Volume 13, Number 1, June, 2010
※ 통신저자: 이 상 수
강원도 춘천시 교동 153
한림대학교 춘천성심병원 정형외과-골격노화연구소
TEL: 033) 240-5198 FAX: 033) 252-9875 E-mail: [email protected] 접수일: 2010년 6월 7일, 게재확정일: 2010년 6월 10일
*이 논문은 2010년도 정부(교육과학기술부)의 재원으로 한국연구재단의 지원을 받아 수행된 기초연구사업(R13- 2005-022-01000-0)과 2009년도 정부(교육과학기술부)의 재원으로 한국연구재단의 대학중점연구소 지원사업으 로 수행된 연구임(2009-0094072).
타이타늄 입자 자극에 의한 조골세포에서의 Wnt/β-catenin 신호계의 영향
한림대학교 의과대학 골격노화연구소∙정형외과학교실1), 천연의약연구소2), 기초의과학연구센터3)
남주석1,3)∙Sinha Nidi1,3)∙Ashish R Sharma1,3)∙이진구1,2)∙권순창1)∙장준동1)∙이상수1,3)
= Abstract =
Affection of Wnt/β-catenin in Titanium Particles Challenged Osteoblasts
Ju-Suk Nam, Ph.D.1,3), Sinha Nidi, B.S.1,3), Ashish R Sharma, M.S.1,3), Jin-Koo Lee, Ph.D.1,2), Sun-Chang Kwon, M.D.1), Jun-Dong Chang, M.D.1), Sang-Soo Lee, M.D.1,3)
Institute for Skeletal Aging & Orthopedic Surgery1), Institute for Natural Medicine2),
Infectious Disease Medical Research Center, Hallym University College of Medicin, Chuncheon, Korea3)
Purpose: The intracellular mechanisms that lead to periprosthetic osteolysis including impaired bone form- ing activity of osteoblast remain incompletely characterized. To determine the possibility that Ti-particles play a role to regulate Wnt/β-catenin signaling pathway in impaired osteogenesis, we analyzed the stability of β- catenin and the transcriptional changes of regulators for Wnt/β-catenin signaling pathway in MC3T3-E1 osteoblast cells.
Materials and Methods: Ti-particles were prepared by sterilizing and counted on the microscopy. Tran- scriptional changes of OPG, RANKL, LRP5, LRP6, DKK1 and sFRP2 were determined by real-time RTPCR.
Protein level of β-catenin and GSK3βwas detected using Western blotting and immunofluorescence staining.
Results: After 4 hours of treatment of Ti-particles, OPG/RANKL mRNA ratio was significantly decreased.
And also, decreased protein levels of β-catenin and phospho-GSK3β were detected. Using immunofluores- cence stain, it was confirmed that Ti-particles suppressed nucleus staining of β-catenin induced by Wnt3a con- ditioned medium. The results of real-time RT-PCR showed reduced level of LRP5 and LRP6 transcripts, and induced level of DKK1 and sFRP2 transcripts by challenging of Ti-particles
Conclusion: Our report suggests that Ti-particles may play a crucial role in the regulation of Wnt/β-catenin signaling pathway in osteoblast through the transcriptional changes of membrane receptors and extracellular inhibitors for Wnt.
Key Words: Wnt/ β-catenin, Osteoblast, Titanium particle
서 론
마모편(wear debris)이 골용해(periprosthet- ic osteolysis)의 발생과 진행의 기전에는 마모편 의 다양한 표적 세포들의 영향과 세포-세포간의 상호 영향에 대한 정확한 이해가 필요하다. 특정 마모편은 대식세포는 물론 인근 조직의 조골세포 등의 세포를 자극 할 수 있다7,16,18). 현재까지의 보 고에 의하면 마모편은 조골세포에 직접적인 작용 으로 조골세포에서의 골기질 생성능의 저하를 유 발할 수 있다고 알려져 있다4,15,19). 그러나 이러한 골용해 모델에서의 조골세포의 골형성능 저하 기 전은 아직까지 명확하지 않아 많은 관심이 필요한 상태이다.
