—160—
서 론
모래주사 Microphysogobio koreensis는 잉어과(Cyprinidae) 모래무지아과(Gobioninae)에 속하는 한국 고유종으로, 섬진 강과 낙동강 수계 등에 서식하는 것으로 알려져 있다(김과 박, 2002). 모래주사속 어류는 공통적으로 입이 아래쪽에 있 고 윗입술에는 유두돌기가 존재하여 돌에 붙어 있는 부착 조류를 섭식하는 특징을 가지고 있는데(김, 1997), 국내에 서식하는 모래주사속 어류는 모두 5종이 알려져 있으며, 그 중 모래주사는 Mori (1935)에 의해 처음 보고된 후 형태적 으로 유사한 돌마자 M. yaluensis와 혼돈되어 왔으나, 윗입
술의 유두상소돌기의 수, 구엽 및 복부 비늘의 유무를 기준 으로 돌마자와 구분한 바 있다(김, 1984).
모래주사는 근연종인 돌마자가 대부분의 하천에 다수 분 포하는 것과는 달리 서식처가 매우 제한적이고 그 개체수 또한 많지 않아 환경부에서는 2001년 멸종위기 야생동∙식 물 II급으로 지정하여 보호받고 있다. 모래주사에 관한 연 구는 모래주사속 어류의 재검토(Kim and Yang, 1999), 버들 매치속과 모래주사속 어류의 분자계통학적 연구(Kim and Bang, 2010) 등 일부 연구에서만 제한적으로 다루었을 뿐 발생 및 생태에 관련된 연구는 아직까지 보고된 바 없다.
어류의 초기생활사 연구는 종의 생물학적 특징을 이해하 는 데 가장 기초가 되는 연구 분야이며, 근연종 사이의 계 통분류학적인 위치 규명에 중요한 요인으로 작용한다(Blax- ter, 1974). 특히 서식처 및 개체수의 감소로 인해 멸종위기
멸종위기종 모래주사 Microphysogobio koreensis의 난발생 및 초기생활사
김치홍∙윤승운1∙김재구1∙김현태1∙박종성1∙박종영1,
*
국립수산과학원 중앙내수면연구소, 1전북대학교 자연과학대학 생명과학과∙생물다양성연구소
Embryonic Development and Early Life History of the Endangered Species Microphysogobio koreensis (Pisces: Cyprinidae)by Chi-Hong Kim, Seung-Woon Yoon1, Jae-Goo Kim1, Hyun-Tae Kim1, Jong-Sung Park1and Jong-Young Park1,*(Central Inland Fisheries Research Institute, Cheongpyeong-ri Cheongpyeong-myeon Gapyeong-gun, Gyeonggi-do 477-816, Korea; 1Department of Biological Science and Institute for Biodiversity Research, College of Natural Sci-ences, Chonbuk National University, Jeonju 561-756, Korea)
ABSTRACT Egg development and early life history of the endangered Korean freshwater fish, Micro- physogobio koreensis, was investigated to get a basic information for establishing its conservation strategy. The matured adult fishes were sampled at Imsil-gun, Jeollabuk-do, Korea in May 1 2012 and artificially fertilized. The fertilized eggs were spherical, separated demersal and 1.8±±0.14 mm in dia- meter. The hatching of the embryo began at about 29 hours after fertilization under water temperature of 23。C. The newly hatched pre-larvae were average 2.2±±0.48 mm in total length. At 4 days after hatching, the post-larvae were 3.6±±0.55 mm in total length and york sac was completely absorbed. At 20 days after hatching, their fin rays were formed and finally the larvae entered juvenile stage and grew up to 6.5±±0.77 mm in total length. At 60 days after hatching, the total length reached 24.4±±1.71 mm, and the band patterns of he head and lateral side were similar to adult fish. Based on this study, the fertilized eggs of M. koreensis hatched more quickly and the pre-larvae length was also smaller, compared with related species.
