38
Copyright © 2016 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
유글레나
(euglenoids)
는직경15-500 µm
크기의나사선모양 으로꼬인길쭉한방추형인데매우자유롭게변형하며,
체내에 엽록체를가지고광합성을하는점으로보아식물에속한다고 하지만,
세포벽이없고편모로유영생활을하므로원생동물의 편모충류로취급하기도한다(Rodríguez-Zavala et al., 2010).
즉
,
식물과동물의중간에위치한다고볼수있는데엽록소를가 지고광합성을하는것은식물적특성이며,
입이나수축포를가 지고자유롭게움직이는것은동물적특성에속한다(Ruiz et al.,
2004).
대부분담수에서살며작은연못이나도랑에서흔히볼수있고내만의염수역에서도볼수있다
.
현재,
유글레나를함 유하는조류에의한유용물질생산의가능성에관해서각종산업계가주목하고있으며
(Hayashi et al., 1994; Chisti and Yan,
2011),
유글레나분말은식품으로서의실용화가이미수행되고있으며
(Choi et al., 2004),
고등식물에비하여석유화학대체원 료,
혹은연료의원료등유용물질을고수율로서생산할수있 는능력을가지고있다(Ishikawa et al., 2008; Lira-Silva et al.,
2011).
또한수많은미세조류중에서도여러분야에서유용성이기대되고있는유글레나는광합성을통해대기중이산화탄 소를섭취해고정하고산소를배출하므로이를이용한하폐수 의영양염류제거및회수된바이오매스를
Bio-fuel
로생산하 는시스템개발을위한연구가진행되고있다(Navarro et al., 1997; Ramalho et al., 1998; Tucci et al., 2006; Jasso-Cha´
vez et al., 2010).
현재유글레나의배양방법으로는자가영양 배양방법과타가영양 배양방법으로이루어지고있으나,
대부배양방법에 따른 Euglena gracilis의 성장 및 지방산 조성
정우철·최종국·강창민
1·최병대·강석중*
경상대학교 해양식품생명의학과, 1안전성평가연구소 경남환경독성본부
Effects of Culture Methods on the Growth Rates and Fatty Acid Profiles of Euglena gracilis
U-Cheol Jeong, Jong-Kuk Choi, Chang-Min Kang
1
, Byeong-Dae Choi and Seok-Joong Kang*Department of Seafood and Aquaculture Science, Gyeongsang National University, Tongyeong 53064, Korea
1
Institute of Toxicology, Jinju 52834, Korea
The quality and quantity of live food sources strongly influence the success of fish production in farming operations.
Thus, critical studies of live forage species are a crucial element for progress in fish aquaculture. The fat content of food is an especially important determinant of growth in marine fish. Omega-3 highly unsaturated fatty acids (HUFA) are essential components of diet that determine the nutritional value of larval fish. Euglena is a protist that has poten- tial as a forage species. These single-celled organisms have plant and animal characteristics they are motile, elliptical in shape and 15–500 µm in diameter. Their nutritional content is excellent, but most studies have focused on cells raised in autotrophic culture. We therefore examined differences in the lipid and fatty acid contents, and the growth of Euglena cells grown under autotrophic, heterotrophic, and mixotrophic conditions. Biomass production reached 15.03 g/L, 12.28 g/L, and 3.66 g/L under mixotrophy, heterotrophy, and autotrophy, respectively. The proportional n-3 HUFA content differed among culture methods: 10.04%, 5.80% and 10.01% in mixotrophic, heterotrophic and autotrophic cultures, respectively. Mixotrophy was to be the best form of cultivation for improving the growth and nutritional content of Euglena .
Key words: Euglena gracilis, Fatty acid, Heterotrophic, Autotrophic, Mixotrophic
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2016.0038 Korean J Fish Aquat Sci 49(1) 038-044, February 2016
Received 16 January 2016; Revised 16 February 2016; Accepted 17 February 2016
*Corresponding author: Tel: +82. 55. 772. 9154 Fax: +82. 55. 644. 4202
E-mail address: [email protected]
분의배양방법은자가영양
(autotrophic)
배양방법이주를이루 고있다(Chae et al., 2006).
타가영양배양방법은제한적이기 는하지만,
유글레나는자가영양배양과타가영양배양이모두 가능한것으로알려져있다(Dos et al., 2007).
타가영양배양방 법은자가영양배양방법과달리광조사가필요하지않으며,
성 장률이높다는장점이있다(Regnault et al., 1995).