당단백질로 구성된 Wnt family는 mes- enchymal progenitor들이 adipocytes와 chondrocytes로의 분화를 억제하면서 골형성을 일으키는 조골세포로의 분화를 촉진시키는 역할을 하여 골형성 및 재형성을 조절한다11,20). 최근에는 조골세포의 Wnt 신호계가 조골세포의 RANKL 의 생성을 억제하거나 Osteoprotegrin (OPG)의 증가를 통하여 파골세포의 활성화를 조절하는 것 으로 보고되고 있다13).
이에 본 연구에서는 골용해 상황에서 Wnt 신 호전달계가 조골세포의 골형성능 저하 기전으로 관여할 가능성을 규명하고자 조골세포에서의 titanium (Ti) 입자에 의한 Wnt 신호전달계의 영향을 분석하였다.
재료 및 방법 1. 세포 배양 및 Ti 입자의 준비
조골세포로 분화할 수 있는 MC3T3-E1 세포주 는 10% 우태아 혈청(FBS; Gibco, Grand Island, New York, USA), 페니실린 및 스트렙 토마이신이 포함된 αMEM (Gibco, Grand Island, New York, USA) 배지를 사용하여 5%
CO2와 37℃ 온도 하에서 배양되었다.
멸균된 Ti입자(Johnson Matthey, catalog
#00681; Alfa-Aesar, a Johnson Matthey Company, Ward Hill, Massachusetts, USA)
를 70% 에탄올에 48시간 이상 담궈 둔 다음, 이 를 다시 PBS로 씻어준 뒤 현미경하에서 5~10 μ m 크기의 입자를 선택하여 농도를 계산한다. 배양 된 세포에 세포당 입자 수를 1:40, 1:80, 1:160의 농도로 7시간 동안 처리하였다17).
2. Real-Time RT-PCR
Ti 입자를 처리한 MC3T3-E1 세포에서 Trizol 을 사용하여 전체 RNA를 추출한 후, 2 μg의 RNA와 Oligo d(T)12-18 및 역전사효소 (Pro- mega, Madison, WI, USA)로 cDNA를 합성 하였다. PCR 반응은 2 μl의 cDNA와 SYBR Green Mix (QIAGEN, Valencia, CA, USA) 를 사용하여 95℃ 15초, 60℃ 15초, 72℃ 20초의 조건으로 수행하였다.
파골세포 형성에 관여하는 OPG, RANKL 및 Wnt 신호전달에 관여하는 LRP5, LRP6, DKK1, sFRP2에 대한 primer를 제작하였다 (Table 1). 정량적 결과 분석을 위하여 GAPDH 의 발현 정도를 가지고 표준화 하였다.
3. Western blot analysis
Ti 입자를 처리한 MC3T3-E1 세포를 PBS로 세척하고 protease inhibitor (Sigma, St.
Louis, MO, U.S.A.)가 포함된 passive lysis buffer (Promega, Madison, WI, U.S.A)를 사용하여 단백질을 추출하였다. 단백질 추출액을 SDS-PAGE를 이용하여 분리한 후, western blotter (Bio-RAD)를 이용하여 PVDF mem- brane (Amersham Pharmacia, Piscat- away, NJ, U.S.A.)으로 옮겼다. Membrane 을 5%의 무지방 우유가 함유된 1X TBST (100 mM Tris (pH7.5), 1.5 M NaCl, 0.5%
tween-20) 용액으로 blocking하고, 1X TBST 로 세척한 후, 일차 항체를 1% 무지방 우유를 포함한 용액에서 부착시켰다. 일차항체 반응 후, membrane을 1X TBST를 이용하여 세척하고, 이차항체를 반응시킨 후, 1X TBST로 세척하고 ECL western blotting detection kit (Amersham Pharmacia)을 사용하여 발광을
유도한 후, X-ray film (Amersham Phar- macia)을 사용하여 현상하였다. 사용된 일차항 체는 anti-β-catenin antibody, anti-phos- pho-GSK3β antibody, anti-GSK3 anti- body, anti-Actin antibody (Santa Cruz, Santa Cruz, CA, U.S.A.)이며, 각 단백질의 발현 정도는 Actin 발현 정도로 표준화하였다.
