50(4) : 291 ∼ 298 (2019)
291
꽃잔디 메탄올 추출물의 RAW264.7 대식세포와 BV2 미세아교세포에서의 항염증 효과
김관우1,2#·이 정1,2#·이 환3·이동성3·오현철1,2·김윤철1,2*
1원광대학교 약학대학, 2원광대학교 약학대학 약품연구소, 3조선대학교 약학대학
Anti-inflammatory Effects of the Methanol Extracts of Phlox subulata on LPS-induced RAW264.7 Macrophages and BV2 Microglia
Kwan-Woo Kim1,2#, Jing Li1,2#, Hwan Lee3, Dong-Sung Lee3, Hyuncheol Oh1,2 and Youn-Chul Kim1,2*
1College of Pharmacy, Wonkwang University
2Institute of Pharmaceutical Research and Development, College of Pharmacy, Wonkwang University
3College of Pharmacy, Chosun University
Abstract Phlox subulata is a perennial herbaceous flower and is a member of the Polemoniaceae family. This plant is known to resist to various stresses including salt, drought, heat, and cold stresses. In this investigation, we evaluated the ant-inflam- matory effect of the methanolic extract of P.subulata(PSM) on lipopolysaccharide(LPS)-induced RAW264.7 macrophages and BV2 microglia. PSM reduced the production of nitric oxide(NO) in LPS-stimulated both RAW264.7 and BV2 cells, but did not affect to the production of prostaglandin E2(PGE2). It inhibited the expression of inducible nitric oxide synthase(iNOS) and cyclooxygenase(COX)-2 in both cells. In addition, PSM suppressed the production of pro-inflammatory cytokines including interleukin(IL)-6 and tumor necrosis factor(TNF)-. These inhibitory effects were contributed by inactivation of nuclear factor kappa B(NF-κB) and mitogen-activated protein kinases(MAPKs) pathways by PSM. Thus, these results suggested that P.sub- ulata can be a candidate material to treat inflammatory diseases.
Keywords Phlox subulata, Lipopolysaccharide, Inflammation, RAW264.7 macrophage, BV2 microglia
염증반응은 체내 혹은 체외의 다양한 자극 및 유해인자 에 반응하여 나타나는 면역 반응으로써, 손상된 세포의 기 능을 정상화를 유도하는 일련의 생체 방어 기전이다.1) 체 내에서 염증반응은 주로 대식세포와 미세아교세포에 의해 서 조절되는데, 염증 반응이 과도하게 발생할 경우 이들 세포에서 염증 반응을 심화시키는 nitric oxide(NO), prostaglandin E2(PGE2), inducible nitric oxide synthase (iNOS), cyclooxygenase-2(COX-2)와 같은 염증 매개체들과, interleukin-1(IL-1), IL-6, tumor necrosis factor-(TNF-
)와 같은 염증성 싸이토카인의 분비량이 증가하여 2형 당 뇨, 류마티스 관절염, 심혈관계 질병, 암, 알츠하이머 병, 파 킨슨 병, 다발성 경화증 등의 병리적 상태에 이르게 할 수 있다.2-4) 따라서, 이러한 염증 유발 매개체들과 염증성 싸이
토카인의 분비를 조절하는 것이 염증성 질환을 치료할 수 있는 방법으로 제시되고 있다.
꽃잔디(Phlox subulate)는 여러해살이 초본식물 중 꽃고비 과에 속하는 꽃 식물이며, 미국 중부 및 동부가 원산지로 알 려져 있다. 염증을 줄여주고 통증을 완화하는 효능이 있어 서, 원산지인 미국 중부 및 동부의 원주민들은 감기나 류마 티스 관절염에 사용했다고 알려져 있다.5) 분홍색이나 흰색 의 아름다운 꽃이 피기 때문에 최근에는 관상용으로 많이 재배하고 있는 식물이다.6) 이 식물은 염분, 가뭄, 고열, 저 온 등의 강한 자극에 대해 내성을 가지고 있다. 특히, 영하 40도까지 떨어지는 겨울 동안 살아남을 수 있고, 녹색 표현 형을 나타내는 유전자는 영하 12도에서 유지할 수 있음이 알려져 있다.7,8) 이와 같은 특징은 꽃잔디에 포함되어 있는 Cor413과 유사한 유전자에 의한 것임이 밝혀진 바 있다. 또 다른 연구에서 꽃잔디 꽃잎에 포함된 성분을 확인한 결과, 꽃잔디 꽃잎의 에탄올 추출물에는 sucrose, glucose, fructose,
#
These authors contributed equally to this work.