Key words : Endangered fish, Microphysogobio koreensis, egg development, early life history
*교신저자: 박종영 Tel: 82-63-270-3344, Fax: 82-63-270-3362, E-mail: [email protected]
http://www.fishkorea.or.kr
에 처해진 종의 경우 종 복원과 관련하여 초기생활사 연구 가 더욱 중요한 연구 분야로 대두되고 있는 실정이며 이에 따라 국내 멸종위기어류의 초기생활사 연구는 꾸구리(Ko et al., 2011), 돌상어(고 등, 2011), 가는돌고기(고 등, 2012), 미호종개(송 등, 2008), 얼룩새코미꾸리(송 등, 2009), 묵납 자루 (백과 송, 2005) 등에서 이루어진 바 있다.
따라서 본 연구에서는 아직까지 연구되지 않은 멸종위기 어류 모래주사의 난발생과정 및 초기생활사를 연구함으로 써 멸종위기종의 종 보존을 위한 인공증식기술 개발에 필 요한 기초자료로 활용하고자 실시되었다.
재료 및 방법
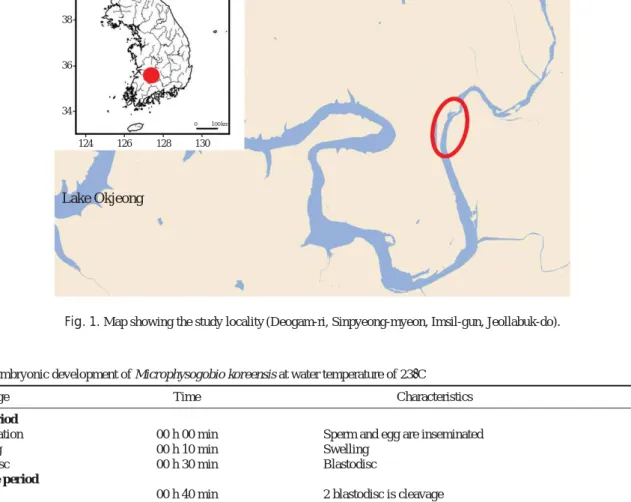
모래주사 친어의 포획은 전주지방환경청의 허가를 통해 이루어졌으며, 전라북도 임실군 신평면 덕암리 일대에서 2012년 4월 말부터 5월 초까지 2~3일 간격으로 투망(망 목 7×7 mm)을 이용하여 실시하였다(Fig. 1). 수정에 사용 된 모래주사는 2012년 5월 1일 포획된 성어를 이용하였으 며, 산란기 친어의 구분은, 현지에서 포획된 성체 가운데 복 부압박법으로 정액 또는 성숙난이 체외로 배출되는 친어를 선별하였다. 선별된 암, 수 각각 10개체를 휴대용 기포기로
Fig. 1. Map showing the study locality (Deogam-ri, Sinpyeong-myeon, Imsil-gun, Jeollabuk-do).
Table 1. Embryonic development of Microphysogobio koreensis at water temperature of 23�C
Stage Time Characteristics Fig. 2
Zygote period
Insemination 00 h 00 min Sperm and egg are inseminated
Swelling 00 h 10 min Swelling A
Blastodisc 00 h 30 min Blastodisc B
Cell cleave period
2 cells 00 h 40 min 2 blastodisc is cleavage C
4 cells 00 h 55 min 2~2 array of blastomeres D
8 cells 01 h 15 min 2~4 array of blastomeres E
16 cells 01 h 35 min 4~4 array of blastomeres F
32 cells 02 h 15 min 4~8 array of blastomeres G
Morula 03 h 10 min Cleavage plans irregular H
Blastula 07 h 00 min Elliptical shape I
Early gastrula 09 h 30 min Early gastrulation J
Gastrula 09 h 45 min Gastrulation K
Late gastrula 13 h 20 min Late gastrulation L
Segmentation period
4~5 mytomes 14 h 20 min 4~5 myotomes, formation of optic vesicles M
9~10 mytomes 21 h 00 min 9~10 myotomes N
20~25 myotomes 24 h 00 min 20~25 myotomes, formation of heart O
35~40 mytomes 29 h 00 min 35~40 mytomes, start hatching P
Lake Okjeong
124 126 128 130
0 100 km
38
36
34
지속적인 산소공급을 해주면서 실험실로 운반하였으며, 생 식소성숙도를 확인하기 위해 암, 수 각각 5개체에 대한 전 장, 체장, 체중, 생식소 무게를 측정하였으며, 생식소성숙도 지수(Gonadosomatic Index, GSI)는 다음과 같이 구하였다.