유글레나는 여러가지기능성물질을생산하는것으로알려져있으며, β-1,3 glucan
으로 구성된 파라밀론(paramylon)
은 천연플라스틱을 만드는원료로도사용되어지고있다(Barsanti et al., 2000ab;
Choi et al., 2013). Vitamin E
를포함한영양소가높기때문 에식품첨가물로도사용되고있으며,
지질함량이15-24% (dry
weight)
로높기때문에바이오연료생산에관한연구도이루어지고있다
(Navarro et al., 1997).
유글레나자체의바이오매스(Biomass)
는일반적으로식품(Food),
섬유(Fiber),
사료(Feed),
비료(Fertilizer),
연료(Fuel)
의 순서로 부가가치가 높다(Roc- chetta et al., 2006; Courchesne et al., 2009). E. gracilis
는세 포건조량의20%
이상이면역증강제인β-1,3 glucan,
고함유 지질(15-24%),
다른미세녹조류에는볼수없는EPA, DHA,
등을함유하고있기때문에어류자치어의먹이생물도관심을 받고있다(Harwood, 1988; Hayashi et al., 1994; James and Browse, 1999; Choi et al., 2013).
또한어류자치어의먹이생 물인로티퍼와알테미아의영양강화제로도사용되어지고있다(Hayashi et al., 1992).
이처럼수산용먹이생물로서각광을받 고있는유글레나(E. gracilis)
가산업적으로유용성을갖기위 해서는어류자치어가요구하는기능성물질은높게유지하면 서경제성있는배양방법의개발이절실하다.
따라서이번연구 에서는그기초실험의일환으로배양방법에따른유글레나의 성장과지질영양학적인변화를조사하고자하였다.
재료 및 방법
실험 주 및 배지
본실험에사용된유글레나는한국해양미세조류은행으로부 터
Euglena gracilis KMMCC-1351
를분양받아사용하였으 며,
배양에사용한기본배지는Table 1
에나타낸바와같이Cra- mer-Myers medium (Cramer and Myers, 1952)
을사용하였다.
타가영양배양과혼합영양배양은Cramer-Myers medium
를기 본배지로사용하여Glucose 15 g/L, Sodium glutamate 5 g/L
추가하여제조하였으며,
각각의배지별로제조한후121℃
에서15
분간고압멸균하였다.
배양방법
배양방법은자가영양배양
(Autotrophic culture),
타가영양배 양(Heterotrophic culture)
그리고혼합영양배양(Mixotrophic culture)
으로하였다.
자가영양배양방법은1 L
둥근플라스크에500 mL
용량으로배양하였으며,
광주기는18L:6D,
주간의조도는
3,000 lx
로유지하였고, Air
는0.20 µm sterilized air filter (Advantec mfs, Inc., Japan)
를통해공급하였다.
타가영양배양 방법은500 mL
삼각플라스크를이용하여200 mL
용량으로배 양하였으며,
광주기는0L:24D
로하였으며,
교반은진탕배양기(KSI-200L, Koencon Co., Ltd)
를이용하여120 rpm
으로하 였다.
혼합영양배양방법은500 mL
삼각플라스크를이용하여200 mL
용량으로배양하였으며,
광주기는18L:6D,
주간의조 도는3,000 lx
로유지하였다.
이때모든실험구의배양온도는25±0.5℃
로유지하였다.
초기접종밀도는10×10
4cells/mL
로7
일간3
반복으로배양하였고, 2
회/
일동일한시간에Haema-
cytometer
로3
회반복계수하여평균세포수를측정계수하였으며
,
배양이완료된배양액은4℃
의고속원심분리기(UNION 32R, Hanil Science Industrial Co., Ltd. Korea)
에서3,000 rpm
으로25
분간원심분리한후,
배양액은버리고세포만회수하여 동결건조하여분석에사용하였다.
총지질
총지질추출은
Bligh and Dyer
방법(1959)
에준하였다.
비커 에 균체5 g
을 취하여세포분쇄기(homogenizer AM-12, Ni- honseiki Kaisha Co. Ltd., Tokyo, Japan)
에서15,000 rpm
로5
분간분쇄한후, Chloroform
과Methanol
을2:1
로혼합한추 출용매를시료의2
배량넣어하루동안방치한다음chloro-
form
층만을분리하기위하여둥근플라스크위에깔때기를놓고
,
그위에Na
2SO
4를넣어서서히chloroform
층만흘러내리게 하였다.