4. 면역형광염색
β-catenin의 세포내 이동 및 발현량을 알아보 기 위하여 coverslip에 배양된 MC3T3-E1 세포 에 Ti 입자를 처리한 후, 4% paraformalde-
hyde로 10분간 고정시킨다. 고정된 세포를 10%
normal goat serum으로 blocking하고 anti-β- catenin antibody로 반응시킨다. 1X PBS로 세 척 후, FITC가 부착된 이차항체와 반응시킨 다 음, DAPI가 포함된 mounting 용액을 사용하여 slide위에 mounting 하였다.
결 과
1. 골용해의 지표로서의 OPG/RANKL 비율
Ti 입자들이 조골세포에 미치는 직접적인 영향 을 알아보기 위하여, Ti 입자들을 처리한 MC-
Table 1. Primers for real-time RT-PCR
Gene Forward primer (5’→3’) Reverse primer (5’→3’) Product size GenBank No.
OPG CCACAATGAACAAGTGGCTGTGCT TAGGTAGGTGCCAGGACACATTT 155 NM_008764.3 RANKL CGTGCAGAAGGAACTGCAACACAT TTGATGGTGAGGTGTGCAAATGGC 135 NM_011613.3
LRP5 CCTGGAGCTGTTGAGTGACA GAGTGGGATAGCCACATCGT 124 NM_008513
LRP6 GGACGATCAGTTGGAGTGGT CAGCCCGTTCAATTTTAGGA 127 NM_008514
DKK1 CTGAAGATGAGGAGTGCGGCTC GGCTGTGGTCAGAGGGCATG 183 NM_010051.2 sFRP2 ATCCTGGAGACAAAGAGCAAGACC TGACCAGATACGGAGCGTTGATG 142 NM_009144.1 GAPDH TCAACAGCAACTCCCACTCTTCCA ACCCTGTTGCTGTAGCCGTATTCA 115 NM_008084.2
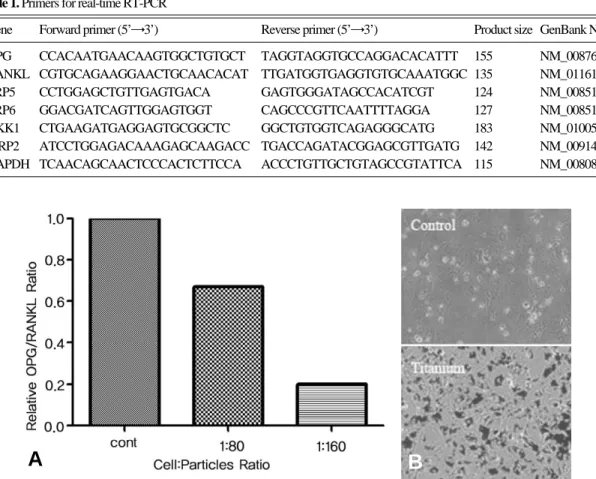
Fig. 1. Decreased OPG/RANKL ratio after treatment of Ti-particles in osteoblasts. (A) MC3T3-E1 cells were grown overnight in 6-well plates (2×105cells/well). Ti-particles were treated with indicated ratios for 3 hours. Total RNA was purified for real-time RTPCR. The ratio of RANKL/OPG was calculated with the gene expression level of RANKL and OPG normalized with GAPDH. (B) Distribution of Ti-particles in MC3T3-E1 cells visualized by phase contrast microscopy. Black dots indicate Ti-particles.
A B
3T3-E1 세포주에서 OPG와 RANKL에 대한 mRNA의 발현 변화를 real-time RTPCR 방법 을 이용하여 측정하였다. 4시간 처리한 실험군에 서의 발현양으로 OPG/RANKL 비율을 계산한 결과, 세포당 80개의 입자를 처리한 세포에서는 대조군과 비교하여 30%의 감소를 보였으며, 세 포당 160개의 입자를 처리한 세포에서는 80%의 감소를 나타냈다(Fig. 1).
