3T3-L1 지방세포에서 DHA와 EPA의 지질축적 억제 효능 및 히스톤아세틸전달효소 활성 저해
정상원․한정수․정민유 한국식품연구원 식품기능연구본부
Hypolipidemic Effect of Omega-3 Fatty Acids in 3T3-L1 Preadipocytes via Inhibition of Histone Acetyltransferase Activity
Sangwon Chung, Jung Su Han, and Min-Yu Chung Research Division of Food Functionality, Korea Food Research Institute
ABSTRACT Obesity causes several metabolic and chronic diseases, including type 2 diabetes and cardiovascular diseases. Among the omega-3 fatty acids, various health benefits of docosahexaenoic acid (DHA; C22:6n-3) and eicosa- pentaenoic acid (EPA; C20:5n-3) have been elucidated. This study examined the hypolipidemic effect of DHA and EPA and the epigenetic regulation of DHA and EPA through histone acetyltransferases (HAT) activity. DHA and EPA inhibited the in vitro HAT activity, and non-toxic levels of DHA and EPA significantly attenuated lipid accumu- lation in 3T3-L1 preadipocytes treated with MDI (IBMX+dexamethasone+insulin) and insulin. DHA and EPA regu- lated the expression of lipogenic, cholesterol-related, or triacylglycerol synthesis-related genes. Indeed, DHA and EPA significantly enhanced PPARα and PPARγ2 in 3T3-L1 preadipocytes treated with MDI and insulin. Although DHA also increased SREBP1α and LXR significantly, EPA significantly attenuated the expression of the SREBP1α and LXR genes. In addition, a lower level of DHA increased AGPAT2 gene expression significantly, which was reduced significantly by the EPA treatment in 3T3-L1 preadipocytes treated with MDI and insulin. These results suggest that the hypolipidemic activity of EPA and DHA is exerted via different gene expression regulation, which is likely to be associated with HAT activity inhibition.
Key words: DHA, EPA, HAT, 3T3-L1 preadipocytes, AGPAT2
Received 18 September 2020; Accepted 27 September 2020 Corresponding author: Min-Yu Chung, Korea Food Research Insti- tute, Jeonbuk 55365, Korea, E-mail: [email protected]
서 론
비만은 신체활동, 기초대사 등의 에너지 소비보다 과도한 에너지 섭취가 장기적으로 지속되어 체내에 체지방이 과도 하게 축적된 상태를 말한다. 비만은 심혈관계 질환, 당뇨병, 암 등 여러 질환의 주요 원인이 되므로 전 세계적인 건강문제 로 주목을 받고 있다(Spiegelman과 Flier, 2001; Visscher 과 Seidell, 2001). 비만과 비만으로 인한 여러 만성질환의 예방을 위해 영양적으로 균형 잡힌 식사와 생리활성이 큰 성 분이 많이 함유된 식품의 섭취가 중요하다(Prostek 등, 2016).
최근에는 지방축적과 관련하여 히스톤 아세틸화(acety- lation)와 같은 후성유전적 조절이 지방합성을 비롯한 다양 한 질병 발병과 연관이 있다는 연구 결과가 보고되고 있다.
예를 들면, 히스톤아세틸전달효소(histone acetyltrans- ferase, HAT)인 CREB-binding protein(CBP)/p300은 지 질축적에 관여하는 전사인자인 sterol regulatory element
binding protein 1c(SREBP-1c)의 아세틸화를 유도하여 지 방합성 관련 유전자의 발현을 촉진한다(Wang 등, 2015).
SREBP-1c의 아세틸화에 의해 fatty acid synthase(FAS) 와 glycerol-3-phosphate acyltransferase(GPAT)와 같 은 유전자의 발현이 증가하는데, 이들은 지방산 합성 기전에 관여한다고 알려져 있다(Yang 등, 2001). 특히, GPAT는 중 성지방과 인지질의 생합성 첫 단계에 관여하는 유전자이다.
GPAT의 isoform인 GPAT3와 GPAT4는 지방조직에서 발 현되며, 이들의 기능은 인슐린이 관여하는 인산화에 의해 조 절된다고 밝혀져 비만, 당뇨, 이상지질혈증 등의 치료 목표 물질로 주목받고 있다(Shan 등, 2010).
지방의 과도한 섭취와 비만, 대사질환 발병 위험은 높은 상관관계를 보이는데, 질병에 대한 영향은 지방을 구성하고 그 질을 결정하는 지방산의 종류에 따라 다르다(Hill 등, 2000; Imamura 등, 2016; Zhu 등, 2019). Guasch-Ferre 등(2015)에 따르면, 불포화지방산의 섭취는 심혈관계 질환 의 위험을 낮추는 반면 포화지방산 및 트랜스 지방산의 섭취는 심혈관계 질환의 위험을 높인다(Guasch-Ferre 등, 2015).
오메가-3 불포화지방산의 섭취는 중성지방을 감소시켜 이 상지질혈증을 개선하고 항염증 작용과 혈압강하를 통해 혈
관 기능을 개선시킨다고 밝혀졌다(Lorente-Cebrian 등, 2013). 오메가-3 지방산 중 eicosapentaenoic acid(EPA;
C20:5n-3)와 docosahexaenoic acid(DHA; C22:6n-3)가 지질대사 개선에 작용하는 기전은 여러 연구를 통해 입증되 었다. DHA와 EPA는 미토콘드리아의 생합성과 베타산화를 유도하여 지방세포에서 지방축적을 감소시킨다(Flachs 등, 2009). 3T3-L1 지방전구세포를 이용하여 DHA와 EPA가 지방세포 분화에 미치는 영향을 비교한 연구에 따르면, EPA 가 아닌 DHA는 아디포넥틴 분비를 증가시키고 염증 신호 기전을 억제시킨다(Murali 등, 2014). 또한, 고지방식이와 DHA와 EPA를 함께 먹인 쥐의 백색지방과 갈색지방 조직의 지질축적과 염증 촉진성 아라키돈산 수준이 고지방식이만 먹인 쥐에 비해 감소하였다(Soni 등, 2019). DHA는 혈중 중 성지질을 낮추고 죽상동맥경화증과 동맥혈전 생성을 예방 하여 심장질환 예방에 효과가 있다(Connor, 2000; Mozaf- farian과 Wu, 2011). 또한 DHA는 망막과 뇌에 존재하며 신경계의 발달, 광학 정보의 전달, 로돕신 재생에도 도움을 준다(Wainwright, 2002; SanGiovanni와 Chew, 2005). 최 근에는 알츠하이머의 촉진을 늦추고 신경세포 사멸을 억제 한다고 보고(Kondo 등, 2013)되었으며, 그 외에 항염증 효 능도 보고되었다(Schwab 등, 2007). 종합적으로 DHA와 EPA는 지질축적 억제뿐만 아니라 다양한 생체 기능 조절에 관여한다.
