516
Copyright © 2014 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
사료의주성분인어분
(fish meal)
의생산량은어족자원의감 소로인하여정체또는감소하는추세에있어어분의수급불 안정에따른어류양식업의 채산성악화가문제점으로 대두되 고있다.
따라서일부어류양식업체에서는어류의사료를제조 함에있어서단백질함량이높고수급이비교적안정적인탈 지대두박을어분대체소재로많이사용되고있다(Kim et al., 2013).
그러나대두박의탄수화물의경우,
단당류가거의없으 며올리고당은sucrose, maltose, raffinose, stachyose
등으로 구성되어있고 다당류에는arabinnan
등으로구성되어 있다.
특히stachyose
와raffinose
는α-galactosyl
결합으로이루어져 있어소화기관내에α-galactosidase (α-D-galactoside galacto- hydrolase, EC 3,2,1,22)
효소를갖지않은인간과단위동물에 게있어서raffinose
와stachyose
는소화되지않은채대장에도 달하고,
대장내의혐기성세균에의해메탄,
이산화탄소그리 고수소가스가생성되어고창증의원인(Rackis, 1981)
이나복부팽만감의원인
(Reddy et al., 1980; Steggerda et al., 1968)
이될수있다.
또한분뇨내질소배출량증가및악취가발생 할수 있고어류의성장을억제하거나질병을유발시키는것 으로알려져있다(Yoo et al., 2007).
이러한항영양인자의영 향으로소화가잘되지않아어류용사료제조에사용이제한 되고있다(Li et al., 1991).
따라서사료생산과정에서이들을 분해할수있는효소인α-galactosidase
에대한연구가(Min et al., 2002; Kim and Yoon, 1998; Choi et al., 1998; Kim et al., 2004; Irish et al., 1995)
활발히진행되고있으며미생물로부터 생산되는α-galactosidase
특성에관한연구(park., 2011; Kim.
et al., 2002; Kum and Yoon., 1998; Choi et al., 1998)
도다양하 게이루어지고있으나아직까지고온성미생물을활용한발효 대두박제조에대한연구는부족한실정이다.
또한stachyose
와raffinose
같은항영양인자를분해할수있는고온성미생물을탐색하고이에대한활용방안연구가필요한실정이다
.
한편, α-galactosidase
는 식품,
미생물,
동물에 널리분포되 어있으며, galactose
잔기를함유한galactomannan
다당류나α-Galactosidase를 생산하는 고온성 Bacillus coagulans KM-1 균주의 생화학적 특성
남기호*·장미순·박희연·이레나 코네바 1
국립수산과학원 식품안전과, 1태평양수산연구소 틴로센터
Biochemical Characterization of α-Galactosidase-Producing Thermophilic Bacillus coagulans KM-1
Ki Ho Nam*, Mi Soon Jang, Hee Yeon Park and Elena Koneva
1
Food and safety Research Division, National Fisheries Research and Development Institute, Busan 619-705, Korea
1
TINRO Centre, Pacific Scientific Research Fisheries Centre, Shevchenko Alley, Vladivostok 690091, Russia
A bacterium producing α-galactosidase (α- d -galactoside galactohydrolase, EC 3.2.1.22) was isolated. The isolate, KM-1 was identified as Bacillus coagulans based on its 16S rRNA sequence, morphology, and biochemical proper- ties. α-Galactosidase activity was detected the culture supernatant of B. coagulans KM-1. The bacterium showed the maximum activity for hydrolyzing para -nitrophenyl-α- d -galactopyranoside ( p NP-αGal) at pH 6.0 and 50°C. It hydro- lyzed oligomeric substrates such as melibiose, raffinose, and stachyose liberating a galactose residue, indicating that the B. coagulans KM-1 α-galactosidase hydrolyzed α-1,6 linkage. The results suggest that the decreased stachyose and raffinose contents in fermented soybean meal are due to the α-galactosidase activity.