분리된chloroform
층은 진공회전농축기(Rotavapor R-114, BUCHI)
를사용하여40℃
이하에서용매를완전히증 Table 1. Composition of Cramer-Myers (C&M)1 mediumComponents Concentration
KH2PO4 1.0 g
(NH4)2SO4 1.0 g
MgSO4·7H2O 0.2 g
CaCl2·2H2O 0.2 g
EDTA-3Na·2H2O 0.8 g
Fe(SO4)2·6H2O 3.0 mg
MnCl2·4H2O 1.8 mg
CoSO4·7H20 1.5 mg
ZnSO4·7H2O 0.4 mg
Na2MoO4·2H2O 0.2 mg
CuSO4·5H2O 0.02 mg
Vitamin B1 0.1 mg
Vitamin B12 0.5 µg
Distilled water final volume 1 L
Final pH pH 3.5
1Cramer-Myers (C&M) medium (Cramer and Myers, 1952).
발시킨후
,
추출된총지질의무게를측정하였다.
모든작업은 질소기류하에서행하였다.
지질
지질의
classes
는TLC/FID
와Iatrorecorder TC-21 intergra- tor
가 장착된Iatroscan New MK-5 (Iatron Laboratory Inc., Tokyo, Japan)
를이용하여 분석하였다.
분석조건에서공기의 유속은2 L/min
이며detector
수소량은160 mL/min
으로, scan- ning speed
는0.30 cm/s
로하였다.
분석과정은먼저Rod S-Ⅲ (0.9×150 mm,
석영봉규산코팅)
를5
분간수세한후,
다시증류수
10 mL
로헹군다음수분을증발시키기위하여아세톤10
mL
로씻고50℃
로조정한Rod-Dryer (TK-5 Iatron Lab. Inc., Tokyo, Japan)
에서5
분간건조시킨후Iatroscan
내에서수소염 이온화불꽃상에서3
회이상반복하여유기물을완전히제거시 켰다. Rod
에시료1 μL
를Microdispenser (Drummond Scien- tific Co., Bromall, PA, USA)
로써점적하여전개조(NaCl
로 포화시킴)
에서10
분간포화시켰다.
전개용매는n-hexane : di- ethyl ether : acetic acid = 97:3:1 (v/v)
를이용하여약10 cm
까 지전개시킨후, Rod
를전개조에서꺼내고Rod-Dryer
에서5
분 간건조시켜서Iatroscan
으로분석하여지질획분의조성비를 구하고그함량을산출하였다.
동정은표품인cholesterol ester, free fatty acid, triglyceride, cholesterol
및phospholipid
에의 하여동정하였다.
지방산
지방산
methyl ester
유도체化
는시료일정량과내부표준물 질(C
23:0methyl ester) 1 mL (1 mg)
를cap tube
에취하고, 0.5 N NaOH-methanol
용액1.5 mL
를가하여질소를충진한다 음, 100℃
에서8
분간가열하여검화하였다.
방냉후12% BF
3- methanol 2 mL
를가한후tube
의뚜껑을닫고, 100℃
에서11
분 간가열하여methyl
화하였다.
약30℃
로냉각한후Iso-octane 1 mL
를첨가하고30
초간vortex mixer
로혼합하였다.
즉시3 mL
의포화식염수를가한다음흔들어방치하여iso-octane
층 이분리되도록하였다. Iso-octane
층을시료병(4 mL)
에옮긴 후,
다시Iso-octane 1 mL
를첨가한다음흔들어재추출하여 시료병에모으고이를지방산methyl ester
시료로하였다.
지 방산분석에사용하는GLC
는Omegawax
TM-320 fused-silica capillary column (30 m×0.32 mm×0.25 µm, i.d., Supelco Co., Bellefonte, PA, USA)
를장착한Clarus 600 (Perkin Elmer Co. Ltd., USA)
를이용하였다.
분석조건으로Column
은185℃
에서
8
분간유지하고3℃/min
씩230℃
까지상승시킨후, 10
분 간유지하였다.
이때주입기는250℃,
검출기는270℃
그리고carrier gas
는He (1.0 kg/cm
2)
을사용하였다.
지방산의분석은 동일조건에서분석한표준품의ECL
과비교하여동정하였고,
지방산표준품은14:0, 16:0, 18:1, 18:2, 18:3, 20:0, 22:1, 24:0 (Sigma Chemical Co., St. Louis, MO, USA)
과GC-MS
로동 정된menhaden oil
을사용하였다.
통계처리
모든 자료는
SPSS (16.0)
프로그램을 이용하여 분산분석(one-way ANOVA)
과회귀분석(Regression Analysis)
을실시 하여Duncan's multiple range test (Duncan, 1955)
로평균간의 유의성(P<0.05)
을검정하였다.