하지만 DHA와 EPA의 비만 혹은 대사질환에서 후성 유 전학적 조절 기능에 대해서는 연구된 바가 없다. 본 연구에 서는 3T3-L1 지방전구세포에 MDI와 인슐린을 처리하여 분화시키고, 분화 기간에 오메가-3 지방산인 DHA와 EPA 를 각각 50, 100, 200 μM 농도로 처리한 후, 지방축적 억제 관찰, 관련 유전자를 측정하였고 후성 유전조절 관여 효소 중 대표적인 히스톤아세틸전달효소의 활성을 측정하였다.
재료 및 방법
실험재료
본 실험에 사용된 insulin, 3-isobutyl-1-methylxanthine (IBMX), dexamethasone, EPA, DHA, Oil Red O 염색 시 약, MTT, bovine serum albumin(BSA)는 Sigma-Aldrich (St. Louis, MO, USA)에서 구입하여 사용하였다. Dulbec- co’s modified Eagle’s medium(DMEM), fetal bovine se- rum(FBS)은 Welgene Inc.(Gyeongsan, Korea), isopro- panol은 EMD Millpore(Billerica, MA, USA)에서 구입하였 다.
DHA, EPA-BSA complex 제조
DHA와 EPA를 각각의 분자량을 고려하여 95% 에탄올에 녹인 stock(10 mM)을 준비하였다(45°C 핫플레이트 이용).
지방산 stock을 질소 농축기(Organomation Associates, Inc., Berlin, MA, USA)를 이용하여 농축시켰다. Fatty acid
-free BSA를 DMEM과 1 M HEPES에 용해시킨 BSA- DMEM 용액을 농축된 지방산이 들어있는 유리튜브에 넣어 완전히 용해시켰다. 0.22 μm 필터를 이용하여 걸러낸 후 희석하여 세포에 처리하였다.
세포배양 및 지질축적 유도
3T3-L1 지방전구세포는 American Type Culture Col- lection(ATCC, Manassas, VA, USA)에서 분양받아 사용 하였다. 세포는 10% calf serum, antibiotics, antimycotic 이 함유된 DMEM 배지를 이용하여 37°C, 5% CO2 조건에서 배양하였다. 세포의 분화를 유도하기 위해 6-well plate에 분주 후 100% confluent 상태가 되기까지 2일간 배양하였 다. 세포가 confluent 상태가 된 후 추가로 2일 동안 배양한 다음 10% FBS가 첨가된 MDI 배지(조성: 0.5 mM IBMX, 10 μg/mL insulin, 0.5 μM dexamethasone)로 옮겨 DHA 와 EPA를 처리하였다. 3일 후부터는 10 μg/mL insulin이 포함된 DMEM 배지로 이틀에 한 번씩 교환하여 4일간 추가 로 배양하였다.
Oil Red O 염색 및 정량
3T3-L1 세포의 지방축적량을 Oil Red O 염색법을 통해 정성 및 정량 분석하여 DHA와 EPA의 항비만 효과를 평가하 였다. 3T3-L1에 DHA와 EPA를 처리하고 7일간 분화시켰 다. 세포를 PBS로 세척하고, 3.7%의 formaldehyde로 실온 에서 고정시켰다. 고정된 세포는 60%의 isopropanol에 용 해된 0.2% Oil Red O 용액으로 1시간 동안 실온에서 염색하 였고 염색 후에는 증류수를 이용하여 염색 시약을 완전히 제거하였다. 염색된 세포는 현미경을 이용하여 관찰한 다음 스캐너를 이용하여 사진을 얻었다. Oil Red O 염색의 정량분 석은 200 μL의 isopropanol을 처리하여 세포 내 염색 시약 을 녹인 후 96-well plate에 100 μL씩 옮겨 담아 510 nm에 서 흡광도를 측정하여 lipid accumulation을 계산하였다.
세포 활성 측정
3T3-L1 지방전구세포를 10%의 FBS를 포함한 DMEM 으로 96-well plate에 분주하고 confluence에 도달하였을 때 MDI와 함께 DHA와 EPA를 50 μM, 100 μM 농도로 처리 하여 48시간 동안 배양하였다. MTT 용액을 첨가하고 37°C 에서 2시간 동안 추가로 배양한 후, MTT 용액을 제거하고 dimethyl sulfoxide(DMSO)를 추가하여 37°C에서 10분간 배양하였다. 이후 생성된 blue-formazan 결정을 DMSO에 용해시켰다. Microplate reader(Molecular Devices, San Jose, CA, USA)를 이용하여 575 nm의 흡광도에서 세포 생존율을 측정하였다.
In vitro HAT 활성 측정
DHA와 EPA에 의한 in vitro HAT 활성을 측정하기 위해 HAT activity assay kit과 HeLa cell nuclear extract(NE;
Table 1. Primer sequences for quantitative real-time PCR
Gene Forward (5′-3′) Reverse (5′-3′) Product size
PPARα PPARγ2 SREBP1α
LXR AGPAT2
GPAT4 GAPDH
GTGTACGACAAGTGTGATCG [Tm 56.61]
ACCCAGAGCATGGTGCCTTC [Tm 62.49]
AGATGTGCGAACTGGACACA [Tm 59.61]
GAGAAGCTGGTGGCTGCCCA [Tm 64.92]
AGCGGACAGAAGAAACTGGA [Tm 58.95]
TGTCTGGTTTGAGCGTTCTG [Tm 58.42]
AGGTCGGTGTGAACGGATTTG [Tm 60.88]
GATTTGAGGTCTGCAGTTTC [Tm 54.61]
CCATCACGGAGAGGTCCACA [Tm 61.26]
CATCTTTAAAGCAGCGGGTG [Tm 57.45]
AGCTGTAGGAAGCCAGGGAG [Tm 60.98]
TGAAGTAGACACCCCCAAGG [Tm 58.64]
TTCTGGGAAGATGAGGATGG [Tm 56.29]
TGTAGACCATGTAGTTGAGGTCA [Tm 58.59]
212 bp 1,223 bp 80 bp 200 bp 1,212 bp 3,399 bp 123 bp PPARα, peroxisome proliferator-activated receptor alpha; PPARγ2, peroxisome proliferator-activated receptor gamma 2; SREBP1α, sterol regulatory element-binding protein 1α; LXR, liver X receptor; AGPAT2, acylglycerol-3-phosphate acyltransferase 2; GPAT4, glycerol-3-phosphate acyltransferase.