*교신저자(E-mail):[email protected]
(Tel): +82-63-850-6823
myo-inositol, soribitol, 2-C-methyl-D-erythritol이 함유 되어 있는 것으로 확인되었다.6) 이와 같이 꽃잔디의 성분 확인 및 특징과 관련된 유전자의 연구는 보고되었으나, 염증 반 응에 대한 효능 및 기전에 관한 연구는 보고된 바가 없었 다. 따라서, 본 연구에서는 꽃잔디의 메탄올 추출물을 이용 하여 대식세포 및 미세아교세포에 적용하여 염증 반응에 대 한 활성 반응을 확인하고 그 기전을 확인하고자 하였다.
재료 및 방법
실험재료 본 실험에 사용한 꽃잔디(Phlox subulate)는 2011년 6월 익산시 원광대학교에서 채집하였으며, 형태학적 평가를 통하여 동정하였고, 원광대학교 김윤철 교수에 의해 시료를 검증하였다. 채집한 표본시료는 원광대학교 약학대 학에 보관하였다.
꽃잔디 메탄올 추출물 제조 꽃잔디(Phlox subulate)의 전체 부위(whole plant)를 건조한 샘플(50 g)을 실온에서 메 탄올 500 mL에 1시간 방치 후 80도에서 3시간 환류 추출 하였다. 추출액을 진공 하에서 농축하여 추출물(3.5 g)을 얻 었다. 세포 실험을 진행하기 위해 추출물을 DMSO(Dimethyl sulfoxide)에 녹여 시료를 제조하였다.
시약 및 기기 RPMI1640 배지와 trypsin-ethylenediami- netetraacetic acid(EDTA), fetal bovine serum(FBS)는 Gibco Laboratories사에서 구입하였다. Lipopolysaccharide(LPS, 055:
B5)와 3'-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide(MTT)는 Sigma사에서 구입하였다. 96-Well tissue culture plates와 기타 tissue culture dishes는 Falcon사 제품 을 이용하였다. 흡광도는 BioRad사의 Microplate Reader를 이용하여 측정하였다.
RAW264.7, BV2 세포 배양 생쥐 대식세포 유래 RAW264.7 세포주와 생쥐 미세아교세포 유래 BV2 세포주 는 기존에 보고된 문헌을 참조하여 배양하였다.9)
Determination of nitrite 배양된 RAW264.7 및 BV2세포 에서 medium으로 분비되어 나온 NO의 양을 Griess 시약을 사용하여 측정하였으며, 자세한 방법은 기존에 보고된 문헌 을 참조하여 실시하였다.9)
PGE2, IL-6, and TNF- assay 배양된 RAW264.7 및 BV2 세포에서 medium으로 분비되어 나온 PGE2 및 염증성 cytokine인 IL-6, TNF-의 측정은 R&D systems(Minneapolis, MN)에서 구입한 enzyme-linked immunosorbent assay(ELISA) kit를 사용하여 측정하였으며, 자세한 방법은 구입한 제품에 동봉된 안내서를 참조하여 실시하였다.
Western Blot Analysis RAW264.7 및 BV2 세포를 60 mm dish에 3×105cells/well 농도로 24시간 배양한 후 꽃잔 디 메탄올 추출물 시료를 농도별로 처리하였다. RAW264.7 및 BV2 세포에 RIPA buffer를 첨가한 다음, 4oC, 14,000×g
에서 20분간 원심분리하고 상등액을 튜브에 옮겼다. 단백질 정량은 BSA 단백질 실험 키트를 이용하였고 각각의 시료 를 7.5% 및 12% SDS-polyacrylamide gel에서 영동하고 Nitrocellulose membrane(NC membrane)으로 전사하였다.