생식소성숙도지수(GSI)==생식소성숙도(g) / 체중(g)×102 실험실로 운반된 친어는 호르몬 처리 없이 암컷의 복부 를 압박하여 수정란을 채란한 후 수컷의 정액을 같은 방법 으로 채정하여 건식법으로 수정시켰다. 수정란은 직경 15 cm의 페트리디쉬에 분산시켜 사육하였으며, 부화 이후부터 는 소형 수조(40×20×25 cm), 치어기 이후에는 바닥에 모 래가 깔린 수조(60×40×30 cm)로 각각 옮겨 사육하였다.
부화 후 난황이 흡수된 자어에는 매일 아침 식물성 배합사 료(Hikari)를 공급하였으며 사육수는 수온과 광주기를 일정 하게 유지시키면서(23�C, 14D : 10N), 매일 저녁 1/2씩 환수 하였다.
수정란의 발생과정은 해부현미경(Carl Zeiss, Germany)과 디지털카메라(Nikon D300S, Japan)를 이용하여 촬영하였으 며, 자치어의 발달과정은 임의로 10개체를 선정하여 MS-222 (Tricaine methane sulfonate; Woojin B&G Co., Korea)로 마 취한 후 촬영 및 관찰하였다.
결 과
1. 친어 및 포란수
인공수정에 사용한 모래주사 친어의 크기는 암컷이 체중 (body weight) 12.1±3.40 g, 전장(total length) 106.8±9.50 mm (n==10), 수컷이 체중 9.9±0.87 g, 전장 102.5±2.84 mm (n==10)으로 나타나 암컷이 수컷에 비해 약간 큰 것으로 확 인되었다.
완전히 성숙한 모래주사 암컷의 포란수는 3331±935개로 확인되었다 (n==5, BW: 10.6±3.55 g, TL: 101.4±9.76 mm, GSI: 12.2±0.90%).
2. 난 발생 과정
모래주사의 수정란은 완전한 구형이며 약한 접착성을 띠 는 분리 침성란으로 확인되었다. 수정 전의 난경은 약 1.0±
0.04 mm (n==30)이었으나 수정 10분 후 수정란들은 수분을 흡수하여 팽창하기 시작하여 1.8±0.14 mm (n==30)까지 커 졌다(Fig. 2A). 수정 후 30분이 경과하자 세포질이 동물극 쪽으로 이동하여 배반을 형성하였으며(Fig. 2B), 40분 후에 는 배반에 난할이 일어나 2세포기(Fig. 2C), 55분 후에는 4 세포기에 이르렀다(Fig. 2D). 수정 후 1시간 15분이 경과하 면서 수정란은 8세포기를 형성하였고(Fig. 2E), 수정 1시간
35분 후에는 16세포기(Fig. 2G), 2시간 15분 후에는 32세포 기를 형성하였다(Fig. 2G). 난할이 계속 진행되면서 3시간 10분 후에는 수정란이 상실기에 진입하였고(Fig. 2H), 7시 간이 경과된 후에는 포배기에 도달하였다(Fig. 2I). 수정 후 9시간 30분이 경과하자 수정란의 배반이 동물극에서 식물 극쪽으로 확장되어 난황의 절반 가까이를 감싸면서 낭배기 에 이르렀으며(Fig. 2J, K), 13시간 20분 후에는 난황의 거 의 대부분을 감싸면서 원구가 폐쇄되어 후기 낭배기를 형 성하였다(Fig. 2L). 수정 후 14시간 20분 후부터 수정란은 기관형성기에 진입하였는데, 두부와 미부의 구분이 뚜렷해 지고 두부에 뚜렷한 안포가 위치하였으며 4~5개의 체절이 관찰되었다(Fig. 2M). 21시간이 지나자 체절의 수가 9~10 개로 증가하였고, 안포 뒤쪽으로 이포가 확인되었으며, 미병 부에는 Kupper’s vesicle이 나타났다(Fig. 2N). 수정 후 24시 간에는 두부와 난황 사이에서 심장이 관찰되었고 안포 바 로 뒤쪽으로 가슴지느러미가 형성되었다. 체절은 20~25개 이며 미병부는 난황과 분리되어 신장하였고 미병부에 존재 하던 Kupper’s vesicle의 흔적은 찾을 수 없었다(Fig. 2O). 수 정 후 29시간이 지나자 배체에서는 활발한 움직임이 확인 되었고, 체절의 수는 35~40개였다. 난황과 분리된 미병부에 는 막지느러미의 분화가 관찰되었으며 일부 개체는 꼬리지 느러미를 이용하여 난막을 뚫고 부화하기 시작했다(Fig. 2P).