BioVision Biotechnology, Milpitas, CA, USA)를 구매하 여 판매사의 실험방법에 따라 측정하였다. 96-well plate에 16 μg의 NE(4 µL)와 DHA 혹은 EPA를 0, 25, 50, 100 μM의 농도로 넣어준 후, 최종 볼륨이 40 µL가 되도록 증류 수를 넣었다. Background 판별을 위하여 NE 및 소재 등을 포함하지 않는 증류수 40 μL만 넣은 well을 준비하였다. 상 업적으로 제공되는 substrate인 histone H3 tail(5 μL), co- factor인 acetyl-CoA(5 μL), NADH generating enzyme (8 μL)을 2×HAT assay buffer(50 μL)에 넣어 섞은 후, 앞서 준비된 well에 68 μL씩 분주하여 넣었다. Microplate reader(Molecular Devices)를 이용하여 440 nm 파장에서 OD값을 측정하고 이를 T0 HAT activity로 간주하였다. 30 분 동안 37°C에서 incubation한 후 동일한 파장에서 OD값 을 측정하고 이를 T30 HAT activity라 하였다. 최종 변화한 HAT 활성을 계산하기 위해 T0, T30의 HAT activity OD값 에서 T0와 T30 background OD값을 빼주어 background 를 보정하고 Δ30분의 HAT activity(T30의 OD값-T0의 OD값)를 계산한 다음, NE의 HAT activity만을 보여주는 positive control 반응 well의 활성 대비 DHA, EPA에 의한 HAT 활성 억제 정도를 상대적으로 계산하였다.
Total RNA 추출과 qRT-PCR
3T3-L1 지방전구세포에 MDI, insulin, DHA, EPA를 위 에서 언급한 농도 및 날짜별로 처리한 후, cold DPBS로 2회 세척하고 원심분리하여 얻어진 세포 pellet에서 total RNA 를 추출하였다. 총 4 μg의 RNA를 이용하여 cDNA를 합성하 였다. qRT-PCR 분석은 SYBR Green PCR Master mix reagent(Thermo Fisher Scientific, Waltham, MA, USA) 를 이용하여 다음과 같은 조건으로 시행하였다(Bio-Rad CFX Connect Real-Time PCR Detection System, Bio- Rad Laboratories, Hercules, CA, USA). 모든 결과는 GADPH에 따라 상대적으로 정량화되었다. PCR은 95°C에 서 30초, 56°C에서 30초, 72°C에서 30초 40 cycle로 시행 되었으며, 마지막 72°C에서 10분간 추가로 반응하여 안정 화시켰다. 증폭의 모든 데이터는 매 cycle의 72°C, 30초 단계에서 수집되었다. 본 분석에 사용된 프라이머는 Table 1과 같다. 모든 반응은 3회 반복 시행되었으며, 상대적 발현
및 SD값은 comparative method를 사용하여 계산하였다.
통계분석
본 실험결과는 GraphPad Prism(version 7.0, GraphPad Software, San Diego, CA, USA)을 사용하여 통계적 검증 을 실시하였다. 모든 결과 값은 3회 반복에 대한 평균±표준 편차로 표시하였고, 대조군과 실험군 간의 유의성은 Stu- dent’s t-test를 사용하여 검증하였다.
결과 및 고찰
DHA 및 EPA의 in vitro HAT 활성 억제
오메가-3 지방산 중 지방축적 억제 등의 기능성이 알려진 DHA와 EPA를 BSA와 conjugation시켜 농도별(25, 50, 100 μM)로 준비하였다. In vitro HAT 활성 측정은 colori- metric HAT 활성 측정 kit(BioVision Biotechnology)을 사용하였고, 설명서의 실험법 및 선행 논문을 참고하여 실험 을 진행하였다(Chung 등, 2019). 그 결과 DHA와 EPA는 모 두 in vitro HAT 활성을 억제하였다(Fig. 1). NE 처리 양성 대조군의 HAT 활성을 100%로 하여 비교했을 때, DHA를 처리한 경우 25 μM에서 100 μM까지 농도 의존적으로 HAT 활성이 감소하였다(P<0.001; Fig. 1A). EPA도 NE에 의해 증가한 HAT 활성을 억제하였으며 저농도의 EPA(25 μM) 부터 유의적으로 HAT 활성을 억제하였다(P<0.001; Fig.
1B). DHA와 EPA 모두 HAT 활성 억제능이 있고, 특히 저농 도 처리군을 비교했을 때 EPA의 HAT 활성 억제능이 DHA 보다 크다는 것을 알 수 있었다.
후성유전적 조절과 여러 만성질병과의 상관관계에 관한 연구는 많이 발표되어왔다. Histone deacetylase(HDAC) 혹은 DNA methyltransferase 억제제 외의 HAT 활성 억제 또한 후성유전적 조절과 질병 예방 및 치료 연구에서 중요한 역할을 한다(Wapenaar와 Dekker, 2016; Manzo 등, 2009).
히스톤 단백질의 아세틸화는 특정 유전화의 활성 조절과 관 련이 있다(de Moraes Maciel 등, 2004). HAT 활성을 지닌 p300은 coactivator로 작용하며 프로모터에서 특정 유전자 의 전사를 활성화시킨다. p300/CBP complex는 SREBP- 1c나 PPARγ를 포함한 다양한 전사인자의 활성을 조절한
A B
Fig. 1. In vitro HAT activity regulated by DHA (A) or EPA (B) in MDI-treated 3T3-L1 preadipo- cytes. DHA or EPA was treated 25, 50, and 100 μM during MDI-mediated adipognesis of 3T3-L1 preadipocytes. All data are expressed as mean±SD from the samples of each group. Asterisks (*) in- dicate significant differences compared between NE only treated group and DHA or EPA treated groups (*P<0.05, **P<0.01, ***P<0.001).
A
B
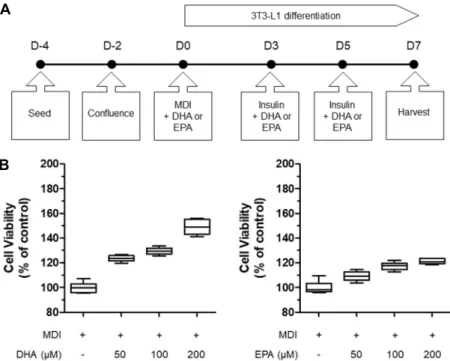
Fig. 2. Schematic description of MDI, insulin, DHA or EPA treatment during 3T3-L1 pre- adipocytes differentiation (A). Cell viability in MDI-treated 3T3-L1 preadipocytes supple- mented with none, DHA (50, 100, 200 μM) or EPA (50, 100, 200 μM) (B).