전사된 NC membrane을 5% 무지방유가 포함된 신선한 blocking buffer(0.1% Tween 20 in Tris-buffered saline)에 서 blocking한후 iNOS, COX-2, p-p65, p-IB-, IB-, p- JNK, p-p38, p-ERK, JNK, p38, ERK, -actin antibody를 1:1000으로 희석하여 넣고 2시간 동안 4oC 조건에서 반응 시켰다. 다시 2차 항체(Anti-mouse, Anti-rabbit IgG)를 1:1000으로 희석하여 넣고 1시간 동안 반응한 다음, ECL 용액을 1:1로 잘 섞어서 NC membrane위에 부어서 발광시 키고 ChemiDoc MP System(Bio-Rad)를 이용하여 단백질의 발현을 확인하였다.
통계처리 본 실험의 통계처리는 GraphPad Prism, version 3.03(GraphPad Software Inc., San Diego, CA)을 사용하였다. 각 실험군의 결과는 평균치와 표준오차로 나타 내었으며, 각 실험군 간의 결과는 ANOVA test를 사용하여 분석하고 유의적인 차이가 있는 항목에 대해서만 검정하였 다. 실험군 간의 차이는 95% 수준(p<0.05)에서 유의성 있 는 것으로 하였다.
결과 및 고찰
본 연구에서는 꽃잔디 메탄올 추출물에 대하여 LPS로 염 증 반응을 유발시킨 RAW264.7 대식세포 및 BV2 미세아교 세포에 있어서의 항염증 효과를 탐색하여 그 작용기전을 밝 히고자 하였다.
꽃잔디 메탄올 추출물이 대식세포 및 미세아교세포에서 항염증 효과를 나타내는지 확인하기 위하여 LPS로 유발되 는 염증 관련 인자들의 억제여부를 확인하였다. 우선, MTT assay를 통하여 꽃잔디 메탄올 추출물의 RAW264.7 대식세 포 및 BV2 미세아교세포에서의 독성을 확인하여 세포독성 을 나타내지 않는 농도를 확인하였다. 꽃잔디 메탄올 추출 물을 12.5~400 μg/mL의 농도로 처리한 후, 24시간 동안 CO2 incubator에서 배양한 후 세포 생존율을 확인한 결과, RAW264.7 세포에서는 200 μg/mL까지(Fig. 1A), BV2 세포 에서는 100 μg/mL까지 세포 독성이 나타나지 않았다(Fig.
1B). 이를 바탕으로 이후 실험에 대하여 RAW264.7 세포에 서는 200 μg/mL, BV2 세포에서는 100 μg/mL을 최고 농도 로 설정하였다.
염증 반응이 과도하게 발생할 경우, 대표적인 염증세포인 대식 세포와 미세아교세포가 활성화되어 NO 및 PGE2의 분 비가 과도하게 발생한다.10) 염증 반응시에 NO는 체내 arginine으로부터 iNOS 단백질에 의해 생성되고,11), PGE2, PGD2, PGI2, PGF2와 같은 prostaglandin류와 thromboxane
A2(TXA2)는 arachidonic acid로부터 COX-2 단백질의 작용 에 의해 생성된다.12) 이들 인자들은 염증 반응의 매개하는 중요한 물질이며, 염증 과정이 발생하는 동안 이들 전염증 매개체들을 다량으로 생성하는 것으로 보고되어 있다. 따라 서, 저자들은 꽃잔디 메탄올 추출물이 염증 반응이 유도된 RAW264.7 대식세포 및 BV2 미세아교세포에서 NO 및
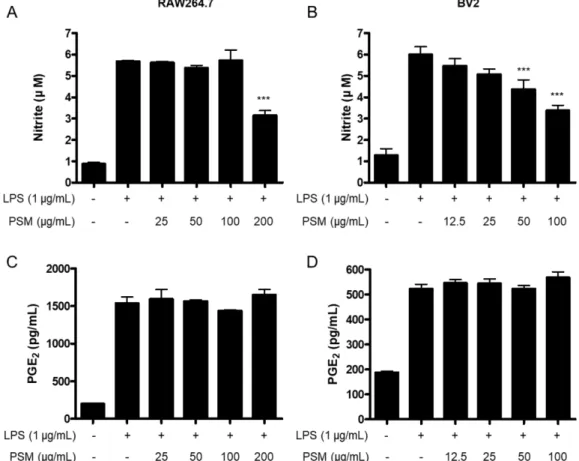
PGE2의 생성, 그리고 iNOS 및 COX-2 단백질의 조절에 관 여하는지를 확인하고자 하였다. RAW264.7 세포에서는 200 μg/mL 농도에서만 NO의 생성을 감소시켰으며, BV2 세포 에서는 농도 의존적으로 NO의 생성을 감소시켰다(Fig. 2A, B). 그러나, PGE2의 생성은 RAW264.7 및 BV2 세포 모두 에서 꽃잔디 메탄올 추출물에 의한 감소효과가 나타나지