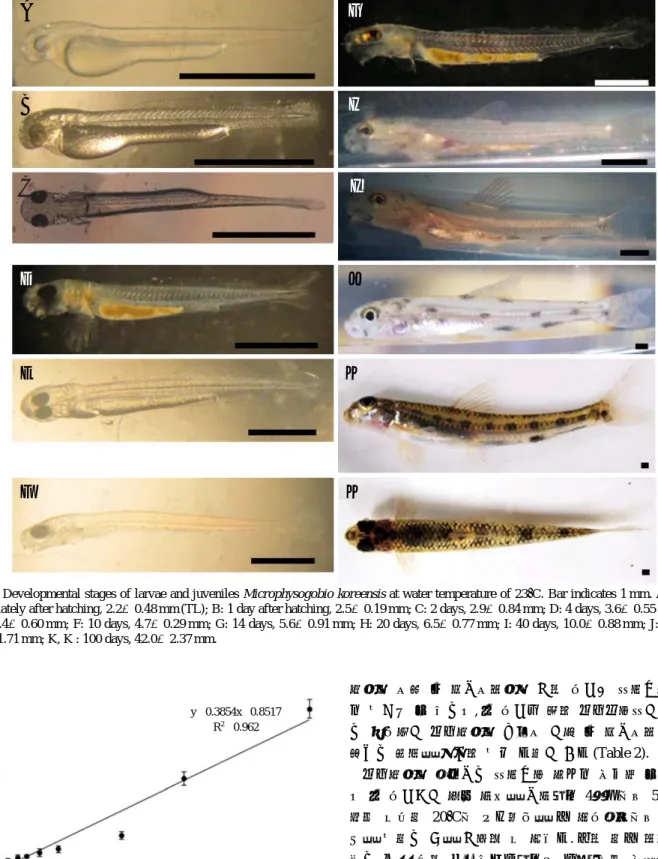
3. 자치어의 발달 과정
1) 전기 자어기
부화 직후 자어의 전장은 평균 2.2±0.48 mm (n==10)로 입 과 항문은 아직 열리지 않은 상태였고 몸은 불투명하였으며 머리 뒤쪽에서부터 시작되는 막지느러미를 가지고 있었다 (Fig. 3A). 부화 후 1일이 지나자 전장은 2.5±0.19 mm (n== 10)에 달하였고 난황이 부화 직후에 비해 1/3 정도 흡수된 상태였으며 안구에 색소가 약하게 침착되는 것을 관찰할 수 있었다(Fig. 3B). 입과 항문이 형성되기 시작하였고 가 슴지느러미가 발달하였다. 2일이 지난 자어의 전장은 2.9±
0.84 mm (n==10)였으며, 난황은 거의 흡수되었고 소화관이 형성되어 일부개체는 먹이활동을 시작하였다(Fig. 3C). 안 구는 검게 착색되었으며 가슴지느러미는 더욱 신장되었다.
2) 후기 자어기
부화 후 4일된 자어의 전장은 3.6±0.55 mm (n==10)로 난 황은 모두 흡수되었고 대부분의 개체가 활발한 먹이활동을 보여주었다(Fig. 3D). 가슴지느러미에는 8~10개의 원기가 형성되기 시작하였고 배면 아래쪽으로 검은색 색소포가 드 물게 관찰되었다. 7일이 지나자 전장은 4.4±0.60 mm (n==10) 에 달하였으며 난황은 완전히 흡수되었으며 배면의 색소포 는 점차 그 범위가 확장되어 가슴지느러미에서부터 꼬리지 느러미까지 이어졌다(Fig. 3E). 하나로 연결되었던 막지느러
미는 미병부가 함입되면서 분리되기 시작하였고, 꼬리지느 러미에서는 등 쪽으로 굴절된 미부봉상골이 관찰되었다. 또 한 두장이 점차 신장되어 앞쪽으로 돌출되었고 두부 정중 앙에서도 색소포가 확인되었다. 부화 후 10일이 경과한 모 래주사의 전장은 4.7±0.29 mm (n==10)였으며 가슴지느러미 원기는 점차 뚜렷해졌지만 기조막은 형성되지 않았다 (Fig.
