증가시 켜 노화에 의해 감퇴된 공간기억력을 회복시켜주는 효과 가 있음을 확인 할 수 있었다. 그러나 운동의 효과는 노화 쥐와 젊은 쥐 모두에서 성별의 차이에 따른 효과의 차이는 없었다. 본 연구를 통하여 트레드밀 달리기는 해마에서 myostatin과 follistatin의 발현을 억제함으로써 신경세포 생 성을 증가시켜 노화에 의한 기억력 감퇴를 경감시킴을 알 수 있었다.
참 고 문 헌
Albeck DS, Sano K (2006) Mild forced treadmill exercise enhances spatial learning in the aged rat. Behav. Brain Res. 168:345-348.
Andreasson K, Worley PF (1995) Induction of b-A activin expression by synaptic activity and during neocortical development.
Neuroscience 69:781-796.
Arida RM, Scorza CA, Scorza FA et al. (2007) Effects of different
types of physical exercise on the staining of parvalbumin-positive neuron in the hippocampal formation of rats with epilepsy. Prog.
Neuropsychopharmacol. Biol. Psychiatry 31:814-822.
Artaza JN, Bhasin S, Magee TR et al. (2005). Myostatin inhibits myogenesis and promotes adipogenesis in C3H 10T(1/2) mesen- chymal multipotent cells. Endocrinology 146:3547-3557.
Bilezikjian LM, Corrigan AZ, Blount AL et al. (1996) Pituitary follistatin and inhibin subunit messenger ribonucleic acid levels are differentially regulated by local and hormonal factors.
Endocrinology 137:4277-4284.
Bizon JL, Lee HJ, Gallagher M (2004) Neurogenesis in a rat model of age related cognitive decline. Aging Cell 3:227-234.
Buckwalter MS, Yamane M, Coleman BS et al. (2006) Chronically increased transforming growth factor-β1 strongly inhibits hippo- campal neurogenesis in aged mice. Am. J. Pathol. 169:154-164.
Colcombe SJ, Erickson KI, Raz N et al. (2003) Aerobic fitness reduces brain tissue loss in aging humans. J. Gerontol. A. Biol.
Sci. Med. Sci. 58:M176-M180.
Cotman CW, Berchtold NC (2002) Exercise: a behavioral inter- vention to enhance brain health and plasticity. Trends Neurosci.
25:295-301.
Drapeau E, Montaron MF, Aguerre S et al. (2007) Learning-induced survival of new neurons depends on the cognitive status of aged rats. J. Neurosci. 27:6037-6044.
Eriksson PS, Perfilieva E, Björk-Eriksson T et al. (1998) Neurogene- sis in the adult human hippocampus. Nature Med. 4:1313-1317.
Fabel K, Kempermann G (2008) Physical activity and the regulation of neurogenesis in the adult and aging brain. Neuromolecular Med.
10:59-66.
Fordyce, DE, Wehner JM (1993) Physical activity enhances spatial learning performance with n associated alteration in hippocampal protein kinase C activity in C57BL/6 and BA/2 mice. Brain Res.
619:111-119.
Fuchs E, Gould E (2000) Mini-review: in vivo neurogenesis in the adult brain: regulation and functional implications. Eur. J. Neurosci.
12:2211-2214.
Galea LA, Spritzer MD, Barker JM et al. (2006) Gonadal hormone modulation of hippocampal neurogenesis in the adult. Hippo- campus 16:225-232.
Gavrieli Y, Sherman Y, Ben-Sasson SA (1992) Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell. Biol. 119:493-501.
Gould E, Tanapat P (1999) Stress and hippocampal neurogenesis.
Biol. Psychiatry 46:1472-1479.
Inokuchi K, Kato A, Hiraia K et al. (1996) Increase in activin bA mRNA in rat hippocampus during long-term potentiation. FEBS Lett. 382:48-52.
Is M, Comunoglu NU, Comunoglu C et al. (2008) Age‐related
changes in the rat hippocampus. J. Clin. Neurosci. 15:568-574.
Jarrard LE (1995) What does the hippocampus really do? Behav.
Brain Res. 71:1-10.
Johansson BB, Ohlsson AL (1996) Environment, social interaction, and physical activity as determinants of functional outcome after cerebral infarction in the rat. Exp. Neurol. 139:322-327.
Kambadur R, Sharma M, Smith TP et al. (1997) Mutations in myostatin (GDF8) in double-muscled Belgian Blue and Piedmon- tese cattle. Genome Res. 7:910-916.