Fig. 1. The effect of PSM on RAW264.7 (A) and BV2 (B) cell viability. Cells were treated with indicated concentrations with PSM for 24 h. The viability was determined by MTT Assay. *p<0.05, and ***p<0.001 compared to the control group.
Fig. 2. The effect of PSM on nitrite and PGE2 production in LPS-induced RAW264.7 (A, C) and BV2 cells (B, D). Cells were pre- treated with indicated concentrations of PSM for 3 h, and then stimulated with LPS (1 μg/mL) for 24 h. The concentration of nitrite and PGE2 was determined as described on Materials and methods. Data represent the mean values of three independent experiments
± SD. ***p<0.001 compared to the LPS treated group.
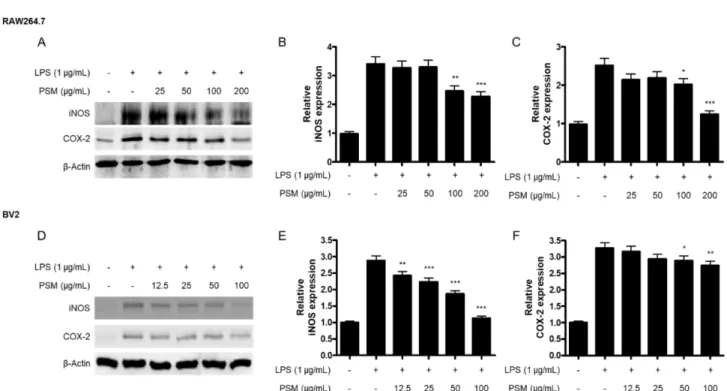
Fig. 3. The effect of PSM on iNOS and COX-2 protein expression in LPS-induced RAW264.7 (A, B, C) and BV2 cells (D, E, F).
Cells were pre-treated with indicated concentrations of PSM for 3 h, and then stimulated with LPS (1 μg/mL) for 24 h. Western blot analyses were performed as described in Materials and methods. Representative blots of three independent experiments are shown. *p<0.05, **p<0.01, and ***p<0.001 compared to the LPS treated group.
Fig. 4. The effect of PSM on the production of IL-6 and TNF- in LPS-induced RAW264.7 (A, C) and BV2 cells (B, D). Cells were pre-treated with indicated concentrations of PSM for 3 h, and then stimulated with LPS (1 μg/mL) for 24 h. The concentration of IL-6 and TNF- was determined as described on Materials and methods. Data represent the mean values of three independent experiments ± SD. *p<0.05, **p<0.01, and ***p<0.001 compared to the LPS treated group.