결과 및 고찰
배양방법에 따른 성장
E. gracilis
를 이용하여 자가영양배양,
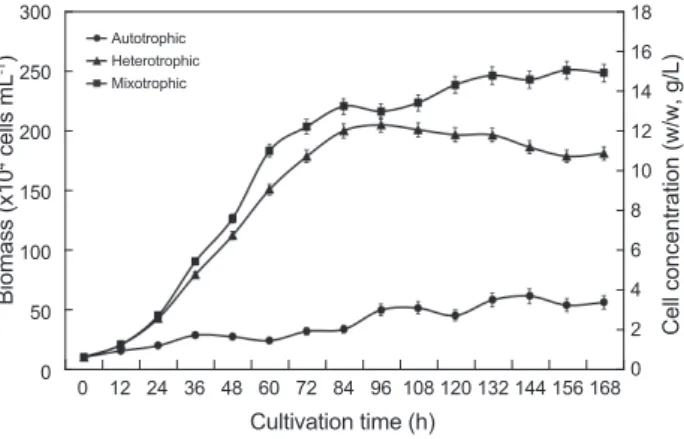
타가영양배양 그리 고혼합영양배양방법에따른결과는Fig. 1
에나타낸바와같 다.
배양방법에따른세포의성장실험은실험구마다동일하게10×10
4cells/mL
로접종하였다.
자가영양배양방법을통한배 양결과는초기배양밀도10.2×10
4cell/mL
로접종한후12
시35 30 25 20 15 10 5 0
Fatty acid
%, Total fatty acid composition
C14:0 C16:0 C18:2n-6 C18:3n-3 C20:5n-3 C22:6n-3 Autotrophic Heterotrophic Mixotrophic Autotrophic
Heterotrophic Mixotrophic 300
250 200 150 100 50 0
18 16 14 12 10 8 6 4 2 0
Cultivation time (h)
Biomass (x10
4cells mL
-1) Cell concentration (w/w, g/L)
0 12 24 36 48 60 72 84 96 108 120 132 144 156 168
Fig. 2. Fatty acid profile of Euglena gracilis grown in different cul- ture conditions. Data were means±S.D. of triplicate.
35 30 25 20 15 10 5 0
Fatty acid
%, Total fatty acid composition
C14:0 C16:0 C18:2n-6 C18:3n-3 C20:5n-3 C22:6n-3 Autotrophic Heterotrophic Mixotrophic Autotrophic
Heterotrophic Mixotrophic 300
250 200 150 100 50 0
18 16 14 12 10 8 6 4 2 0
Cultivation time (h)
Biomass (x10
4cells mL
-1) Cell concentration (w/w, g/L)
0 12 24 36 48 60 72 84 96 108 120 132 144 156 168
Fig. 1. The effect of cultivation methods on the cell growth of Eu- glena gracilis. Data were means±S.D. of triplicate.
간간격으로성장을측정하였다
.
그결과접종이후24
시간경 과19.8×10
4cell/mL, 48
시간경과27.3×10
4cell/mL, 72
시간 경과31.8×10
4cell/mL, 96
시간경과49.7×10
4cell/mL, 120
시간경과45.0×10
4cell/mL, 144
시간경과61.3×10
4cell/
mL
로증가하였으며그이후감소하는경향을나타냈다.
이로 서자가영양배양에의한성장은접종후144
시간째61.3×10
4cell/mL
로최대성장을나타냈으며,
이때바이오매스는4.0 g/
L
였다.
타가영양배양방법을통한배양결과는초기배양밀도10.2×10
4cell/mL
로접종한후12
시간간격으로성장을측정 하였다.
최초접종후24
시간경과42.7×10
4cell/mL, 48
시간 경과112.0×10
4cell/mL, 72
시간경과178.3×10
4cell/mL, 96
시간경과204.6×10
4cell/mL, 120
시간경과196.4×10
4cell/
mL, 144
시간경과186.0×10
4cell/mL
였으며그리고168
시간 경과시180.8×10
4cell/mL
였다.
타가영양배양에의한성장은 접종후96
시간경과204.6×10
4cell/mL
일때가장높은성장 을보였으며,
이때바이오매스는12.0 g/L
였으며,
이후점차감 소하기시작하였다(P<0.05).
이러한경향은초기투여된배지 가모두소진되어더이상성장을할수없는배양조건이된것 으로보여짐에따라타가영양배양의경우배양시작후84
시간 전후로추가배지공급이이루어져야지속적인성장이이루어 질것으로사료된다.
혼합영양배양방법으로배양한결과는초 기배양밀도10.2×10
4cell/mL
로접종한후, 12
시간간격으 로성장측정을하였다.