Key words: Bacillus coagulans KM-1, α-galactosidase, Stachyose, Raffinose, Fermented soybean meal
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/)which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2014.0516 Kor J Fish Aquat Sci 47(5) 516-521, October 2014
Received 22 September 2014; Revised 20 October 2014; Accepted 29 October 2014
*Corresponding author: Tel: +82. 51. 720. 2652 Fax: +82. 51. 720. 2669
E-mail address: [email protected]
Bacillus coagulans KM-1의 생화학적 특성
517
melibiose, raffinose, stachyose
등의저당류또는galactoman- nan
에서α-1,6
결합의α-galactose
잔기를가수분해하는것으 로알려져있다(Dey and Campillo, 1984).
이에본연구에서는 멸치(Engraulis japonica)
로부터α-galactosidase
를 생산하는 고온성신규미생물을확보하여생화학적특성을규명하고산 업적으로품질이우수한발효대두박제조에활용하고자한다.
재료 및 방법
배지 및 시약
미생물분리를위하여
Nutrient broth (NB, Difco, St. Louis, MO, USA)
와Nutrient Agar (NA, Difco)
를사용하였고계대 배양에는NA
를사용하였다.
균주의보존을위하여균배양액에glycerol
이15% (V/V)
가되도록하여-75℃
심온동결고(IIshin DF8514l, Seoul, Korea)
에보관하면서사용하였다.
균주의 분리
기장군연안에서어획한생멸치
(raw anchovy, Engraulis Ja- ponica ) 10 g
을내장을제거하지않고멸균수40 mL
와함께균 질기로파쇄하여5
분간방치한뒤상층액10 mL
을취하여100 mL
의NB
에5 mL
를접종한후50℃, 100 rpm
의진탕배양기(VISION SCI., VS-8480SR, Daejeon, Korea)
에서24
시간동 안배양하였다.
배양액1 mL
을NA
에도말하여50℃ incubator
에서배양하면서발생하는독립적인colony
를분리하여실험 에사용하였다.
α-galactosidase의 활성 측정
위에서 분리한 미생물 배양액 각각
1 mL
를 채취하여12,000×g
에서15
분동안원심분리하고맑은상층액을 취하여
α-galactosidase
의 활성을측정을하였다. α-galactosidase
활성의 측정은 기질인p-nitrophenyl-α-D-galactopyranoside (pNPαGal)
를 사용하여 실시하였다(Food Research Interna- tional, 2009).
즉, 700 μL
의100 mM potassium phosphate buffer (pH 6.5)
에100 μL
의배양액과200 μL
의9.9 mM pNP αGal
을혼합하여5
분간50℃
에서반응시킨후, 2 mL
의0.2 M borate buffer (pH9.8)
를첨가하여반응을중지시키고405 nm
에서흡광도를측정하여α-galactosidase
활성정도를특정하여 활성이높은균주를선발하였다.
미생물 균주의 최적 발육조건 분석
분리된미생물균주의최적발육온도를측정하기위하여
30- 70℃
의온도범위에서각각12
시간동안배양한후균체의농도 를660 nm (Hitachi U-2900, Tokyo, Japan)
에서흡광도로측정 하였으며최적pH
를알아보기위해NB
의pH
를4.0-8.0
으로각 각조정하여50℃
에서12
시간동안배양한후균체의농도를역 시660 nm
에서흡광도로측정하였다.
주사전자현미경을 이용한 균주의 관찰
미생물 균주의 외형 및 크기의 측정은
Scanning Electron Microscopy (SEM, S-2400, Hitachi, JAPAN)
을이용하였다. SEM
측정을위하여고체배지위에세포를고정시키고0.1 M sodium phosphate buffer (pH 7.2-7.4)
로제조한2.5% glutar- aldehyde (TAAB Laboratoties Equipment Ltd, Berks, UK)
용 액으로3
시간고정시킨후0.1 M sodium phosphate buffer
로2
회세척한다음1% osmium tetroxide (Sigma, St Louis, MO, USA)
용액으로30
분간최종고정하였다.
그다음50-100%
농 도구배알코올로각각단계적으로5
분씩탈수시킨후100%
hexamethyldisilazane
로완전히탈수하고건조하였다.
전처리 된샘플은SEM
으로20 kV
의가속전압에서7500
배율로관찰 하였다.