다. 포도당에 의해 활성화되기도 하는 p300/CBP complex 의 증가는 여러 lipogenic 유전자의 발현을 증가시켜 지방간 을 더욱 악화시키게 된다(Giandomenico 등, 2003; Gelman 등, 1999; Tian 등, 2013).
HAT 활성 억제제가 질환의 발병을 예방 혹은 늦출 수 있 을 것이라는 가설하에 HAT 활성 억제제로서 플라보노이드 의 역할에 관한 연구가 많이 진행되고 있다. 녹차 카테킨 중 epigallocatechin gallate(EGCG)는 대표적인 HAT inhibi- tor로 p300에 의해 유도된 p65의 아세틸화를 억제시켜 NF- kappaB 활성을 억제하고, 이로 인해 림프구 형질전환에서 중요한 역할을 한다는 연구 보고가 있다(Choi 등, 2009).
선행논문에서 탄닌산 또한 HAT inhibitor로 작용함을 밝힌 바 있다(Chung 등, 2019). 탄닌산은 p300이 FASN, ACLY 프로모터의 SRE region에 결합하는 것을 방해하여 아세틸 화를 막고, FASN, ACLY 관련 유전자의 발현을 감소시켜 지방간 유도 과정 중 지방축적을 억제하였다(Chung 등, 2019). 이외에도 식품 성분으로서 오미자(Chung 등, 2017),
도토리(Hwang 등, 2017), 냉이(Choi 등, 2017) 등 추출물 의 지방간 억제능이 HAT 활성 억제를 통해 일어나며 곰취 추출물 또한 HAT 활성 억제제로서 p65 아세틸화를 저해하 고 p65의 핵 내 이동을 막아 LPS 처리한 RAW264.7 세포에 서 사이토카인 증진 등의 염증반응을 억제하였다(Kim 등, 2019). 종합적으로 천연물, 식품소재 및 플라보노이드류의 HAT 활성과 이로 인한 질환의 예방 가능성을 제시한 논문 은 다수 존재하며 관련 연구가 활발히 진행 중이다. 하지만 오메가-3 지방산의 HAT 활성 조절과 이를 통한 비만 예방 의 효능 가능성을 제시한 논문은 본 연구가 최초이며 기능성 지방산을 활용한 후성유전적 조절 및 질환 예방에 관한 지속 적인 연구가 필요하다.
3T3-L1 지방세포 분화 중 DHA와 EPA의 지방축적 억제 3T3-L1 지방전구세포의 분화 중 HAT 활성 억제를 통한 지방축적 저해에 관한 보고가 있다(Hwang 등, 2017). 본 연구에서는 HAT 활성 억제능을 보인 DHA와 EPA(Fig. 1)
A B
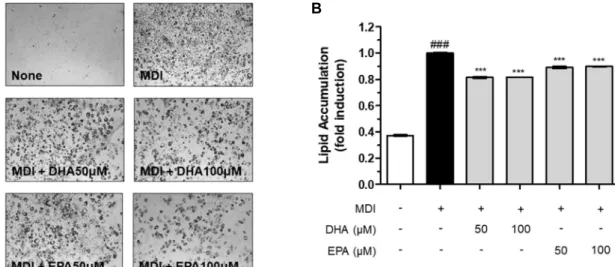
Fig. 3. Oil Red O staining of MDI-treated 3T3-L1 preadipocytes supplemented with none, DHA (50, 100 μM) or EPA (50, 100 μM) (A), which were quantified (B). All data are expressed as mean±SD from the samples of each group. Hashes (#) indicate significant differences compared to controls treated with no MDI (###P<0.001). Asterisks (*) indicate significant differences compared groups between MDI-treated controls and DHA or EPA-treated groups (***P<0.001).
의 지방축적 억제를 확인하기 위해 MDI와 인슐린으로 3T3- L1 지방전구세포를 분화시켜 지방축적을 유도하고 Fig. 2A 에서와 같이 분화 유도 중 EPA와 DHA를 각각 처리하였다.
DHA(~200 μM)와 EPA(~200 μM)에 의한 세포 활성 조절 정도를 MTT로 측정하였다. DHA와 EPA 모두 세포 활성을 감소시키지 않았다(Fig. 2B). DHA와 EPA 처리는 MDI 및 인슐린을 처리한 대조군 대비 세포 활성을 오히려 증가시켜 3T3-L1 지방전구세포의 분화 과정 중 DHA와 EPA에 의한 독성유발은 일어나지 않는 것으로 간주하고 실험을 진행하 였다.
MDI와 인슐린을 동시에 처리했을 때 MDI를 처리하지 않 은 음성대조군에 비해 약 5배의 지방축적이 일어남을 관찰 할 수 있었고 이러한 증가는 통계적으로도 유의하였다(P<
0.001; Fig. 3A, B). DHA와 EPA를 각각 50, 100 μM의 농도 로 지방세포 분화 과정 중 처리한 결과, DHA와 EPA 두 지방 산 모두 MDI에 의한 지방축적을 유의적으로 감소시켰다 (P<0.001; Fig. 3A, B). MDI 양성 대조군 대비 지방 축적률 은 DHA에 의해 약 20%, EPA에 의해 약 15%의 감소율을 보였다(Fig. 3B).
세포모델을 활용한 연구에서 지방축적 억제의 기능성은 여러 메커니즘을 통해 일어난다. 예를 들어, 3T3-L1 지방전 구세포의 분화 과정 중 EPA와 DHA는 lipolytic 유전자의 발 현을 증가시켜 지방축적을 억제한다(Kim 등, 2006). EPA 와 DHA는 또한 lipolysis(Barber 등, 2013)와 긴사슬 지방 산의 베타산화에 관여하는 여러 유전자의 발현을 증가시켜 지질축적을 억제한다(Lee 등, 2008). 하지만 동물 모델을 이용한 몇몇 기능성 연구들은 상반된 결과를 제시하고 있다.