3F). 등지느러미의 원기가 관찰되었으며 미병부의 막지느러 미는 더욱 함입되어 꼬리지느러미가 뚜렷해졌다. 부화 후 14일이 지난 자어의 전장은 5.6±0.91 mm (n==10)로 뒷지느 러미의 원기가 관찰되었으며 동공주변으로 홍채가 자리 잡 아 성체와 유사한 모습을 보였다(Fig. 3G).
3) 치어기
모래주사 자어는 부화 후 20일이 경과하자 치어기에 진 입하였는데, 전장은 6.5±0.77 mm (n==10)로 성장하였고 등 지느러미와 뒷지느러미를 비롯해 배지느러미의 기조가 분 화하였으며, 꼬리지느러미는 위아래가 분명하게 갈라지고 등
지느러미 기점부터 꼬리지느러미에 이르는 네 개의 검은 반 점이 뚜렷해졌다(Fig. 3H). 40일이 지난 치어의 전장은 10.0
±0.88 mm (n==10)였으며 등 면과 두부에 나타나는 색소포 의 크기가 더욱 크고 뚜렷해졌다(Fig. 3I). 배면에 산재하던 색소포들은 점차 반문을 형성하였으며, 주둥이가 돌출되기 시작하였고 꼬리지느러미에 반점이 희미하게 나타났다. 부 화 후 60일이 지나자 치어의 전장은 24.4±1.71 mm (n==10) 로 성장하였고 체측과 지느러미에 나타나는 반문이 성어와 동일해졌다(Fig. 3J). 100일이 지난 모래주사 치어는 전장이 42.0±2.37 mm (n==10)에 달하였다(Fig. 3K, K′, Fig. 4).
고 찰
모래주사는 섬진강과 낙동강 수계에 분포하나 그 서식처 가 제한적이고 개체수 또한 적어 환경부지정 멸종위기 야 생동∙식물 II급으로 분류되어 있는 어류이다. 때문에 모래 Fig. 2. Egg development stages of Microphysogobio koreensis at water temperature of 23�C. Time required for each embryonic stage is shown in Table 1.
A B C D E
F G H I J
K L M N
O P
1 mm
주사의 생태 및 초기생활사에 관한 연구가 아직까지 제대 로 보고된 바 없는데, 본 연구를 통해 모래무지아과에 속하 는 유연종과 모래주사의 난발생 과정 및 초기생활사에 나 타나는 특징을 비교해 보면 다음과 같다(Table 2).
모래주사의 산란기는 아직까지 제대로 알려진 바가 없는 데 본 연구결과 전북 임실을 기준으로 4월말에서 5월초 하 천의 수온이 20�C에 도달하였을 때 자연상태에서 산란행 동을 보이는 것을 관찰할 수 있었다. 또한 이때 포획된 친 어는 매우 약한 복부압박만으로도 많은 양의 알을 체외로 배출시켰으며, 수컷도 정자를 방출하였다. 한편 모래주사의 산란은 5월 초반에 지속적으로 관찰되다가 중순에는 전혀 확인되지 않았으며 이때의 수온은 23�C였다. 따라서 모래 주사의 산란기는 하천의 수온이 20�C에 도달하는 4월 말 부터 5월 초까지로 판단된다. 이는 같은 모래주사속 어류인 Fig. 3. Developmental stages of larvae and juveniles Microphysogobio koreensis at water temperature of 23�C. Bar indicates 1 mm. A: 0 day, immediately after hatching, 2.2±0.48 mm (TL); B: 1 day after hatching, 2.5±0.19 mm; C: 2 days, 2.9±0.84 mm; D: 4 days, 3.6±0.55 mm; E: 7 days, 4.4±0.60 mm; F: 10 days, 4.7±0.29 mm; G: 14 days, 5.6±0.91 mm; H: 20 days, 6.5±0.77 mm; I: 40 days, 10.0±0.88 mm; J: 60 days, 24.4±1.71 mm; K, K′: 100 days, 42.0±2.37 mm.