그결과접종후24
시간경과45.0×10
4cell/mL, 48
시간 경과126.3×10
4cell/mL, 72
시간 경과203.2×10
4cell/mL, 96
시간경과216.0×10
4cell/mL, 120
시 간경과238.2×10
4cell/mL, 144
시간경과242.4×10
4cell/
mL
였으며그리고168
시간경과후248.2×10
4cell/mL
였다.
혼합영양배양에의한성장은접종후156
시간후에250.6×10
4cell/mL
로이때가장높은성장을보였으며(P<0.05),
이때바 이오매스는15.0 g/L
였다.
배양방법에따른세포의성장실험 은실험구마다동일하게10×10
4cells/mL
로접종하였다.
자가 영양배양구에서는접종후서서히증가하는경향을보이며,
접 종후144
시간경과시점에서61.3×10
4cells/mL
로가장높게 나타났지만(P<0.05).
타가영양배양과혼합영양배양에서는접 종후급격히증가하는 경향을보이며,
타가영양배양의경우 는접종후84
시간째199.8×10
4cells/mL
로급격한증가를보 였으며이후증가속도가낮아지며96
시간째204.6×10
4cells/
mL
로가장높게나타났다(P<0.05).
혼합영양배양의경우는접 종후84
시간째220.2×10
4cells/mL
까지급격한증가를보였으 나,
이후증가속도가낮아지며168
시간배양후에250.2×10
4cells/mL
로가장높은성장을나타내었다(P<0.05).
이로써배 양방법에따른E. gracilis
의배양결과는혼합영양배양법을통 한배양이가장높은성장을나타냈으며,
다음으로타가영양배 양법그리고자가영양배양법순으로나타났다.
이로써성장측 면에서E. gracilis
의생산을위한가장적합한방법은혼합영양 배양법인것으로나타났다.
총 지질 및 지질 종류의 변화
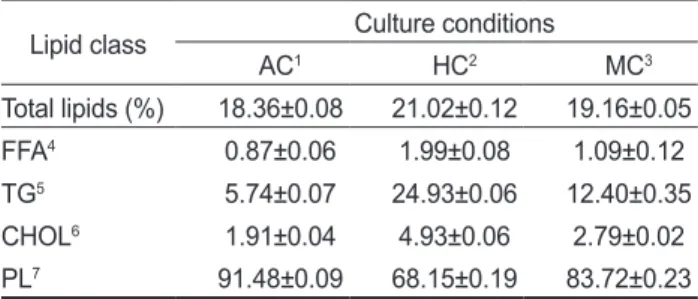
배양방법에따른총지질의변화는
Table 2
에나타낸바와같 이자가영양배양방법18.36%,
타가영양배양방법21.02%,
혼 합영양배양19.16%
으로타가영양배양방법이자가영양배양방 법에비하여총지질의함량이높은것으로나타났다(P<0.05).
배양방법에따른
E. gracilis
의지질class
조성은Table 2
에나 타낸바와같다.
극성지질과비극성지질의조성을보면극성지 질이모든실험구에서68.15-91.48%
로지질조성의대부분을 차지하였다.
특히자가영양배양방법을통하여배양하였을때91.48%
로가장높게나타났으며,
그다음이혼합영양배양법을통한배양에서
83.72%
였으며,
타가영양배양법을통한배양의경우에
68.15%
로가장낮게나타났다(P<0.05).
자가영양배양 방법이타가영양배양방법에비하여극성지질이뚜렷하게증가 하는이유는광합성의유무에의한차이로생각된다.
이러한경 향은El-Sheekh and Fathy (2009)
의연구결과에서Chlorella
vulgaris
를자가영양배양과타가영양배양의조건에서배양하였을때자가영양배양방법의
chlorophyll
함량이타가영양배양 방법에비하여2
배증가한다는결과와거의일치하고있다.
따 라서본실험에서의극성지질의현저한차이는chlorophyll
색 소,
인지질,
당지질의증가에기인할가능성이높으므로향후이 에대한자세한연구가요망된다.
비극성지질의조성에서는모든실험구에서
triglycerols (TG)
가가장높게나타났으며그정도는타가영양배양밥법24.93%,
혼합영양배양방법12.40%,
자가영양배양방법5.74%
의순으 로 나타났다(P<0.05).