미생물 균주의 생화학적 특성 조사
분리한 미생물균주의 생화학적 특성 검사를위해서
API-
ZYM (BioMèrieux, France)
를 사용하여19
종의효소활성을 조사하였으며α-galactosidase
활성이높게나타난균주를대 상으로API-50CH (BioMèrieux, France)
을사용하여탄소원의 이용패턴을조사하였다.
16S rRNA 유전자 염기서열과 계통수(phylogenetic tree) 분석
미생물균주의동정은
16S rDNA
염기서열의상동성을 분석하여수행되었다
.
즉, 16S rDNA
염기서열을결정하기위해 일반적인방법에따라정제된DNA
를주형으로하여PCR
로 증폭하였다.
증폭된DNA
단편을Big Dye Terminator Cycle Sequencing kit (Applied Biosystems, USA)
로 정제한다음, DNA sequencer (Applied Biosystems model 3730XL, USA)
를 이용하여염기서열을결정하였다.
결정된염기서열을Ez- Taxon server 2.1
을이용하여상동성을분석하였다.
계통수는NCBI
의BLASTN
결과상위50
개를사용하여EMBL-EBI
에 서제공하는ClustalW2
의Neighbor Joining (Saitou and Nei, 1987)
방법을통해phylogenetic tree
를작성하였다.
난소화성 탄수화물 분해능 측정
미생물균주가난소화성탄수화물을분해하는지알아보기위 해서
(Scalabrini et al., 1998) stachyose
와raffinose
를각각10 g/L
씩첨가한NB
에미생물배양액을5% (w/v)
씩각각접종하 여50℃
항온수조에서48
시간동안배양하였다.
배양액1 mL
을12,000×g
에서15
분간원심분리하여얻은상층액250 µL
에 아세토니트릴750 μL
첨가하여같은조건에서다시원심분리하여침전물을제거하고맑은상층액을
0.2 μm
막여과기(membrance filter)
로여과한것을최종분석시료로사용하였 다. Stachyose
와raffinose
의 정량은APS-2 hypersil column
(thermo scientific; 250×4.6 mm, 5 μm, 120A)
이장착된고속액체크로마토그래피를이용하여
(column
온도, 40℃;
이동상, 75% ACN;
유속, 1.5 mL/min)
분석하였다.
검출기는RI
검출 기(refractive index detector, Dionex)
를사용하였다.
미생물 균주가 생산하는 α-galactosidase의 최적 반 응온도 측정
미생물균주가생산하는
α-galactosidase
의최적반응온도를 알아보기위하여미생물균주를50℃
에서의48
시간진탕배양 한다음8,000 rpm
에서20
분간원심분리하여균체와배양액 을분리하였다.
배양액은ammonium sulfate
를35-85%
로포 화되게첨가하고4℃
에서12
시간동안방치하였다.
침전된단 백질은13,000×g
에서20
분간원심분리하여50 mM Tris-HCl (pH7.0)
에용해시킨후동일완충액으로4℃
에서12
시간동안 투석하여조효소액을획득하였다. α-galactosidase
활성측정은 기질로p-nitrophenyl-α-D-galactopyranoside (pNPαGal)
를 사용하여실시하였다(Sigma, St. Louis, MO, USA). 700 μL
의100 mM potassium phosphate buffer (pH 6.5)
에200 μL
의9.9 mM pNPαGal
과100 μL
의조효소액을혼합하여5
분간30-70℃
범위에서각각반응시킨후, 2 mL
의200 mM borate buffer (pH 9.8)
를첨가하여반응을중지시키고405 nm
에서흡 광도를측정하여α-galactosidase
활성을측정하였다.
효소활성1 unit
는1
분에1 μmol
의p-nitrophenol
이효소에의해생성되 는양으로나타내었으며,
활성도(specific activity)
는단백질1 mg
당효소1 unit
로나타내었다.
결과 및 고찰
미생물 균주의 분리 및 α-galactosidase 활성 NA
및NB
배지를이용하여기장일대에서서식하는멸치로부 터고온에서독립적인colony
를나타내는20
개의미생물균주 를분리하였고α-galactosidase
활성을알아보기위하여효소액 에대한기질의분해도를측정하였다.