DHA만(EPA는 효과 없음) 백색지방조직의 갈색화를 촉진 시킨다는 보고가 있다(Zhuang 등, 2019). 또한 ob/ob 마우 스에서는 EPA만(DHA는 효과 없음) 고지방-고자당 식이로
비만이 유도된 마우스 모델에서 체중 증가를 억제하는데, 이는 포도당의 항상성 조절을 통해 일어난다는 연구 결과로 보고된 바 있다(Pinel 등, 2016). 종합적으로 DHA나 EPA 에 의한 지방축적 억제능은 연구되어 왔지만, 현재까지의 연구 결과들을 종합하여 두 오메가-3 지방산의 지방축적 억 제능을 명확히 결론 내리기는 어렵다.
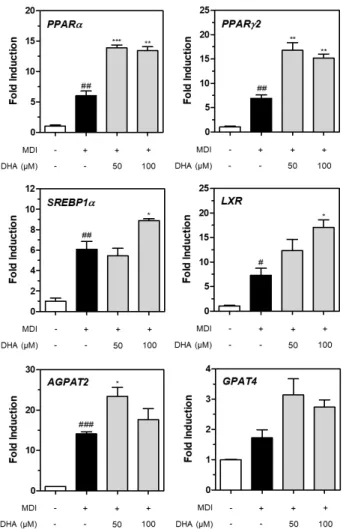
3T3-L1 지방세포 분화에서 DHA의 유전자 발현 조절 DHA와 EPA는 MDI와 인술린을 이용한 3T3-L1 지방세 포의 분화 과정 중 증가한 지방축적을 유의적으로 감소시켰 다. 이에 DHA와 EPA의 지방 억제 관련 유전자의 조절을 관찰하고자 qRT-PCR을 수행하였다. DHA는 3T3-L1 지 방전구세포에서 MDI 처리에 의해 증가한 PPARα와 PPARγ 2의 유전자 발현량을 유의적으로 증가시켰다(P<0.01; Fig.
4, upper panel). 대식세포에서 주로 발현한다고 알려진 SREBP1α 유전자는 MDI 처리에 의해 그 발현량이 증가하 였고, 고농도의 DHA(100 μM) 처리에 의해 유의적으로 증 가하였다(P<0.05). Liver X receptor(LXR) 유전자 또한 MDI 처리에 의해 증가하였고, 고농도(100 μM) 처리에 의해 유의적으로 더욱 증가하였다(P<0.05; Fig. 4, middle pan- el). Diacylglycerol의 전구물질인 phosphatidic acid와 ly- sophosphatidic acid의 합성에 관여하는 sn-1-acylglyc- erol-3-phosphate acyltransferase(AGPAT)2와 GPAT4 의 유전자 발현량을 관찰하였을 때 두 유전자 모두 MDI 처 리에 의해 증가하였고, 저농도의 DHA(50 μM) 처리에 의해 서만 유의적으로 증가하였다(P<0.05). DHA에 의해 GPAT4 의 유전자 발현이 증가하는 경향을 보였으나, 이는 통계적으 로 유의하지 않았다(Fig. 4, lower panel).
DHA와 EPA 모두 PPARγ의 잠재적인 ligands로 알려져 있다. PPARγ는 아디포넥틴 조절의 주요 전사인자이므로
Fig. 4. Adipogenic genes expression regulated by DHA in 3T3- L1 preadipocytes differentiated by MDI. All data are expressed as mean±SD from the samples of each group. Hashes (#) in- dicate significant differences compared to controls treated with no MDI (#P<0.05, ##P<0.01, ###P<0.001). Asterisks (*) indicate significant differences compared groups between MDI-treated controls and DHA-treated groups (*P<0.05, **P<0.01, ***P<0.001).
Fig. 5. Adipogenic genes expression regulated by EPA in 3T3- L1 preadipocytes differentiated by MDI. All data are expressed as mean±SD from the samples of each group. Hashes (#) in- dicate significant differences compared to controls treated with no MDI (#P<0.05, ##P<0.01, ###P<0.001). Asterisks (*) indicate significant differences compared groups between MDI-treated controls and EPA-treated groups (*P<0.05; **P<0.01, ***P<0.001).
DHA와 EPA는 PPARγ-아디포넥틴 조절을 통해 항염증 효 능을 나타낼 수 있다(Kadowaki와 Yamauchi, 2005). 하지 만 오메가-3의 PPARγ 조절능에 대해서는 현재까지 명확히 결론짓기 어렵다. 마우스 모델을 이용한 연구에서는 오메가 -3가 많이 함유된 피쉬오일이 아디포넥틴 유전자의 발현을 증가시켰으며, 이는 PPARγ에 의존적이다(Neschen 등, 2006). Oster 등(2010)의 연구에서는 3T3-L1 지방세포에 서 EPA가 아닌 DHA만이 PPARγ의 유전자 발현을 유의적 으로 증가시킴을 밝혔다. 반면, Prostek 등(2016)의 연구에 서는 3T3-L1 preadipocyte의 분화 과정 중 PPARγ의 유전 자 발현이 EPA와 DHA 모두에 의해 감소하였다.
3T3-L1 지방세포 분화에서 EPA의 유전자 발현 조절 3T3-L1 지방세포의 분화 중 MDI 처리에 의해 유의적으 로 증가한 PPARα와 PPARγ는 EPA에 의해 증가하는 경향 을 보였다(Fig. 5, upper panel). 하지만 이는 모두 농도 의
존적으로 일어나지 않았다. 3T3-L1 지방세포 분화 중 SREBP1α와 LXR 유전자 발현은 MDI 처리에 의해 유의적 으로 증가하였다(P<0.05). SREBP1α 발현은 EPA(50, 100 μM)에 의해 유의적인 수준으로 감소하였고, LXR 발현은 고 농도의 EPA(100 μM) 처리에 의해서만 유의적으로 감소하 였다(Fig. 5, middle panel). 흥미롭게도 MDI 처리에 의해 유의적으로 증가한 AGPAT2 유전자 발현량은 EPA(50, 100 μM)에 의해 유의적으로 증가하였으며(P<0.001), GPAT4 의 유전자 발현량은 MDI 혹은 EPA 처리에 의해 변화하지 않았다(Fig. 5, lower panel).
체내 중성지질의 합성과 산화의 항상성 유지는 매우 중요 하며, 이의 불균형은 비만, 심장질환, 대사질환 등을 포함한 만성질환의 원인이 될 수 있다(Unger와 Orci, 2001). GPAT 과 AGPAT는 생체 내(de novo) 중성지질 합성의 각각 첫 번째와 두 번째 과정에 관여하는 중요한 효소들이다(Nagle 등, 2008). GPAT는 glycerol-3-phosphate의 아세틸화를
통해 lysophosphatidic acid(LPA)를 만들고, AGPAT는 LPA를 아세틸화하여 phosphatidic acid(PA; triacylglyc- erol(TAG)의 전구물질)를 생성해낸다. Phosphatidic acid phosphohydrolase에 의해 PA는 diacylglycerol(DAG)로, 이후 diacylglycerol acyltransferase의 작용에 의해 DAG 는 TAG로 아세틸화된다(Han 등, 2006; Donkor 등, 2007).