이는 극성지질과 역관계를 나타냈다. TG
는에너지원인저장지질로서타가영양방법의배지의주성 분인포도당이저장지질로서의전환가능성을나타낸다. Cho- lesteryl ester (CHOL)
의함량은자가영양배양법을통하여배 양하였을때1.91%
로가장낮게나타났으며(P<0.05),
혼합영 양배양법을통한배양에서2.79%
였으며,
타가영양배양법을통Table 2. Lipid classes in cells cultivated under different culture conditions
Lipid class Culture conditions
AC1 HC2 MC3
Total lipids (%) 18.36±0.08 21.02±0.12 19.16±0.05
FFA4 0.87±0.06 1.99±0.08 1.09±0.12
TG5 5.74±0.07 24.93±0.06 12.40±0.35
CHOL6 1.91±0.04 4.93±0.06 2.79±0.02
PL7 91.48±0.09 68.15±0.19 83.72±0.23
1AC: Autotrophic culture, 2HC: Heterotrophic culture, 3MC:
Mixitrophic culture, 4FFA: free fatty acids, 5TG: triacylglycer- ols, 6CHOL: cholesteryl ester, 7PL: polar lipids and chlorophylls.
The values are mean±S.D. (n=3). Different superscript letters within rows represent significant differences between treatments (P<0.05).
한배양의경우에
4.93%
로가장높게나타났다(P<0.05).
배양 방법에따라서는TG
와같은경향을나타냈다. Free fatty acids
(FFA)
함량은자가영양배양법을통하여배양하였을때0.87%
로가장낮게나타났으며
(P<0.05),
혼합영양배양법을통한배양에서
1.09%
였으며,
타가영양배양법을통한배양의경우에1.99%
로가장높게나타났다(P<0.05).
유글레나의바이오매 스의획득이목적이아니라,
색소와극성지질과같은기능성물 질을중점적으로얻기위한배양방법은광조건에서배양하는 자가영양배양방법이유효한것으로나타났다.
지방산 변화
배양방법에따른
E. gracilis
의지방산조성은Table 3
에나타 낸바와 같다.
자가영양배양,
타가영양배양 그리고혼합영양 배양방법을통해서배양된결과는포화지방산(Saturated fatty
acid)
의경우자가영양배양,
타가영양배양그리고혼합영양배양에서각각
51.54%, 70.09%
그리고34.78%
로타가영양배양 을할경우포화지방산이가장높았으며,
자가영양배양그리고 혼합영양배양순으로나타났다(P<0.05).
포화지방산의종류에 있어서도자가영양배양,
타가영양배양그리고혼합영양배양방 법에서공통적으로16:0, 18:0
가주요지방산을차지하였다.
그 러나그함량에는배양방법에따라조성비차이가큰것으로 나타났다.
불포화지방산(unsaturated fatty acid)
의경우는각각48.46%, 29.91%
그리고65.22%
로혼합영양배양을할경우불 포화지방산이가장높았으며,
자가영양배양그리고타가영양배 양순으로나타났다(P<0.05). 18:3n-3
의경우자가영양배양그 리고혼합영양배양에서각각14.92%
그리고12.30%
로나타났 으나,
타가영양배양에서는0.73%
로자가영양배양과혼합영양 배양한결과와비교해서낮게나타났으며(P<0.05). 18:2n-6
의 경우자가영양배양그리고혼합영양배양에서각각8.23%
그리 고7.06%
로나타났으나,
타가영양배양에서는0.42%
로자가 영양배양과혼합영양배양한결과와비교해서낮게나타났다(P<0.05). 20:5n-3(EPA)
의경우자가영양배양,
타가영양배양 그리고혼합영양배양에서각각4.79%, 2.22%
그리고3.88%
로자가영양배양에서가장높았으며
,
혼합영양배양그리고타 가영양배양순으로나타났으며(P<0.05), 22:6n-3(DHA)
의경 우는각각1.74%, 1.56%
그리고2.47%
로혼합영양배양을할 경우가장높았으며(P<0.05),
자가영양배양그리고타가영양배 양순으로나타났다(P<0.05).
그리고∑n-3 HUFA (highly un- saturated fatty acid)
의경우자가영양배양,
타가영양배양그리 고혼합영양배양에서각각10.01%, 5.80%
그리고10.04%
로 자가영양배양과혼합영양배양을할경우가높았으며(P<0.05),
타가영양배양에서는다른두실험구에비해서낮게나타났으 며(P<0.05), ∑n-6 HUFA
의경우는각각8.88%, 8.10%
그리고10.13%
로혼합영양배양에서가장높았으며(P<0.05),
자가영 양배양그리고타가영양배양순으로나타났다(P<0.05).