그결과(Table 1) KM-1
이0.482 Unit/mL
로19
개의다른미생물보다활성이가장높 게나타났으며이는사료제조에있어항영양인자인stachyose
와raffinose
를분해시켜어류의사료섭취시소화력을더욱높 여줄것이라판단된다.
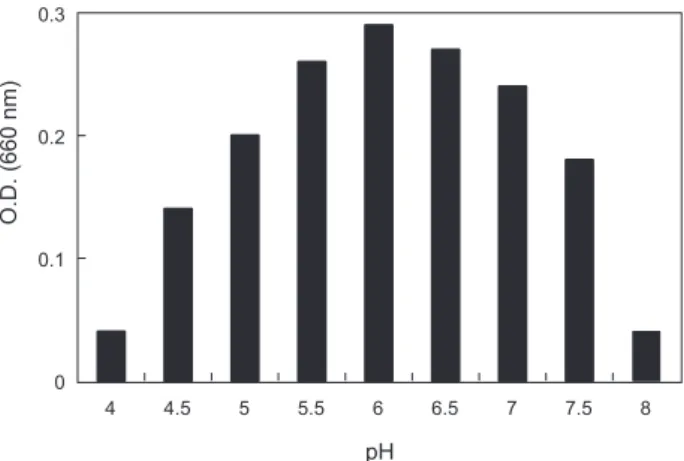
배양시간에 따른 KM-1 균주의 발육조건
α-galactosidase
활성이가장높은KM-1
균주의최적발육조 건을조사한결과는Fig. 1
과같다.
온도는45-55℃
의범위에서 잘자랐으며최적발육온도는50℃
인것으로나타났다. pH
의 경우, pH 5.5-7.0
의범위에서잘자랐으며최적발육pH
는6.0
인것으로나타났다.
이는시료에따라약간의차이를나타내고 있는데pH
변화에따른콩,
메주,
된장의α-galactosidase
의활성(Kim and Yoon, 1998)
연구에서는pH 7.0-7.5
에서가장활성이 높아본연구결과와약간의차이를보였다.
주사전자현미경을 이용한 균주의 관찰
Table 1. α-galactosidase activity of microorganisms isolated from raw anchovy
Isolate Activity (U/mL) Isolate Activity (U/mL)
KM-1 0.482 KM-11 0.154
KM-2 0.230 KM-12 0.161
KM-3 0.128 KM-13 0.143
KM-4 0.153 KM-14 0.112
KM-5 0.133 KM-15 0.113
KM-6 0.112 KM-16 0.135
KM-7 0.154 KM-17 0.164
KM-8 0.151 KM-18 0.160
KM-9 0.152 KM-19 0.135
KM-10 0.132 KM-20 0.157
0 0.1 0.2 0.3
35 40 45 50 55 60
O.D. (660 nm)
Temperature (℃)
0 0.1 0.2 0.3
4 4.5 5 5.5 6 6.5 7 7.5 8
O.D. (660 nm)
pH
0 0.02 0.04 0.06 0.08 0.1
30 40 50 60 70
α-galactosidase activity (Unit/ mL)
Temperature (℃)
Fig. 1. Effect of temperature on growth of Bacillus coagulans KM-1 after 12 hours cultivation.
0 0.1 0.2 0.3
35 40 45 50 55 60
O.D. (660 nm)
Temperature (℃)
0 0.1 0.2 0.3
4 4.5 5 5.5 6 6.5 7 7.5 8
O.D. (660 nm)
pH
0 0.02 0.04 0.06 0.08 0.1
30 40 50 60 70
α-galactosidase activity (Unit/ mL)
Temperature (℃)
Fig. 2. Effect of pH on growth of Bacillus coagulans KM-1 after 12 hours cultivation.
Bacillus coagulans KM-1의 생화학적 특성
519
멸치로부터분리한
20
종의미생물중α-galactosidase
활성이가장높게나타난
KM-1
균주에대한형태학적특성을분석하기위해
KM-1
균주를최적조건에서배양하여그람염색(Gram staining)
과포자염색(Spore staining)
를실시한결과,
포자를형 성하는그람양성균으로나타났으며KM-1
균주를전자현미경 으로관찰한결과(Fig. 3)
세균의지름과길이는각각0.5-0.7 μm, 3.3-4.4 μm
로길이가지름의2
배이상되는장간균(long rod)
인것으로관찰되었다.