2. Wnt 신호전달 경로를 조절하는 β-catenin 단백질의 변화
OPG/RANKL의 비율을 유지하면서 조골세포 에 의한 파골세포의 활성 정도를 조절하는 기작으 로 Wnt/β-catenin 신호전달 경로가 잘 알려져 있다14). 따라서, Ti 입자에 의한 Wnt/β-catenin 신호전달 경로의 변화를 알아보고자 Wnt/β- catenin 신호전달 경로의 주요 조절인자인 β- catenin과 β-catenin의 분해에 관여하는 GSK3β 에 대한 변화를 확인하였다. β-catenin의 전체 단백질 양은 Ti 입자에 의해 현저하게 감소됨을 나타냈으며, GSK3β의 활성을 저해하는 인산화도 감소됨을 Western blotting으로 보여주었다 (Fig. 2A). 또한, Wnt3a를 포함한 조건화 배지 와 Ti 입자를 동시에 처리 하였을 때, Wnt3a로
증가된 β-catenin의 핵내 이동을 감소시킴을 면 역형광염색을 이용하여 확인하였다(Fig. 2B).
3. Wnt 수용체인 LRP5/6 유전자 발현
β-catenin의 단백질 양과 핵내 이동을 조절 할 수 있는 인자들 중에 조골세포에서 골형성에 관여 하는 것으로 잘 알려진 Wnt 수용체인 LRP5와 LRP6가 존재한다13). MC3T3-E1 세포주에서 Ti 입자에 의한 LRP5와 LRP6의 발현양은 두 수용 체 모두 유사하게 80% 이상 감소됨을 real-time RTPCR로 확인 하였으며, 1:80과 1:160의 Ti 입자 농도를 처리한 세포에서 두 수용체 모두 감 소량의 차이는 보이지 않았다(Fig. 3).
4. Wnt 저해인자인 DKK1 및 sFRP2 유전자 발 현
Wnt/β-catenin 신호전달 경로의 활성은 다양 한 방법으로 조절 될 수 있는데, 조골세포에서 대 표적으로 Wnt 수용체인 LRP와 결합하여 Wnt 리간드의 신호전달을 저해하는 DKK1, Wnt 리 간드와 직접 결합하여 수용체를 통한 신호전달을 저해하는 sFRP2등이 알려져 있다3,10). real-time RTPCR을 통한 DKK1과 sFRP2의 발현 양상은
Fig. 2. Reduced β-catenin level after treatment of Ti-particles in osteoblasts. (A) MC3T3-E1 cells were grown overnight in 6-well plates (2×105cells/well). Ti-particles were treated with indi- cated ratios for 3 hours. Total cellular proteins were immunoblotted with anti-β-catenin anti- body, anti-phospho-GSK3βantibody, anti-GSK3 antibody, and anti-Actin antibody. (B) For immunofluorescence staining, cells were treated with L-cell conditioned medium (50%), Wnt3a conditioned medium (50%), and Wnt3a conditioned medium (50%) including Ti-parti- cles (1:160). β-catenin was labeled with anti-β-catenin antibody and detected with anti-mouse antibody conjugated with FITC. DAPI was used for nucleus staining.
β
β β
β
A B
Ti 입자를 처리한 실험군에서 두 유전자 모두 현 저하게 감소하고 있음을 보여주었다(Fig. 4). 또 한, 이들의 발현 양상은 Ti 입자에 의한 OPG/
RANKL 비율의 감소에서와 유사한 농도 의존적 변화를 나타내고 있다.
고 찰
골용해는 인공 고관절 전치환술에서 장기간 사 용한 뒤 인공관절면에서 발생된 마모편에 의한 생 물학적 반응의 결과로 삽입물 주변부의 골조직의 손실 및 인공관절의 고정력이 소실되어 그 수명을
마치게 하는 주요한 합병증이다5). 골용해의 진행 에 있어서 마모편에 대한 향염 반응이 주요한 요 인이라는 이론은 여러 실험적 증거들에 의해서 뒷 받침되어오고 있다. 일반적으로 마모편을 분리된 대식세포에 함께 배양 하였을 때, 혹은 골용해 동 물모델에 도입하였을 경우 pro-inflammatory cytokine들의 합성을 개시하는 현상을 나타낸다.