Kerr JF, Wyllie AH, Currie AR (1972) Apoptosis: a basic biological phenomenon with wide ranging implications in tissue kinetics.
Br. J. Cancer 26:239-257.
Kim YP, Kim H, Shin MS et al. (2004) Age-dependence of the effect of treadmill exercise on cell proliferation in the dentate gyrus of rats. Neurosci. Lett. 355:152-154.
Ko IG, Kim SE, Shin MS et al. (2011) Treadmill exercise reduces lipopolysaccharide-induced apoptotic neuronal cell death in the hippocampus of the young and old rats. Korean J. Str. Res.
19:97-105.
Kogure K, Omata W, Kanzaki M et al. (1995) A single intraportal administration of follistatin accelerates liver regeneration in parti- ally hepatectomized rats. Gastroenterology 108:1136-1142.
Kuhn HG, Dickinson‐Anson H, Gage FH (1996) Neurogenesis in the dentate gyrus of the adult rat: age‐related decrease of neuronal progenitor proliferation. J. Neurosci. 16:2027-2033.
Lee SJ, McPherron AC (2001) Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 98:9306-9311.
Lemaire V, Koehl M, Le Moal M et al. (2000) Prenatal stress produces learning deficits associated with an inhibition of neurogenesis in the hippocampus. Proc. Natl. Acad. Sci. USA 97:11032-11037.
Li Y, Chopp M, Powers C et al. (1997) Apoptosis and protein expre- ssion after focal cerebral ischemia in rat. Brain Res. 765:301-312.
Mattson MP, Magnus T (2006) Ageing and neuronal vulnerability.
Nat. Rev. Neurosci. 7:278-294.
McKoy G, Bicknell KA, Patel K et al. (2007) Developmental expression of myostatin in cardiomyocytes and its effect on foetal and neonatal rat cardiomyocyte proliferation. Cardiovasc. Res.
74:304-312.
Patel K (1998) Follistatin. Int. J. Biochem. Cell Biol. 30:1087-1093.
Patrylo PR, Williamson A (2007) The effects of aging on dentate circuitry and function. Prog. Brain Res. 163:679-696.
Perfilieva E, Risedal A, Nyberg J et al. (2001) Gender and strain influence on neurogenesis in dentate gyrus of young rats. J.
Cereb. Blood Flow Metab. 21:211-217.
Pollack M, Phaneuf S, Dirks A et al. (2002) The role of apoptosis
in the normal aging brain, skeletal muscle, and heart. Ann. NY
Acad. Sci. 959:93-107.
= 국문초록 =
성별에 따른 트레드밀 달리기가 노화 쥐의 공간기억력에 미치는 영향을 연구하였다. 2개월 된 암컷 쥐와 수컷 쥐를 젊은 쥐로 하였고, 15개월 된 암컷 쥐와 수컷 쥐를 노화 쥐로 하였다. 실험동물은 젊은 암컷 대조군(N=6), 젊은 암컷 운동군(N=6), 젊은 수컷 대조군(N=6), 젊은 수컷 운동군(N=6), 노화 암컷 대조군(N=6), 노화 암컷 운동군(N=6), 노 화 수컷 대조군(N=6), 노화 수컷 운동군(N=6)의 8군으로 나누었다. 운동군은 소형동물용 트레드밀에서 하루 일회 30분간 4주 동안 트레드밀 달리기를 실시하였다. 노화 쥐는 젊은 쥐와 비교하였을 때 공간기억력의 감퇴를 나타내었 고 해마에서 신경세포 생성은 억제된 반면 세포 사멸은 증가되었다. 노화 쥐에서 트레드밀 달리기를 시행하였을 때, 공간 기억력의 감퇴가 억제되었고 해마의 신경세포 생성은 증가된 반면 세포사멸은 억제되었다. 해마에서의 myostatin과 follistatin 발현은 뇌화 쥐에서 억제되었으나, 트레드밀 달리기에 의하여 증가되었다. 그러나 트레드밀 달 리기의 이러한 효과는 성별에 따른 차이는 나타나지 않았다. 본 연구를 통하여 트레드밀 달리기는 해마에서 myostatin과 follistatin의 발현을 억제함으로써 신경세포 생성을 증가시켜 노화에 의한 기억력 감퇴를 경감시킴을 알 수 있었다.
중심단어: 노화, 트레드밀 달리기, 공간기억력, 신경세포 생성, 세포사멸