A G
B H
D J
E K
F K
′′C I
Fig. 4. Early growth of total length of Microphysogobio koreensis. The diagrams indicate the mean (circle) and standard deviation (horizontal line).
Total length(mm)
45 40 35 30 25 20 15 10 5 0
0 20 40 60 80 100
Days after hatching
y==0.3854x++0.8517 R2==0.962
돌마자의 5월 중순부터 7월까지에 비해서는 빨랐다(Baek, 1978).
모래주사의 수정란은 점착성의 침성란이었으며 꾸구리 Gobiobotia macrocephala (Ko et al., 2011), 돌마자 M. yaluen- sis (Baek, 1978), 모래무지 Pseudogobio esocinus (Lee et al., 2008)와 유사하였는데 이는 수정 후 가라앉은 수정란이 하 상의 돌 틈에 부착된 상태로 부화하는 어류의 일반적인 특 징으로 보였다. 반면 꺽지 Coreoperca herzi의 산란장에 탁 란하는 감돌고기 Pseudopungtungia nigra (Lee et al., 2004)나 조개껍데기에 수정란을 부착하는 참붕어 Pseudorasbora parva (Han et al., 2001)의 경우 강한 점착성을 띠어 모래주 사와 차이를 나타냈다.
모래주사 수정란의 크기는 1.8±0.14 mm로 모래주사속 어류인 돌마자(Baek, 1978)의 2 mm, 다른 모래무지아과 어 류인 모래무지(Lee et al., 2008)의 1.79~2.18 mm, 돌고기 Puntungia herzi (Lee et al., 2001)의 2.1±0.25 mm보다는 약 간 작았으며, 감돌고기(Lee et al., 2004)의 2.0~3.1 mm보다 는 작았고 꾸구리(Ko et al., 2011a)의 1.08±0.05 mm보다 는 컸다.
완전히 성숙한 모래주사 암컷의 포란수는 2041~4404 (3331±935)개로 돌마자(Baek, 1978)의 276~1230개, 감돌 고기(Lee et al., 2004)의 1400~1900개, 참붕어(Han et al., 2001)의 250~420개, 꾸구리(Ko et al., 2011)의 205~3361 개에 비해 상당히 많은 것으로 나타났다.
모래주사의 수정란은 23�C의 수온에서 29시간 후에 부 화하였는데 모래무지(Lee et al., 2008), 감돌고기(Lee et al., 2004), 돌고기 (Lee et al., 2001), 참붕어 (Han, 2001), 꾸구 리(Ko et al., 2011)보다는 빨랐지만, 돌마자(Baek, 1978)에 비해서는 느렸다. 수정란의 부화 시간은 수온과 밀접하게 관련되는데(Hokanson et al., 1973; Rana, 1990; Yoo et al., 1991; Yoon et al., 2007), 모래주사의 경우 자연산란장의 수 온이 20~23�C 정도인 부분을 고려하였을 때 자연상태 수 정란의 부화시간도 비슷할 것으로 보인다.
부화 직후 자어의 크기는 2.2±0.48 mm로 돌마자(Baek, 1978)의 0.7 mm를 제외한 나머지 모래무지아과 어류인 모
래무지(Lee et al., 2008), 감돌고기(Lee et al., 2004), 돌고기 (Lee et al., 2001), 참붕어(Han et al., 2001), 꾸구리(Ko et al., 2011)에 비해 작았는데 수정란의 크기에서 큰 차이를 보이 지 않았음에도 부화 직후 자어의 크기가 서로 다른 것은 수정란이 부화하는 데 소요되는 시간과 어느 정도 관련 있 을 것으로 판단된다.