또한카탈라제(catalase)
실험결과 양성반응을보여바실러스속(Bacillus sp.)
의형태적특징과일 치하였다.
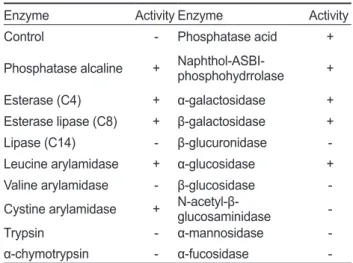
API-ZYM과 API-50CHL Kit를 이용한 균주의 특성
KM-1
균주의 생화학적 특성 분석하기 위해API-ZYM
(BioMèrieux, France)
를 사용하여19
종의 효소활성을 조사 한결과(Table 2) KM-1
균주는alkaline phosphatase, esterase (C4), esterase lipase (C8), leucine arylamidase, cystine aryl- amidase, phosphatase acid, naphthol-ASBI-phosphohydrro- lase, α-galactosidase, β-galactosidase
및α-glucosidase
에대 한 양성반응을 나타내었다.
또한API-50CH (BioMèrieux,
France)
을사용하여50
종의탄소원이용패턴을조사하였는데(Table 3) glycerol, ribose, D-xylose, galactose, D-glucose, D- fructose, D-manose, N-acetyl glucosamine, maltose, melibi- ose, trehalose, starch, D-arabitol
및5-keto-gluconate
를이용하 는것으로나타났다.
16S rRNA 유전자 염기서열 분석과 계통수(phyloge- netic tree)
α-galactosidase
활성이 높은KM-1
균주의16S rDNA
의 염기서열을분석한결과B. coagulans 99.797% (3/1478), B.
acidiproducens 97.171% (40/1414), B. shackletonii 95.831%
Table 2. Enzyme activities of Bacillus coagulans KM-1 by API- ZYM kit
Enzyme Activity Enzyme Activity
Control - Phosphatase acid +
Phosphatase alcaline + Naphthol-ASBI- phosphohydrrolase +
Esterase (C4) + α-galactosidase +
Esterase lipase (C8) + β-galactosidase +
Lipase (C14) - β-glucuronidase -
Leucine arylamidase + α-glucosidase + Valine arylamidase - β-glucosidase - Cystine arylamidase + N-acetyl-β-glucosaminidase -
Trypsin - α-mannosidase -
α-chymotrypsin - α-fucosidase -
+, positive; -, negative.
Fig. 3. Scanning electron microscopy image of Bacillus coagulans KM-1 (×7,500).
Table 3. Carbohydrate utilization pattern of Bacillus coagulans KM-1 by API-50CH kit
Carbohydrate Activity Carbohydrate Activity
Control - Esculin -
Glycerol + Salicin -
Erythritol - Celobiose -
D-arabinose - Maltose +
L-arabinose - Lactose -
Ribose + Melibiose +
D-xylose + Sucrose -
L-xylose - Trehalose +
Adonitol - Inulin -
β-methyl-D-xyloside - Melezitose -
Galactose + D-raffinose -
D-glucose + Starch +
D-fructose + Glycogen -
D-manose + Xylitol -
L-sorbose - β-gentiobiose -
Rhamnose - D-turanose -
Dulcitol - D-lyxose -
Inositol - D-tagatose -
Manitol - L-fucose -
Sorbitol - D-arabitol +
α-methyl-D-manoside - L-arabitol -
α-methyl-D-glucoside - Gluconate -
N-acetyl glucosamine + 2-keto-gluconate -
Amygdalin - 5-keto-gluconate +
Arbutin -
+, positive; -, negative.
남기호
ㆍ
장미순ㆍ
박희연ㆍ
이레나 코네바520
(62/1487)
인것으로나타났다.
따라서신규로분리한KM-1
균 주를B. coagulans KM-1
로명명하고(Fig. 4)
그유전자를미국 국립생물정보센터(NCBI)
에등록(JX569800, 2012. 9. 4)
하였 으며균주를한국미생물종보존센터(KCCM 112989P, 2012. 9.