결국 TNFα나 IL-1과 같은 대식세포로부터 분비 된 pro-inflammatory cytokine들은 직접적으 로 혹은 RANKL의 합성 증가를 통해서 파골세 포의 생성과 활성을 증가 시키게 되고, 이는 인공 관절 후 발생하는 골용해증에서 나타나는 골흡수
Fig. 4. Increased expression of DKK1 and sFRP2 transcriptional level after treatment of Ti-particles in osteoblasts. Total RNA was isolated from Ti-treated MC3T3-E1 cells. DKK1 and sFRP2 were detected by real-time RTPCR. The expression level of DKK1 and sFRP2 was demon- strated as a relative expression level to GAPDH.
Fig. 3. Down-regulation of LRP5/6 in transcriptional level after treatment of Ti-particles in osteoblasts. Total RNA was isolated from Ti-treated MC3T3-E1 cells. LRP5 and LRP6 were detected by real-time RTPCR. The expression level of LRP5 and LRP6 was demonstrated as a relative expression level to GAPDH.
(bone-resorption)로 이어진다1,7,16). 이와 같은 연구 결과들은 골용해를 설명하는 비교적 단순화 된 모델을 제공하고 있다.
마모편은 대식세포는 물론 인근 조직의 조골세 포 등의 다른 종류의 세포의 활동에도 영향을 미 치게 된다. 골용해에서 보여지는 과도한 파골세포 형성을 완전히 이해하기 위해서는 파골세포 형성 을 조절하는 여러 cytokine들의 신호전달에 관여 할 수 있는 마모편의 다양한 영향을 이해하여야 한다. 최근 연구에 의하면 Ti이 대식세포에서 anti-osteoclastogenic 활성을 갖는 IFN-γ와 IL-6 신호전달을 각각 p38의 활성과 SOCS3, 그 리고 SOCS1을 통해서 저해한다는 보고를 하기도 하였다17). 따라서, 마모편이 골용해의 발생과 진 행에 어떻게 관여하는가에 대하여 마모편이 다양 한 표적 세포들에 미치는 영향, 인근 세포들간의 상호 작용들에 대한 자세한 이해가 필요하다. 특 히 조골세포와 파골세포와 관련된 영향은 골 재형 성의 항상성 기전에 주로 영향을 미치게 되므로 주의를 요한다.
임상적 관점에서 인공관절 후 발생하는 골용해 증은 단순히 골 흡수에 의한 현상으로만 설명 할 수 없으며 골형성능 저하에 대한 초점을 두고 접 근하여야 치료 타깃으로 발전시킬 수 있다. 실제 로 실험적으로도 폴리에틸렌과 금속 입자는 조골 세포에 의하여 탐식되어 골과 기질 형성을 억제하 게 된다6). 이 현상은 in vitro에서 대식세포에서 처럼 마모편의 크기, 조성, 용량에 따라 결정된다.
골 형성능의 억제와 함께 마모 입자는 조골세포에 서의 RANKL, OPG, IL-1, TNF, IL-11 생성 에 영향을 미치게 된다12). UHMWPE (Ultra- High Molecular-Weight Polyethylene)는 인 간 조골세포에서의 RANKL의 분비를 증가시키며 OPG를 억제한다. 실제로 인간 조골세포에 UHMWPE 마모편을 처리한 배양액에서 파골세 포 형성능의 증가가 확인되었다7-9). 그러므로 조골 세포는 마모편에 반응하여 골형성능의 억제와 파 골세포 분화 유도를 통하여 삽입물 주위 골용해증 병인에 기여하고 있음이 분명 하며 이는 향후 집 중해야 할 내용들이다.