GPAT의 여러 isoform 중에 GPAT1과 GPAT2는 미토콘드 리아 membrane에 위치해 있고 GPAT3, GPAT4는 endo- plasmic reticulum에 위치하여 백색지방조직에서 중성지질 의 합성에 중요한 역할을 한다(Nagle 등, 2008).
AGAPT의 여러 isoform(1~8) 중, AGPAT1과 AGPAT2 가 AGAPT의 활성을 잘 나타내며(Aguado와 Campbell, 1998; Eberhardt 등, 1997) 나머지 3~5는 AGPAT의 활성 이 매우 낮다(Lu 등, 2005). 흥미롭게도 AGPAT2는 특히 지방조직에서 그 유전자가 많이 발현되고, AGPAT2의 mu- tation은 lipodystrophy의 원인이 되어 중성지질의 합성이 나 저장을 방해하여 인슐린 저항성이나 고중성지질 혈증 등 을 나타낼 수 있다고 보고된 바 있다(Agarwal 등, 2002).
3T3-L1 지방전구세포의 분화 중 AGPAT 유전자의 조절 (증가 혹은 감소)이 어떤 physiological한 변화를 가져오는 지에 대한 결론을 내리기는 쉽지 않다. Mardian 등(2017)의 연구에서 AGPAT4가 knock-out된 수컷 쥐에서 대조군 (WT) 쥐 대비 백색지방조직의 무게가 40%가량 증가해있음 을 보고하였다.
본 연구에서는 3T3-L1 지방전구세포의 분화 과정 중 DHA(50 μM)와 EPA(50, 100 μM)는 MDI 및 인슐린 처리에 의해 증가한 PPARα와 PPARγ 유전자의 발현량을 유의적으 로 증가시킨 반면 AGPAT2 유전자의 발현량을 증가시켰다.
AGPAT2 유전자 증가는 DHA에 의해서라기보다 EPA에 의 한 증가 효능이 더욱 유의적으로 보인다. GPAT4의 유전자 발현은 DHA나 EPA에 의해 모두 통계적 유의성은 없으나 EPA에 의해 감소하는 경향을 보였다(DHA에 의해서는 변 화 없음). EPA에 의한 GPAT4 감소는 통계적으로 유의하지 는 않으나, 이의 감소는 MDI로 분화시키지 않은 3T3-L1 지방세포 음성 대조군과 유사한 수준이다.
따라서 EPA는 각각 lipogenic 전사인자이자 지방산 및 콜레스테롤의 항상성 유지에 관여하는 SREBP1α, LXR 유 전자 발현을 유의적으로 감소시켰고, AGPAT2 및 GPAT4 의 유의적이지는 않지만 대조군과 유사한 수준으로 감소시 킴 등이 EPA의 지방분화 억제와 연관이 있을 것으로 사료된 다. 반면, DHA의 AGPAT2 증진 효능 등이 지방억제에 관여 하였을 가능성이 있다. AGPAT2 등이 mutation된 동물 모 델에서 오메가-3 지방산의 유전자 혹은 단백질 조절에 관한 추가적인 연구가 추후에 이루어져야 하겠다.
요 약
본 연구에서는 오메가-3 지방산인 DHA와 EPA의 3T3-L1
지방세포 분화 과정 중 지방축적 및 HAT 활성 억제능을 관찰하였으며, DHA는 AGPAT2, EPA는 SREBP1α, LXR, AGPAT2 및 GPAT4의 유전자 발현 조절을 통하여 지질축 적을 억제함을 밝혔다. HeLa 세포의 NE를 HAT의 source 로 하여 in vitro에서 HAT 활성 억제능을 평가한 결과 DHA 와 EPA 모두 유의적으로 HAT 활성을 억제하였다. DHA는 농도 의존적으로 HAT 활성을 억제한 반면, EPA는 저농도 처리군에서부터 유의적으로 HAT 활성을 억제하였다. DHA 와 EPA 저농도 처리군만 비교했을 때 EPA의 HAT 억제제 로서의 활성이 좀 더 큼을 알 수 있었다. 지방전구세포인 3T3-L1 세포에 MDI와 인슐린을 처리하여 지방분화를 유 도하였고, 동시에 DHA와 EPA를 처리한 결과 200 μM 농도 까지 세포 생존률을 억제시키지 않고 오히려 증가시켰다.
Oil Red O 염색을 통해 지방구를 염색하고 정량한 결과, DHA와 EPA 모두 3T3-L1 지방세포에서 지질축적을 유의 적으로 억제하였다. DHA와 EPA에 의한 지질축적 억제는 세포 활성 억제에 의한 것이 아님을 알 수 있었다. HAT는 lipogenic 유전자의 전사인자 발현에 관여하므로, HAT 억 제제는 비만 혹은 지방간의 촉진을 막을 수 있다. 이는 여러 식품소재 혹은 다양한 페놀계 화합물을 활용한 실험에서 입 증된 바 있다. DHA 및 EPA는 PPARα와 PPARγ 유전자 발 현을 유의적으로 증가시킨 반면, SREBP1α와 LXR은 DHA 에 의해 증가하였고 EPA에 의해서는 유의적으로 감소하였 다. 또한, AGAPT2의 유전자 발현은 EPA에 의해서만 유의 적으로 증가하였고 DHA에 의한 변화는 관찰되지 않았다.