6)
에기탁하였다.
난소화성 탄수화물 분해능
B. coagulans KM-1
에의한난소화성탄수화물분해시험결 과(Table 4)
발효stachyose
와raffinose
의 함량이 각각0.49 mg/100 g, 0.75 mg/100 g
이었으나24
시간발효후각각0.25 mg/100 g , 0.56 mg/100 g , 48
시간후에는각각0.16 mg/100 g , 0.39 mg/100 g
로지속적으로감소하는결과를나타내었다. Stachyose
와raffinose
의 감소와는 상대적으로saccharose
와galactose
의함량이증가하는것으로볼때B. coagulans KM-1
이 생산하는α-galactosidase
에 의해 난소화성 탄수화물인stachyose
와raffinose
가잘분해된다는것을알수있었다. Kim and Yoon (1998)
은대두의발효에따른α-galactosidase
활성에 관한연구에서발효가진행됨에따라stachyose
와raffinose
함 량이각각감소하는현상을보였으며이는미생물이분해하는α-galactosidase
에의한것이라고보고한바있으며본연구결 과도이와유사함을알수있었다.
미생물 균주가 생산하는 α-galactosidase의 최적 반응온도
B. coagulans KM-1
이 생산하는α-galactosidase
의 반응온 도별효소활성을Fig. 5
에나타내었다. B. coagulans KM-1
이 Fig. 4. Phylogenetic tree of Bacillus coagulans KM-1 based on 16S rDNA sequencing.Table 4. Non-digestible carbohydrate resolution of Bacillus coagu- lans KM-1
Carbohydrate Fermentation time (h)
0 24 48
Raffinose 0.75* 0.56 0.39
Stachyose 0.49 0.25 0.16
Saccharose 0 0.23 0.42
galactose 0 0.33 0.49
*mg/100 g
0 0.1 0.2
35 40 45 50 55 60
O.D. (660 nm)
Temperature (℃)
0 0.1 0.2 0.3
4 4.5 5 5.5 6 6.5 7 7.5 8
O.D. (660 nm)
pH
0 0.02 0.04 0.06 0.08 0.1
30 40 50 60 70
α-galactosidase activity (Unit/ mL)
Temperature (℃)
Fig. 5. Effect of temperature on α-galactosidase activity of Bacillus coagulans KM-1.
Bacillus coagulans KM-1의 생화학적 특성
521
생산하는
α-galactosidase
의활성은55℃-65℃
의범위에서높 은활성을보였으며60℃
에서가장높게나타나B. coagulans KM-1
의최적발육온도인50℃
와는차이를보였다.
따라서분 리된B. coagulans KM-1
으로난소화성탄수화물효율적으로 분해제거하기위해서는먼저50℃
에서충분히미생물을발육 시킨다음60℃
에서효소반응을유도하여야할것으로판단된 다.
일반적으로저온및중온발효는잡균의번식을방지하기위 해살균과정을따로실시하고있으나고온발효는고온에서발 효가이루어지기때문에살균과정을따로하지않아도부패성 미생물의발육이억제된상태에서신속히발효를할수있다는 장점이있다(Jang et al., 2013).
사 사
본연구는국립수산과학원
(
젓갈의속성제조및저염화기술 개발, RP-2014-FS-020)
의지원에의해운영되었습니다.
References
Coombs J and Brenchley. 2001. Characterization of two new glicosyl hydrolases from the lactic acid bacterium Carno- bacterium piscicola strain BA. Appl Environ Microbiol 67, 5094-5099.
Choi C, Choi KS, Lee SH, Hong SP, Lee HD and Bae DK. 1998.
Characteristics and action pattern of α-galactosidase from
Scopulariopsis brevicaulis in Korean traditional Meju. ACB
41, 489-495.Dey PM and Campillo ED. 1984. Biochemistry of the multiple forms of glycosidases in plants. Adv Enzymol Relat Areas Mol Biol 56, 141-249.