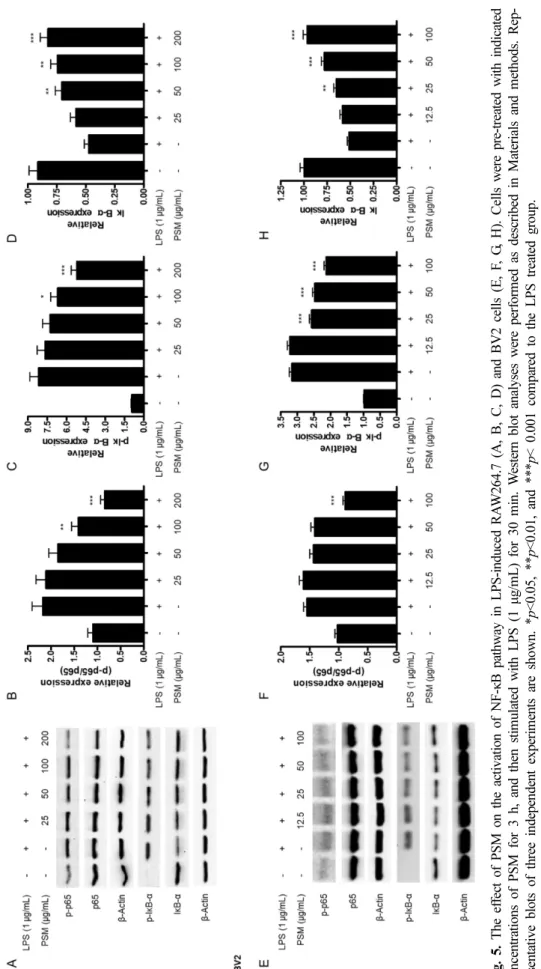
Fi g. 5 . T he ef fe ct of PSM on the activa tion of NF-κB pathwa y in LPS-induce d RA W 264. 7 (A , B , C , D) and BV2 cells ( E, F , G , H ). Ce lls w ere pre-treated w ith ind icated conc entrations o f PSM f or 3 h , and th en stim ulate d with LPS (1 μg/m L) for 30 m in. W es tern blot analy ses were perf or m ed as d es cr ibed i n M aterials a nd m ethod s. Re p- re se ntative blots of three in depen dent experim ents are show n. * p<0.05, ** p< 0. 01 , and *** p< 0.001 co m pared to the LPS trea ted gr ou p.
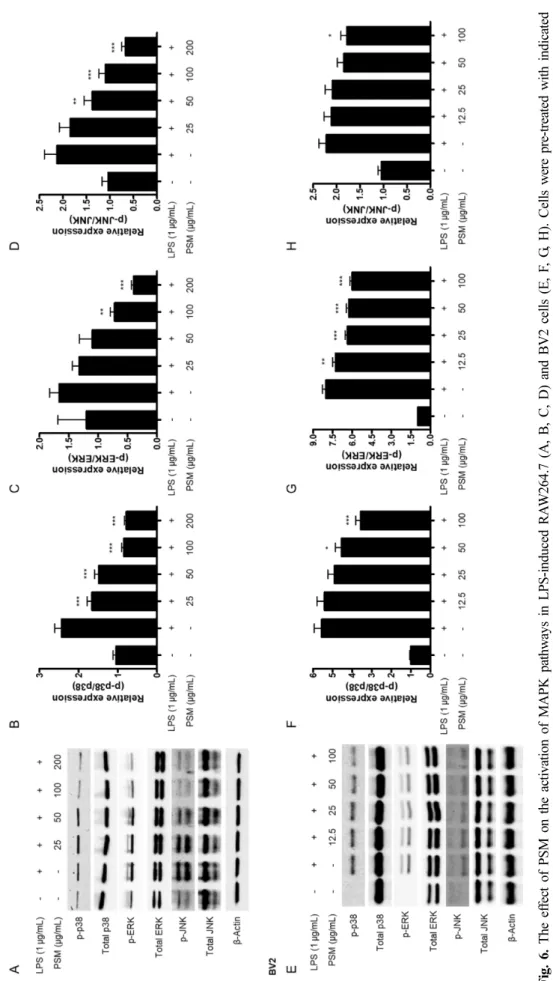
Fi g. 6 . T he ef fe ct of PSM on the activa tion of M APK pathwa ys in LPS-induc ed RA W 264 .7 (A, B, C, D) and BV 2 ce lls (E , F , G , H). Ce lls we re p re-treate d w ith ind icated conc entrations o f PSM f or 3 h , and th en stim ulate d with LPS (1 μg/m L) for 30 m in. W es tern blot analy ses were perf or m ed as d es cr ibed i n M aterials a nd m ethod s. Re p- re se ntative blots of three in depen dent experim ents are show n. * p<0.05, ** p< 0. 01 , and *** p<0.001 co m pared to the LPS trea ted group.