중성지방 합성에 관여하는 GPAT4 또한 DHA나 EPA에 의 한 유의적 변화는 관찰되지 않았다. EPA에 의한 지방분화 억제는 SREBP1α, LXR 유전자 발현의 감소, AGPAT2 증가 및 GPAT4의 유의적이지는 않지만 대조군과 유사한 수준으 로의 감소 등을 통해 일어났다고 사료된다. DHA의 AGPAT2 증진 효능 등이 지방억제에 관여하였을 가능성이 있다. 이러 한 유전자 발현의 조절은 앞서 관찰된 DHA 및 EPA의 HAT 활성 억제와 관련이 있을 수 있다. 염색질 면역침전법 등을 활용한 입증 연구 및 동물 모델을 활용한 입증이 추후 필요 할 것이다. 식품소재 혹은 지표 성분들의 후성유전적 조절을 통한 질병예방에 관한 연구는 최근 활발하게 이루어지고 있 다. 비만 모델에서 오메가-3 지방산을 이용한 후성유전적 조절, 특히 히스톤아세틸전달효소의 활성 억제를 통한 유전 자 발현 조절을 밝힌 연구는 본 연구논문이 최초이다. 오메 가-3는 풍부한 식사 혹은 식이보충제로 많이 이용되는 피쉬 오일 등의 복용으로 HAT 활성을 억제시키고, 이를 통해 비 만 및 관련 질환의 예방에 도움이 될 수 있다는 가능성을 제시하고 있다.
감사의 글
본 연구는 한국식품연구원 사업연구비(과제번호 E0150301- 06)의 지원을 받아 수행한 연구 결과로 이에 감사드립니다.
REFERENCES
Agarwal AK, Arioglu E, de Almeida S, Akkoc N, Taylor SI, Bowcock AM, et al. AGPAT2 is mutated in congenital gener- alized lipodystrophy linked to chromosome 9q34. Nat Genet.
2002. 31:21-23.
Aguado B, Campbell RD. Characterization of a human lysophos- phatidic acid acyltransferase that is encoded by a gene located in the class Ⅲ region of the human major histocompatibility complex. J Biol Chem. 1998. 273:4096-4105.
Barber E, Sinclair AJ, Cameron-Smith D. Comparative actions of omega-3 fatty acids on in-vitro lipid droplet formation.
Prostaglandins Leukot Essent Fatty Acids. 2013. 89:359-366.
Choi HK, Shin EJ, Park SJ, Hur HJ, Park JH, Chung MY, et al. Ethanol extract of Capsella bursa-pastoris improves he- patic steatosis through inhibition of histone acetyltransferase activity. J Med Food. 2017. 20:251-257.
Choi KC, Jung MG, Lee YH, Yoon JC, Kwon SH, Kang HB, et al. Epigallocatechin-3-gallate, a histone acetyltransferase inhibitor, inhibits EBV-induced B lymphocyte transformation via suppression of RelA acetylation. Cancer Res. 2009. 69:
583-592.
Chung MY, Shin EJ, Choi HK, Kim SH, Sung MJ, Park JH, et al. Schisandra chinensis berry extract protects against stea- tosis by inhibiting histone acetylation in oleic acid-treated HepG2 cells and in the livers of diet-induced obese mice.
Nutr Res. 2017. 46:1-10.
Chung MY, Song JH, Lee J, Shin EJ, Park JH, Lee SH, et al.
Tannic acid, a novel histone acetyltransferase inhibitor, pre- vents non-alcoholic fatty liver disease both in vivo and in vitro model. Mol Metab. 2019. 19:34-48.
Connor WE. Importance of n-3 fatty acids in health and disease.
Am J Clin Nutr. 2000. 71:171S-175S.
de Moraes Maciel R, da Silva Dutra DL, Rumjanek FD, Juliano L, Juliano MA, Fantappié MR. Schistosoma mansoni histone acetyltransferase GCN5: linking histone acetylation to gene activation. Mol Biochem Parasitol. 2004. 133:131-135.
Donkor J, Sariahmetoglu M, Dewald J, Brindley DN, Reue K.
Three mammalian lipins act as phosphatidate phosphatases with distinct tissue expression patterns. J Biol Chem. 2007.
282:3450-3457.
Eberhardt C, Gray PW, Tjoelker LW. Human lysophosphatidic acid acyltransferase. cDNA cloning, expression, and local- ization to chromosome 9q34.3. J Biol Chem. 1997. 272:20299- 20305.
Flachs P, Rossmeisl M, Bryhn M, Kopecky J. Cellular and mo- lecular effects of n-3 polyunsaturated fatty acids on adipose tissue biology and metabolism. Clin Sci (Lond). 2009. 116:1- 16.
Gelman L, Zhou G, Fajas L, Raspé E, Fruchart JC, Auwerx J.
p300 interacts with the N- and C-terminal part of PPARγ2 in a ligand-independent and -dependent manner, respectively.
J Biol Chem. 1999. 274:7681-7688.
Giandomenico V, Simonsson M, Grönroos E, Ericsson J. Coacti- vator-dependent acetylation stabilizes members of the SREBP family of transcription factors. Mol Cell Biol. 2003. 23:2587- 2599.
Guasch-Ferre M, Babio N, Martinez-Gonzalez MA, Corella D, Ros E, Martin-Pelaez S, et al. Dietary fat intake and risk of cardiovascular disease and all-cause mortality in a population at high risk of cardiovascular disease. Am J Clin Nutr. 2015.
102:1563-1573.
Han GS, Wu WI, Carman GM. The Saccharomyces cerevisiae lipin homolog is a Mg2+-dependent phosphatidate phosphatase
enzyme. J Biol Chem. 2006. 281:9210-9218.
Hill JO, Melanson EL, Wyatt HT. Dietary fat intake and regu- lation of energy balance: implications for obesity. J Nutr.
2000. 130:284S-288S.
Hwang JT, Choi HK, Kim SH, Chung S, Hur HJ, Park JH, et al. Hypolipidemic activity of Quercus acutissima fruit ethanol extract is mediated by inhibition of acetylation. J Med Food.
2017. 20:542-549.
Imamura F, Micha R, Wu JHY, de Oliveira Otto MC, Otite FO, Abioye AI, et al. Effects of saturated fat, polyunsaturated fat, monounsaturated fat, and carbohydrate on glucose-insulin ho- meostasis: A systematic review and meta-analysis of rando- mised controlled feeding trials. PLoS Med. 2016. 13:e1002087.
https://doi.org/10.1371/journal.pmed.1002087
Kadowaki T, Yamauchi T. Adiponectin and adiponectin receptors.
Endocr Rev. 2005. 26:439-451.
Kim HJ, Choi HK, Chung MY, Park JH, Chung S, Lee SH, et al. Ethanol extract of Ligularia fischeri inhibits the lipopoly- saccharide-induced inflammatory response by exerting anti- histone acetyltransferase activity to negatively regulate p65.
J Med Food. 2019. 22:1127-1135.
Kim HK, Della-Fera M, Lin J, Baile CA. Docosahexaenoic acid inhibits adipocyte differentiation and induces apoptosis in 3T3-L1 preadipocytes. 2006. 136:2965-2969.