이상의결과를종합해보면유글레나의지방산조성은배양방
Table 3. Fatty acid compositions of Euglena gracilis grown in dif- ferent culture conditions (% of total fatty acids) Fatty acid Culture Conditions
AC1 HC2 MC3
14:0 12.30±0.20 31.15±0.18 8.90±0.06
15:0 2.28±0.08 17.78±0.14 8.68±0.03
Iso16:0 5.81±0.06 8.93±0.08 0.90±0.01
16:0 27.65±0.18 6.15±0.06 10.25±0.12
16:1n-5 3.08±0.02 3.54±0.03 4.26±0.06 16:2n-4 0.47±0.01 2.00±0.04 1.36±0.04
17:0 0.90±0.01 4.09±0.02 4.37±0.02
16:3n-4 1.26±0.03 3.05±0.01 9.00±0.08 16:3n-1 0.00±0.00 0.12±0.01 0.45±0.01 16:4n-1 0.00±0.00 0.35±0.00 1.54±0.01
18:0 2.45±0.02 1.44±0.03 1.45±0.00
18:1n-9 1.35±0.01 0.79±0.02 3.33±0.03 18:1n-7 0.00±0.00 0.14±0.00 0.71±0.02 18:2n-6 8.23±0.06 0.42±0.01 7.06±0.10 18:2n-4 0.25±0.00 0.15±0.00 1.23±0.08 18:3n-3 14.92±0.10 0.73±0.00 12.30±0.16 18:4n-1 0.00±0.00 1.65±0.03 1.41±0.02
20:0 0.16±0.01 0.54±0.00 0.23±0.01
20:1n-7 0.00±0.00 2.10±0.06 1.43±0.00 20:2n-6 2.55±0.03 1.19±0.02 1.71±0.01 20:3n-6 0.61±0.00 0.69±0.00 0.61±0.00 20:4n-6 3.39±0.03 3.39±0.10 3.64±0.03 20:3n-3 1.38±0.02 0.50±0.00 0.77±0.01 20:4n-3 1.77±0.06 0.74±0.00 1.54±0.02 20:5n-3 4.79±0.08 2.22±0.08 3.88±0.05 22:1n-7 0.00±0.00 0.95±0.02 0.98±0.00 22:4n-6 0.23±0.01 0.99±0.01 1.31±0.03 22:5n-6 2.11±0.06 1.84±0.06 2.87±0.08 22:4n-3 0.05±0.00 0.18±0.00 0.17±0.00 22:5n-3 0.28±0.01 0.61±0.01 1.21±0.03 22:6n-3 1.74±0.02 1.56±0.06 2.47±0.04
SFA4 51.54 70.09 34.78
USFA5 48.46 29.91 65.22
n-3 HUFA6 10.01 5.80 10.04
n-6 HUFA 8.88 8.10 10.13
n-6/n-3 0.89 1.40 1.01
1AC: Autotrophic culture, 2HC: Heterotrophic culture, 3MC: Mixi- trophic culture, 4SFA: Saturated fatty acid, 5USFA: Unsaturated fatty acid, 6HUFA: Highly unsaturated fatty acid. The values are mean±S.D. (n=3). a-fDifferent superscript letters within rows repre- sent significant differences between treatments (P<0.05).
법에따라서현저히달라지는특징을나타낸다
.
현재미세조류의분류
biomarker
로서지방산이주요한도구로사용되고있으며
(Volkman et al., 1998),
유글레나조류는엽록체의유무를기 준으로독립영양군과종속영양군으로구분된다.
녹색유글레 나류는엽록체와안점(stigma)
이있고,
광합성을하며,
무색유 글레나는오랜진화적역사를통해엽록체를잃은것으로추정 된다고하였으나(Walne 1980),
이번실험을통해서종에따른 차이보다광조사의유무에따라극성지질이현저히변동하므 로극성지질의많은부분을차지하고있는엽록체의생성의차 이로종분류하는것은문제점이있는것으로사료된다.
이처럼 배양방법과환경조건에따라지질생합성형태가달라지므로(Coleman et al., 1988; Girotti 2001),
유글레나를냉암소에서starvation
시키면서지속적으로관찰한결과분류학적으로중요한엽록체나파라밀론의형태적인변이가일어나는점에서
(Kim and Boo, 1998; Watanabe and Suzuki, 2002)
향후여기 에대한세밀한연구가요구된다.
배양방법에따른지방산의변 동은포화지방산의경우자가영양배양방법보다타가영양배양방법에서높은함량을나타내는반면에
∑n-3 HUFA
의경우에는타가영양배양보다자가영양배양과혼합영양배양에서높게 나타났다
(P<0.05).
또한식물류에서특이적으로높은18:2n-6
은자가영양배양과혼합영양배양에서높게나타났다.