Halstead JR, Fransen MP. Eberhart RY, Park AJ, Gilbert HJ and Hazlewood GP. 2000. α-galactosidase a from Pseudomonas
fluorescens subsp. cellulosa: cloning, high level expression
and its role in galactomannan hydrolysis. FEMS Microbiol Lett 192, 197-203.Irish GG, Barbour GW, Classen HL, Tyler RT and Bedford MR. 1995. Removal of the α-galactosidase of sucrose from soybean meal using either ethanol extraction or exogenous alpha-galactosidase and broiler performance. Poult Sci 74, 1484-1494.
Jang MS, Park HY, Nam KH, Han HS, Kim KW, Kim KD and Lee BJ. 2013. Effect of extruded pellets containing fer- mented soybean meal as a partial substitute for fish meal on growth performance and muscle quality of olive flounder (Paralichthys olivaceus). J Agric Life Sci 47, 203-215.
Jindou S, Shuichi K, Emi F, Tsuchiyoshi F, Hidenori H, Tetsuya K, Kazuo S and Kunio O. 2002. α-galactosidase Agar27A, an enzymatic component of the Clostridium josui cellulo- some. J Bacteriol 184, 600-604.
Kim KW, Kim KD, Lee BJ, Lee JH, Han HS, Koo JW, Choi YH and Bai SC. 2013. Dietary fermented soybean meal as a
replacement for fish meal in juvenile olive flounder Parali-
chthys olivaceus. Kor J Fish Aquat Sci 46, 769-776.
Kim HS, Lee KS, Jae HS, Lee MS, Choi JH and Yoon KH.
2004. Characterization of extracellular α-galactosidase pro- duced by Bicillus licheniformis YB-42. Kor J Microbiol Biotechnol 32, 128-134.
Kim JS and Yoon S. 1998. The Changes of α-galactosidase ac- tivities and stachyose and raffinose contents during fermen- tation of soybeans. Korea J Soc Food Sci 14, 509-512.
Li DF, Nelssen JL, Reddy PG, Blecha F, Klemm RD, Giesting DW, Hancock JD, Allee GL and Goodband RD. 1991. Tran- sient hypersensitivity to soybean meal in the early-weaned pig. J Anim Sci 68, 1790-1799.
Min BJ, Kim HI, Hong JW, Moon TH, Lee JH, Han YK, Kwon OS, Lee SH and Lee WB. 2002. Influence of dietary carbo- hydrase on egg quality and nutrient digestibility in laying hens. Korea J Poult Sci 29, 19-23.
Rackis JJ. 1981. Flatulence caused by soya and its control through processing. J Am Oil Chem Soc 58, 503-510.
Reddy NR, Salunkhe DK and Aharma RP. 1980. Flatulence in rats following ingestion of cooked and germinated black and a fermented product of black gram and rice blend. J Food Sci 45, 1161.
Silvestroni A, Connes C, Sesma F, Giori GS and Piard JC. 2002.
Characterization of the mel locus for α-galactosidase in
Lactobacillus plantarum. Appl Environ Microbiol 68, 5464-
5471.Scalabrini, Rossi P, Spettoli M, Matteuzzi P. 1998. Characteriza- tion of Bifidobacterium strains for use in soymilk fermenta- tion. Int J food microbiol 39, 213-219.
Saitou N and Nei M. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4, 406-425.
Steggerda FR, Richards EA and Rackis JJ. 1968. Effects of vari- ous soybean products on flatulence in the adult man. Proc Soc Exp Biol Med 121, 1235.
Sanada CTN, Karp SG, Spier MR, Portella AC, Gouvea PM, Yamaguishi CT, Vandenberghe LPS, Pandey A and Soccol CR. 2009. Utilization of soybean vinasse for α-galactosidase production. Food Res Int 42, 476-483. http://dx.doi.
org/10.1016/j.foodres.2009.01.017.
Yoo JS, Cho JH, Chen YG, Kim HJ, Wang Q, Hyun Y, Ko TG, Park CS and Kim IH. 2007. The Effects of environment- friendly diets on the growth performance, nutrient digestibil- ity, fecal excretion, nitrogen excretion and emission gases in manure for growing pigs. J Anim Sci Technol 49, 491-500.