어류에 있어서 색소포의 형성은 부화 시간이 짧은 종의 경우 부화 후에 관찰되는 반면 부화 시간이 긴 종의 경우 난 발생 과정 중에 관찰된다. 부화 시간이 20시간인 돌마 자(Baek, 1978)와 24시간인 미호종개 Cobitis choii (송 등, 2008)의 경우 부화 후 안구의 색소포가 침착되기 시작하였 으나 부화하는 데 100시간 이상 소요되는 모래무지(Lee et al., 2008), 감돌고기(Lee et al., 2004), 돌고기(Lee et al., 2001), 참붕어(Han et al., 2001), 꾸구리(Ko et al., 2011)의 경우 부 화 전에 안구뿐 아니라 몸통 및 꼬리에 색소포가 산재되는 것으로 보고되었다. 모래주사 자치어의 발달단계 중 색소포 는 부화 후 1일에 안구에서 가장 먼저 관찰되었으며 복부, 두부 순으로 형성되어 부화 시간이 짧은 돌마자 (Baek, 1978), 미호종개(송 등, 2008)와 유사하였다. 한편 일부 부 화가 지연된 수정란에서 부화 전에 이미 안구의 색소포가 침착된 것을 관찰할 수 있었다.
모래주사 자어가 치어기에 진입되는 시기는 부화 후 20 일이 경과했을 때였으며 이는 꾸구리(Ko et al., 2011)보다 는 빨랐으나, 돌마자(Baek, 1978), 모래무지(Lee et al., 2008), 감돌고기(Lee et al., 2004), 돌고기(Lee et al., 2001), 참붕어 (Han et al., 2001)보다는 느린 편이었다. 이는 종에 따른 차 이점으로 보이며, 동일종의 경우라도 사육환경에 따라 달라 질 수 있는 부분이므로 추가적인 연구를 통해 검토할 부분 으로 사료된다.
요 약
멸종위기 담수어인 모래주사의 종 보존을 위한 기초자료 를 얻기 위해 난발생 및 초기생활사를 연구하였다. 2012년 Table 2. Comparison of egg and larva developmental characteristics in Gobininae fishes
Species Fertilized
Fecundity Time of hatching Hatching Days until Days until
Reference
egg size (water temp.) size postlarva juvenile
Micropysogobio koreensis 1.8±0.14 2041~4404 29 h (23�C) 2.2±0.48 4 20 Present study (3331±935)
Micropysogobio yaluensis 2 276~1230 20 h (22~25�C) 0.7 11 31 Baek, 1978
Pseudogobio esocinus 1.79~2.18 - 168 h (19�C) 3.78~5.54 8 42 Lee et al., 2008
Pseudopungtungia nigra 2.0~3.1 1400~1900 189 h (19�C) 5.70~5.91 6 44 Lee et al., 2004
Puntungia herzi 2.1±0.25 - 143 h
5.65~6.15 15 - Lee et al., 2001
(1.95~2.25) (23.5~25.5�C)
Pseudorasbora parva 250~420 184 h (17.6�C) 4.5~5.5 3 65 Han et al., 2001
Gobiobotia macrocephala 1.08±0.05 205~3361 107 h (23�C) 4.4~4.8 4 15 Ko et al., 2011
5월 1일 전북 임실에서 포획된 성어를 이용하여 인공 수정 시켰으며, 수정란은 구형의 분리침성란에 난경은 1.8±0.14
mm였다. 23�C의 수온에서 29시간 후에 부화하였으며, 부화
직후 전기자어의 크기는 2.2±0.48 mm였다. 부화 후 4일째 난황을 모두 흡수하여 후기자어기에 진입하였으며, 전장은 3.6±0.55 mm였다. 20일이 지나자 지느러미 기조수가 정수 가 되어 치어기로 이행하였으며, 전장은 6.5±0.77 mm였다.
부화 후 60일이 지나자 두부와 체측에 나타나는 반문이 성 어와 유사해졌으며, 전장은 24.4±1.71 mm에 달하였다. 모 래주사의 난발생 과정 및 초기생활사를 모래무지아과 어류 들과 비교한 결과 수정란의 부화시간은 비교적 빨랐으며 부 화 직후 전기자어의 크기도 작았다.
사 사
본 연구는 국립수산과학원의 경상과제(과제번호 RP-2012- AQ-091)에 의해 수행되었음을 밝힙니다.
인 용 문 헌
고명훈∙박상용∙이일로∙방인철. 2011. 멸종위기 어류 돌상어의 난발생 및 초기생활사. 한국하천호수학회지, 44: 136-143.