않았다(Fig. 2C, D). 이후, 저자들은 꽃잔디 메탄올 추출물 이 iNOS 및 COX-2 단백질의 조절에 관여하는지를 확인하 고자 하였다. 꽃잔디 메탄올 추출물은 RAW264.7 및 BV2 세포 모두에서 iNOS 및 COX-2 단백질의 발현을 억제하는 것으로 확인되었다(Fig. 3). 특이하게도, 꽃잔디 메탄올 추 출물에 의한 PGE2의 감소는 발생하지 않았으나, PGE2의 생 성을 담당하는 COX-2 단백질의 발현은 억제가 되는 것을 확인할 수 있었다. Arachidonic acid가 COX-2에 의해 PGH2 로 변환된 후, prostaglandin E synthase(PGES)의 작용으로 인해 PGE2가 생성되는 것으로 알려져 있다.13,14) 따라서, 꽃 잔디 메탄올 추출물이 PGES의 활성에 미치는 영향에 대한 추가적인 연구가 필요할 것으로 사료된다.
염증 세포가 활성화하면 NO 및 PGE2 뿐만 아니라 IL- 1, IL-6, TNF- 등의 염증성 싸이토카인이 과다 분비되어 염증 반응을 유지시키거나 악화시킬 수 있다.15) 따라서, 염 증성 싸이토카인의 분비를 조절함으로써 염증성 질환의 발 생의 예방 및 치료 효과를 기대할 수 있다. 이에 따라 꽃잔 디 메탄올 추출물의 LPS로 유도한 염증성 싸이토카인인 IL- 6와 TNF-의 분비에 미치는 영향을 확인하고자 하였다. 두 가지 세포 모두에서 꽃잔디 메탄올 추출물을 최고 농도인 200(RAW264.7) 및 100(BV2) μg/mL에서 IL-6 및 TNF-
분비를 억제하였다(Fig. 4.).
Nuclear factor kappa B(NF-κB) 경로는 염증 인자 분비에 필수적인 전사 인자로 알려져 있다. 자극이 없는 조건에서 NF-κB의 구성 인자인 p65 및 p50 단백질은 세포질 내에서 inhibitor kappa B-(IκB-)와 결합된 상태로 존재한다. LPS 자극으로 인해 세포내 기전들이 활성화가 되면 p65 및 p50 이 IκB-와 분리되고, p65의 인산화 반응이 발생하고, IκB-
는 인산화가 발생한 후 소멸하게 된다.16,17) 분리된 p65와 p50은 핵 내부로 이동하게 되어 DNA와 결합하게 되어 iNOS, COX-2, NO, PGE2, TNF-, IL-6 등의 염증성 인자 의 유전자의 전사를 활성화시킨다.18) 따라서, 저자들은 꽃 잔디 메탄올 추출물이 NF-κB 경로의 조절에 관여하는지를 확인하고자 하였다. 꽃잔디 메탄올 추출물 처리에 의해 RAW264.7 및 BV2 세포에서 p65의 인산화 반응, IκB-의 인산화 반응 및 소멸 반응이 억제되었다(Fig. 5).