Kondo T, Asai M, Tsukita K, Kutoku Y, Ohsawa Y, Sunada Y, et al. Modeling Alzheimer’s disease with iPSCs reveals stress phenotypes associated with intracellular Aβ and differ- ential drug responsiveness. Cell Stem Cell. 2013. 4:487-496.
Lee MS, Kwun IS, Kim Y. Eicosapentaenoic acid increases lip- olysis through up-regulation of the lipolytic gene expression and down-regulation of the adipogenic gene expression in 3T3-L1 adipocytes. Genes Nutr. 2008. 2:327-330.
Lorente-Cebrian S, Costa AGV, Navas-Carretero S, Zabala M, Martinez JA, Moreno-Aliaga MJ. Role of omega-3 fatty acids in obesity, metabolic syndrome, and cardiovascular diseases:
a review of the evidence. J Physiol Biochem. 2013. 69:633- 651.
Lu B, Jiang YJ, Zhou YT, Xu FY, Hatch GM, Choy PC. Cloning and characterization of murine 1-acyl-sn-glycerol 3-phosphate acyltransferase and their regulation by PPARα in murine heart.
Biochem J. 2005. 385:469-477.
Manzo F, Tambaro FP, Mai A, Altucci L. Histone acetyltransfer- ase inhibitors and preclinical studies. Expert Opin Ther Pat.
2009. 19:761-774.
Mardian EB, Bradley RM, Aristizabal Henao JJ, Marvyn PM, Moes KA, Bombardier E, et al. Agpat4/Lpaatδ deficiency highlights the molecular heterogeneity of epididymal and peri- renal white adipose depots. J Lipid Res. 2017. 58:2037-2050.
Mozaffarian D, Wu JHY. Omega-3 fatty acids and cardio- vascular disease: effects on risk factors, molecular pathways, and clinical events. J Am Coll Cardiol. 2011. 58:2047-2067.
Murali G, Desouza CV, Clevenger ME, Ramalingam R, Sara- swathi V. Differential effects of eicosapentaenoic acid and do- cosahexaenoic acid in promoting the differentiation of 3T3- L1 preadipocytes. Prostaglandins Leukot Essent Fatty Acids.
2014. 90:13-21.
Nagle CA, Vergnes L, Dejong H, Wang S, Lewin TM, Reue K, et al. Identification of a novel sn-glycerol-3-phosphate acyltransferase isoform, GPAT4, as the enzyme deficient in Agpat6-/- mice. J Lipid Res. 2008. 49:823-831.
Neschen S, Morino K, Rossbacher JC, Pongratz RL, Cline GW, Sono S, et al. Fish oil regulates adiponectin secretion by a peroxisome proliferator-activated receptor-γ-dependent me- chanism in mice. Diabetes. 2006. 55:924-928.
Oster RT, Tishinsky JM, Yuan Z, Robinson LE. Docosahexae- noic acid increases cellular adiponectin mRNA and secreted adiponectin protein, as well as PPARγ mRNA, in 3T3-L1 adipocytes. Appl Physiol Nutr Metab. 2010. 35:783-789.
Pinel A, Pitois E, Rigaudiere JP, Jouve C, Saint-Vincent SD, Laillet B, et al. EPA prevents fat mass expansion and meta- bolic disturbances in mice fed with a western diet. J Lipid Res. 2016. 57:1382-1397.
Prostek A, Gajewska M, Bałasińska B. The influence of eicosa- pentaenoic acid and docosahexaenoic acid on expression of genes connected with metabolism and secretory functions of ageing 3T3-L1 adipocytes. Prostaglandins Other Lipid Media- tors. 2016. 125:48-56.
SanGiovanni JP, Chew EY. The role of omega-3 long-chain poly- unsaturated fatty acids in health and disease of the retina.
Prog Retin Eye Res. 2005. 24:87-138.
Schwab JM, Chiang N, Arita M, Serhan CN. Resolvin E1 and protectin D1 activate inflammation-resolution programs. Nature.
2007. 447:869-874.
Shan D, Li JL, Wu L, Li D, Hurov J, Tobin JF, et al. GPAT3 and GPAT4 are regulated by insulin-stimulated phosphoryl- ation and play distinct roles in adipogenesis. J Lipid Res.
2010. 51:1971-1981.
Soni N, Ross AB, Scheers N, Nookaew I, Gabrielsson BG, Sandberg AS. The omega-3 fatty acids EPA and DHA, as a part of a murine high-fat diet, reduced lipid accumulation in brown and white adipose tissues. Int J Mol Sci. 2019. 20:
5895. https://doi.org/10.3390/ijms20235895
Spiegelman BM, Flier JS. Obesity and the regulation of energy
balance. Cell. 2001. 104:531-543.
Tian Y, Wong VWS, Chan HLY, Cheng ASL. Epigenetic regu- lation of hepatocellular carcinoma in non-alcoholic fatty liver disease. Semin Cancer Biol. 2013. 23:471-482.
Unger RH, Orci L. Diseases of liporegulation: new perspective on obesity and related disorders. FASEB J. 2001. 15:312-321.
Visscher TLS, Seidell JC. The public health impact of obesity.
Annu Rev Public Health. 2001. 22:355-375.
Wainwright PE. Dietary essential fatty acids and brain function:
a developmental perspective on mechanisms. Proc Nutr Soc.
2002. 61:61-69.
Wang Y, Viscarra J, Kim SJ, Sul HS. Transcriptional regulation of hepatic lipogenesis. Nat Rev Mol Cell Biol. 2015. 16:678- 689.
Wapenaar H, Dekker FJ. Histone acetyltransferases: challenges in targeting bi-substrate enzymes. Clin Epigenet. 2016. 8:59.
https://doi.org/10.1186/s13148-016-0225-2
Yang J, Goldstein JL, Hammer RE, Moon YA, Brown MS, Hor- ton JD. Decreased lipid synthesis in livers of mice with dis- rupted site-1 protease gene. Proc Natl Acad Sci U S A. 2001.
98:13607-13612.
Zhu Y, Bo Y, Liu Y. Dietary total fat, fatty acids intake, and risk of cardiovascular disease: a dose-response meta-analysis of cohort studies. Lipids Health Dis. 2019. 18:91. https://doi.
org/10.1186/s12944-019-1035-2
Zhuang P, Lu Y, Shou Q, Mao L, He L, Wang J, et al. Differen- tial anti-adipogenic effects of eicosapentaenoic and docosa- hexaenoic acids in obesity. Mol Nutr Food Res. 2019. 63:
e1801135. https://doi.org/10.1002/mnfr.201801135