고명훈∙박상용∙방인철. 2012. 가는돌고기의 난발생 및 초기생 활사. 한국어류학회지, 24: 48-55.
김익수. 1984. 한국산 모래무지아과 어류의 계통분류학적 연구.
한국수산학회지, 17: 436-448.
김익수. 1997. 한국동식물도감, 제37권 동물편(담수어류). 교육부, 629pp.
김익수∙박종영. 2002. 한국의 민물고기. 교학사, 465pp.
백현민∙송호복. 2005. 묵납자루의 난 형태와 초기생활사. 한국생 태학회지, 28: 281-286.
송하윤∙김우중∙이완옥∙방인철. 2008. 미호종개의 난 발생 및 자어 형태 발달. 한국하천호수학회지, 41: 104-110.
송하윤∙양 현∙조은미∙신현철∙방인철. 2009. 멸종위기에 처 한 얼룩새코미꾸리의 난 발생 및 자치어 형태 발달. 한국 어류학회지, 21: 247-252.
Baek, Y.G. 1978. On the life history of Microphysogobio yaluensis.
Korean J. Limnol., 11: 43-49.
Blexter, J.H.S. 1974. The early life history of fish. Spring-Verlag, Berlin, 765pp.
Han, K.H., S.H. Lee, W.I. Seo, D.J. Yoo, D.S. Jin, S.H. Oh and C.C.
Kim. 2001. The spawning behavior and early life history of the false dace, Pseudorasbora parva. J. Inst. Sci. Yosu Natl.
Univ., 3: 67-76. (in Korean)
Hokanson, K.E.F., J.H. McCormick and B.R. Jones. 1973. Tempera- ture requirements for embryos and larvae of the northern pike, Esox lucius (Linnaeus). Trans. Am. Fish. Sic., 102:
89-100.
Kim, I.S. and H. Yang. 1999. A revision of the genus Microphyso- gobio in Korea with description of a new species. Korean J.
Ichthyol., 11: 1-11.
Kim, K.Y. and I.C. Bang. 2010. Molecular phylogenetic position of Abbotina springeri based on nucleotide sequences of RAG1 gene. Korean J. Ichthyol., 22: 273-278.
Ko, M.H., W.J. Kim, S.Y. Park and I.C. Bang. 2011. Egg develop- ment and early life history of the endangered species Gobio- botia macrocephala (Cyprinidae). Korean J. Ichthyol, 23:
198-205. (in Korean)
Lee, S.H., G.N. Oh, K.S. Kim, Y.S. Oh, K.W. Kang, J.H. Hwang, B.I. Lee, W.K. Lee and K.H. Han. 2008. Embryonic and larval development of goby minnow, Pseudogobio esocinus.
Dev. Reprod., 12: 293-288. (in Korean)
Lee, S.H., K.H. Han, S.M. Yoon, D.S. Hwang, D.J. Yoo, C.L. Lee, I.S. Kim and Y.M. Son. 2004. Early life history and spawn- ing behavior of Pseudopuntungia nigra. Korean J. Ichthyol., 16: 309-316. (in Korean)
Lee, W.O., D.H. Kim, J.Y. Lee and E.O. Kim. 2001. Early develop- ment of Puntungia herzi (Pisces, Cyprinidae). Bulletin of Fisheries Research and Development Agency, 59: 61-67.
(in Korean)
Mori, T. 1935. Description of two new genera and seven species of cyprinidae from Korea. Ann. Zool. Japan, 15: 161-166.
Rana, K.G. 1990. Influence of incubation temperature on Oreochro- mis niloticus (L.) eggs and fry: I. Gross embryology, tempera- ture tolerance and rates of embryonic development. Aquacul- ture, 87: 165-181.
Yoon, S.J., D.H. Kim, H.G. Hwang, G.C. Song and Y.C. Kim. 2007.
Effects of water temperature, stocking density and feeding frequency on survival and growth in the oblong rockfish Sebastes oblongus lavae. Korean J. Ichthyol., 19: 1-7. (in Korean)
Yoo, S.K., Y.J. Chang and K.H. Kang. 1991. Influence of water tem- perature on egg development of the red sea bream, Pagrus major. J Aquaculture, 4: 13-18.