본 연구에서는 Ti 입자를 사용하여 골용해 상황 에서 조골세포에 직접적인 영향을 줄 수 있는 신
호전달 중 Wnt 신호전달계의 관련성을 규명하고 자 하였다. 골대사 및 골형성의 조절에 관여하는 여러 신호전달 가운데, 당단백질로 구성된 Wnt family는 골형성과 골 재형성을 포함한 다양한 세포활동에 관여하고 있다. 발생 과정 혹은 세포 분화시에 Wnt 신호전달계는 mesenchymal progenitor들이 adipocytes와 chondrocytes로 의 분화를 억제하면서 골형성을 일으키는 조골세 포로의 분화를 촉진시키는 역할을 하는 것으로 알 려져있다11). 또한, 조골세포에서의 Wnt 신호전달 계는 파골세포의 형성을 조절하는 것에도 관여한 다. Wnt 신호전달이 조골세포에서의 OPG의 발 현을 증가시키며, OPG는 결국 파골세포 형성의 필수 인자인 RANKL을 억제하게 된다. 따라서, Wnt 신호전달이 조골세포에서 RANKL의 억제 와 OPG의 증가를 통하여 파골세포에 의한 골흡 수를 억제하며, 이는 Wnt 신호전달계가 조골세 포를 통한 골형성은 물론 파골세포를 통한 골흡수 에도 관여함을 보여주는 것이다13).
실험 결과, OPG/RANKL 비율은 Ti 입자를 조골세포에 처리한 후 감소되었는데, 이를 조절하 는 기작에 Wnt/β-catenin 신호전달 경로가 관여 할 수 있는 가능성을 Wnt 수용체인 LRP5, LRP6의 발현 감소 및 Wnt 저해인자인 DKK1, sFRP2의 발현 증가를 통하여 확인할 수 있었다.
이는 canonical 신호전달 경로인 β-catenin을 감 소시키는 기작이 다양한 수준에서 이루어질 수 있 음을 시사한다. 결국 이러한 유전자 발현의 변화 가 Wnt/β-catenin 신호전달 경로의 활성화에 직 접적인 영향을 주게 됨으로써 파골세포 형성을 조 절할 뿐만 아니라 조골세포에 의한 골형성 작용에 도 주요하게 관여할 수 있을 것이라 사료된다.
최근 골형성의 조절에 다양한 역할을 보여주는 Wnt/β-catenin 신호전달 경로에 대한 많은 연구 들이 진행되고 있으며, 임상 연구 결과들 또한 LRP5의 유전자 변이가 골밀도 및 골절과 관련되 어 있음을 보여주고 있다. Wnt-10b, LRP-5/6, sFRP-1, DKK-2, Axin-2, β-catenin과 같은 Wnt 신호전달 경로의 다양한 유전자들에 관한 형질전환 및 유전자 결핍 동물 연구들은 canoni- cal Wnt 신호전달 경로가 파골세포 형성 및 골 흡수 뿐만이 아니라 골기질 형성, 세포분화, 세포
분열, 세포사멸 등을 포함하는 다양한 조골세포의 생리활성의 조절에 관여하고 있음을 증명한다2). 이러한 Wnt 신호전달 경로에 대하여, 조골세포 의 기능과 관련된 다양한 Wnt 리간드들과 수용 체들의 조합이 갖는 특이성, β-catenin을 경유하 는 canonical 경로 이외의 noncannonical 경로 들의 가능한 역할, 골 재형성 과정에서 파골세포 에 미칠 수 있는 직접적인 영향 등에 관한 향후 연구의 필요성을 제시 하고자 한다.
결 론
Ti 입자는 조골세포에서 Wnt 수용체 발현을 억제하고, 신호저해 조절물질의 발현을 증가시켜 조골세포에서의 β-catenin 신호전달 경로의 활성 화를 억제하였다. 이로 인하여 파골세포 조절에 관여하는 OPG/RANKL 비율을 감소시키며 Wnt 신호전달에 의한 골형성기능을 저해할 것으 로 사료된다.
REFERENCES
1) Baumann B, Rader CP, Seufert J et al.: Effects of polyethylene and TiAlV wear particles on expression of RANK, RANKL and OPG mRNA.
Acta Orthop Scand.75: 295-302, 2004.
2) Bodine C, Scherer MJ: Technology for improv- ing cognitive function. A workshop sponsored by the US Interagency Committee on Disability Research (ICDR): reports from working groups.
Disabil Rehabil. 28: 1567-71, 2006.