이러한 원인은광의의한영향이큰것으로사료된다.
유글레나는엽록 소를가지고있어광합성을하는식물성특성과입이나수축포 를가지고자유롭게움직이는동물성특성모두를가지고있어 광의유무와유기탄소원의공급에따른성장특성이다르게나 타났는데자가영양배양에서는광합성을통한식물성특성의성 장을보였지만,
타가영양배양에서는공급된유기탄소원을흡 수하여성장을하는동물성특성을보였다.
혼합영양배양방법 에서는식물성과동물성특성을모두보이며성장한것으로사 료된다.
또한배양온도는미세조류에의해생산된지방산의종류에중요한영향을미치는것으로보고되고있으며
(Thomp-
son et al.,1992),
일반적으로많은미세조류종들은배양온도가 감소하였을때불포화지방산의비율이증가하는것으로알려져 있다(Jiang and Chen, 2000).
또한유기탄소원의농도가낮으 면지질의축척이촉진하였는데이것은성장률이낮았기때문 이라고보고되고있으며(Jiang and chen, 1992),
배지의영양소 중에서질소성분이부족할경우에지질축적이된다고알려져 있다(Honda et al., 1998).
이번실험을통해서배양방법에따른 유글레나의성장과오메가-3
고도불포화지방산과같은기능성 물질과지방산의함량은달라지는것으로나타났다.
유글레나 의성장측면에서는유기탄소원을사용하여배양하는것이안 정적인바이오매스를확보할수있는것으로나타났으며,
어류 자치어의먹이생물로서지질영양학적인측면에서는광을이용 한배양방법이우수한것으로나타나양측을고려해보면혼합 영양배양을하는것이가장좋은것으로나타났다.
사 사
이논문은안전성평가연구소에서
2015
년도(
바이오화학소 재용바이오매스의개발을위한유글레나대랑배양기술개발)
으로수행하는연구사업의위탁연구임.
References
Barsanti LR, Bastianini A, Passarelli V, Tredici MR and Gual- tieri P. 2000a. Fatty acid content in wild type and WZSL mutant of Euglena gracilis. J Appl Phycol 12, 515-520.
Barsanti LR, Vismara R, Passarelli V and Gualtieri P. 2000b.
Paramylon(b-1,3- glucan) contentin wild type and WZSL mutant of Euglena gracilis. Effectsof growth conditions. J Appl Phycol 13, 59-65.
Bligh EG and Dyer WJ. 1959. A rapid method of total lipid ex- traction and purification. Can J Biochem Physiol 37, 911- Chae SR, Hwang EJ and Shin HS. 2006. Single cell protein pro-917.
duction of Euglena Gracilis and carbondioxide fixation in an innovative photo-bioractor. Bioresour Technol 97, 322- Choi JA, Oh TH, Choi JS, Chang DJ and Joo CK. 2013. Impact 329.
of beta-1,3-glucanisolated from Euglena gracilis on corneal epithelial cell migration and on wound healing in a rat alkali burn model. Curr Eye Res 38, 1207-1213.Chisti Y and Yan J. 2011. Energy from algae: current status and future trends algal biofuels a status report. Appl Energy 88, 3277-3279.
Choi SW, Park IK and Park BS. 2004. Effect of dietary supple- mentation of fresh water algae euglena on the performance and egg quality and fatty acid composition of egg yolk in laying hens. Korean J Poult Sci 31, 283-291.
Coleman LW, Rosen BH and Schwartzbach SD. 1988. Envi- ronmental-control of carbohydrate and lipid-synthesis in Euglena. Plant Cell Physiol 29, 423-432.
Courchesne NMD, Parisien A, Wang B and Lan CQ. 2009. En- hancement of lipid production using biochemical, genetic and transcription factor engineering approaches. J Biotech- nol 141, 31-41.
Cramer M and Myers J. 1952. Growth and photosynthetic char- acteristics of Euglenagracilis. Arch Microbiol 17, 384-402.
Dos SFV, Rocchetta I, Conforti V, Bench S, Feldman R and Levin M. 2007. Gene expression patterns in Euglena graci-
lis: insights into the cellular response to environmental
stress. Gene 389, 136-145.Duncan DB. 1955. Multiple range and multiple F test. Biometric 11, 1-42.
El-Sheekh MM and Fathy A. 2009. Variation of some nutritional constituents and fatty acid profiles of chlorella vulgaris bei- jerinck grown under auto and heterotrophic conditions. In- tern J Botany 5, 153-159
Girotti AW. 2001. Photosensitized oxidation of membrane lip-