Mitogen-activated protein kinase(MAPK) 경로는 세포 사 멸, 세포 성장, 세포 분화, 세포 증식 및 면역 반응에 중요 한 역할을 하는 것으로 알려져 있다.19) MAPK 경로에 관여 하는 3가지 주요한 단백질로는 extracellular signal regulated kinases 1, 2(ERK1/ERK2), c-Jun N-terminal kinases(JNKs), p38 MAPK가 있다.20) 기존 여러 연구들에서 MAPK경로가 LPS로 유도한 염증 반응에 관여함을 증명하였다.21-23) 이에 따라 저자들은 꽃잔디 메탄올 추출물이 MAPK 경로의 조 절에 관여하는지를 확인하고자 하였다. 그 결과, LPS 자극 으로 인한 p38, ERK, JNK의 인산화 반응이 두 가지 세포
모두에서 억제되는 것을 확인하였고, total p38, total ERK, total JNK의 발현양에는 변화가 없음을 확인하였다(Fig. 6.).
결 론
본 연구에서는 꽃잔디 메탄올 추출물(PSM)의 LPS로 염 증 반응을 유도한 RAW264.7 대식세포 및 BV2 미세아교세 포에서의 항염증 활성과 그 기전을 확인해보고자 하였다.
꽃잔디 메탄올 추출물은 RAW264.7 및 BV2 세포에서 LPS 자극으로 인한 NO, IL-6, TNF- 등의 생성을 억제하였으 나, PGE2의 생성은 억제하지 못하였다. 또한, NO 및 PGE2 생성에 관여하는 단백질인 iNOS 및 COX-2의 발현이 감소 하는 결과를 얻었다. 이와 같은 항염증 효과는 NF-κB 및 MAPK 경로의 활성화의 억제에 의한 것으로 판단된다. 특 이한 것은, PGE2의 생성은 억제하지 못하였으나, PGE2 생성 에 관여하는 단백질인 COX-2의 발현은 억제하는 점인데, 이는 PGE2 생성에 관여하는 또 다른 단백질인 PGES에 대 한 추가적인 연구가 필요함을 시사하고 있다. 이와 같은 연 구 결과는 꽃잔디 메탄올 추출물이 항염증 효과를 나타냄 으로써 추후에 꽃잔디가 천연물 유래 신약 후보 물질이 될 수 있음을 보여주는 기초적인 실험 근거를 제시하는 것으 로, 추후 함유성분 가운데 직접적인 효과를 나타내는 성분 을 확인하는 연구가 진행되어야 할 것으로 사료된다.
사 사
이 논문은 2018학년도 원광대학교의 지원에 의해서 수행 되었음.
인용문헌
1. Nathan, C. (2002) Points of control in inflammation. Nature 430: 846-852.
2. Zhu, T., Zhang, W., Feng, S. J. and Yu, H. P. (2016) Emodin suppresses LPS-induced inflammation in RAW264.7 cells through a PPARγ-dependent pathway. Int. Immunopharma- col. 34: 16-24.
3. Zhang, H., Shan, Y., Wu, Y., Xu, C., Yu, X., Zhao, J., Yan, J.
and Shang, W. (2017) Berberine suppresses LPS-induced inflammation through modulating Sirt1/NF-κB signaling pathway in RAW264.7 cells. Int. Immunopharmacol. 52: 93- 100.
4. Lull, M. E. and Block, M. L. (2010) Microglial activation &
chronic neurodegeneration. Neurotherapeutics 7: 354-365.
5. Romero, J. B. (1954) The Botanical Lore of the California Indians. Vantage Press. pp 59.
6. Enomoto, H., Kohata, K., Nakayama, M., Yamaguchi, Y. and Ichimura, K. (2004) 2-C-methyl-D-erythritol is a major car-
bohydrate in petals of Phlox subulata possibly involved in flower development. J. Plant. Physiol. 161: 977-980.
7. Qu, Y., Zhou, A., Zhang, X., Tang, H., Liang, M., Han, H.
and Zuo, Y. (2015) De novo transcriptome sequencing of low temperature-treated Phlox subulata and analysis of the genes involved in cold stress. Int. J. Mol. Sci. 16: 9732-9748.
8. Zhou, A., Sun, H., Feng, S., Zhou, M., Gong, S., Wang, J.
and Zhang, S. (2018) A novel cold-regulated gene from Phlox subulata, PsCor413im1, enhances low temperature tol- erance in Arabidopsis. Biochem. Biophys. Res. Commun.