3) Cianferotti L,Demay MB: VDR-mediated inhi- bition of DKK1 and SFRP2 suppresses adi- pogenic differentiation of murine bone marrow stromal cells. J Cell Biochem. 101: 80-8, 2007.
4) Dean DD, Schwartz Z, Blanchard CR et al.:
Ultrahigh molecular weight polyethylene parti- cles have direct effects on proliferation, differen- tiation, and local factor production of MG63 osteoblast-like cells. J Orthop Res. 17: 9-17, 1999.
5) Gallo J, Kaminek P, Ticha V, Rihakova P,Dit-
mar R: Particle disease. A comprehensive theory of periprosthetic osteolysis: a review. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 146: 21-8, 2002.
6) Goodman SB, Ma T, Chiu R, Ramachandran R,Smith RL: Effects of orthopaedic wear parti- cles on osteoprogenitor cells. Biomaterials. 27:
6096-101, 2006.
7) Granchi D, Amato I, Battistelli L et al.: Molec- ular basis of osteoclastogenesis induced by osteoblasts exposed to wear particles. Biomateri- als. 26: 2371-9, 2005.
8) Granchi D, Ciapetti G, Amato I et al.: The influence of alumina and ultra-high molecular weight polyethylene particles on osteoblast-osteo- clast cooperation. Biomaterials. 25: 4037-45, 2004.
9) Gutwein LG,Webster TJ: Increased viable osteoblast density in the presence of nanophase compared to conventional alumina and titania particles. Biomaterials. 25: 4175-83, 2004.
10) Heider U, Hofbauer LC, Zavrski I, Kaiser M, Jakob C, Sezer O: Novel aspects of osteoclast activation and osteoblast inhibition in myeloma bone disease. Biochem Biophys Res Commun.
338: 687-93, 2005.
11) Hill TP, Spater D, Taketo MM, Birchmeier W,Hartmann C: Canonical Wnt/beta-catenin signaling prevents osteoblasts from differentiat- ing into chondrocytes. Dev Cell. 8: 727-38, 2005.
12) Hofbauer LC, Khosla S, Dunstan CR, Lacey DL, Boyle WJ, Riggs BL: The roles of osteo- protegerin and osteoprotegerin ligand in the paracrine regulation of bone resorption. J Bone Miner Res. 15: 2-12, 2000.
13) Holmen SL, Zylstra CR, Mukherjee A et al.:
Essential role of beta-catenin in postnatal bone acquisition. J Biol Chem. 280: 21162-8, 2005.
14) Kieslinger M, Folberth S, Dobreva G et al.:
EBF2 regulates osteoblast-dependent differentia- tion of osteoclasts. Dev Cell. 9: 757-67, 2005.
15) Lohmann CH, Dean DD, Koster G et al.:
Ceramic and PMMA particles differentially affect osteoblast phenotype. Biomaterials. 23:
1855-63, 2002.
16) Purdue PE, Koulouvaris P, Potter HG, Nestor BJ,Sculco TP: The cellular and molecular biolo- gy of periprosthetic osteolysis. Clin Orthop Relat Res. 454: 251-61, 2007.
17) Rakshit DS, Ly K, Sengupta TK et al.: Wear Debris Inhibition of Anti-Osteoclastogenic Sig- naling by Interleukin-6 and Interferon-{gamma}
Mechanistic Insights and Implications for Periprosthetic Osteolysis. J Bone Joint Surg Am.
88: 788-99, 2006.
18) Vermes C, Chandrasekaran R, Jacobs JJ, Galante JO, Roebuck KA,Glant TT: The effects of particulate wear debris, cytokines, and growth factors on the functions of MG-63 osteoblasts. J Bone Joint Surg Am. 83-A: 201- 11, 2001.
19) Wang ML, Nesti LJ, Tuli R et al.: Titanium particles suppress expression of osteoblastic phe- notype in human mesenchymal stem cells. J Orthop Res. 20: 1175-84, 2002.
20) Zou L, Zou X, Li H et al.: Molecular mecha- nism of osteochondroprogenitor fate determina- tion during bone formation. Adv Exp Med Biol.
585: 431-41, 2006.