495: 1688-1694.
9. Ko, W., Sohn, J. H., Jang, J. H., Ahn, J. S., Kang, D. G., Lee, H. S., Kim, J. S., Kim, Y. C. and Oh, H. (2016) Inhibitory effects of alternaramide on inflammatory mediator expression through TLR4-MyD88-mediated inhibition of NF-кB and MAPK pathway signaling in lipopolysaccharide-stimulated RAW264.7 and BV2 cells. Chem. Biol. Interact. 244: 16-26.
10. Sharma, J. N., Al-Omran, A. and Parvathy, S. S. (2007) Role of nitric oxide in inflammatory diseases. Inflammopharma- cology 15: 252-259.
11. Tang, Y. and Le, W. (2016) Differential roles of M1 and M2 microglia in neurodegenerative diseases. Mol. Neurobiol. 53:
1181-1194.
12. Andreasson, K. (2010) Emerging roles of PGE2 receptors in models of neurological disease. Prostaglandins Other Lipid Mediat. 91: 104-112.
13. Ikeda-Matsuo, Y. (2017) The Role of mPGES-1 in Inflam- matory Brain Diseases. Biol. Pharm. Bull. 40: 557-563.
14. Pierre, C., Guillebaud, F., Airault, C., Baril, N., Barbouche, R., Save, E., Gaigé, S., Bariohay, B., Dallaporta, M. and Troadec, J. D. (2018) Invalidation of microsomal prosta- glandin E synthase-1 (mPGES-1) reduces diet-induced low- grade inflammation and adiposity. Front Physiol. 9: 1358.
15. Du, R. W., Du, R. H. and Bu, W. G. (2014) -Arrestin 2 mediates the anti-inflammatory effects of fluoxetine in lipo-
polysaccharide-stimulated microglial cells. J. Neuroimmune Pharmacol. 9: 582-590.
16. Viatour, P., Merville, M. P., Bours, V. and Chariot, A. (2005) Phosphorylation of NF-kappaB and IkappaB proteins: impli- cations in cancer and inflammation. Trends Biochem. Sci. 30:
43-52.
17. Li, C., Li, L., Chen, K., Wang, Y., Yang, F. and Wang, G.
(2019) UFL1 alleviates lipopolysaccharide-induced cell dam- age and inflammation via regulation of the TLR4/NF-κB pathway in bovine mammary epithelial cells. Oxid. Med.
Cell. Longev. 2019: 6505373.
18. Kim, B. W., Koppula, S., Hong, S. S., Jeon, S. B., Kwon, J.
H., Hwang, B. Y., Park, E. J. and Choi, D. K. (2013) Reg- ulation of microglia activity by glaucocalyxin-A: attenuation of lipopolysaccharide-stimulated neuroinflammation through NF-κB and p38 MAPK signaling pathways. PLoS One 8:
e55792.
19. Dong, C., Davis, R. J. and Flavell, R. A. (2002) MAP kinases in the immune response. Annu. Rev. Immunol. 20: 55–72.
20. Liu, Y., Shepherd, E. G. and Nelin, L. D. (2007) MAPK phos- phatases--regulating the immune response. Nat. Rev. Immu- nol. 7: 202-212.
21. Lai, J. L., Liu, Y. H., Liu, C., Qi, M. P., Liu, R. N., Zhu, X.
F., Zhou, Q. G., Chen, Y. Y., Guo, A. Z. and Hu, C. M. (2017) Indirubin inhibits LPS-induced inflammation via TLR4 abro- gation mediated by the NF-κB and MAPK signaling path- ways. Inflammation 40: 1-12.
22. Yin, P., Zhang, Z., Li, J., Shi, Y., Jin, N., Zou, W., Gao, Q., Wang, W. and Liu, F. (2019) Ferulic acid inhibits bovine endometrial epithelial cells against LPS-induced inflamma- tion via suppressing NK-κB and MAPK pathway. Res. Vet.
Sci. 126: 164-169.
(2019. 11. 21 접수; 2019. 12. 6 심사;
2019. 12. 18 게재확정)