한수지 53(1), 107-111, 2020
107
Copyright © 2020 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815 Korean J Fish Aquat Sci 53(1),107-111,2020
Note
서 론
황돔
(Dentex tumifrons)
은농어목(Perciformes)
도미과(Spari-
dae)
에속하는온대성어류로우리나라중·
남부해,
일본중부이남해역
,
동중국해및타이완북부해역의대륙붕연변부에 분포하는저서정착성어류이다(Chyung, 1977).
황돔의산란장 은분포해역과동일하며,
연중2
회산란하고,
여름에는얕은곳 에서식하다가겨울에는수심이깊은곳으로계절회유를한다(Kim et al., 2004).
황돔은저인망어업에서중요한어획대상 종으로수산자원으로써의가치가매우높은어종으로알려져 있으며,
황돔자원을지속적으로이용하기위해서생태학적연 구는반드시필요하다.
황돔의생태에관한국내연구로는식성(Kim et al., 2012)
이있고,
국외연구로는연령과성장(Oki and Tebeta, 1998),
생식생태(Yoda and Yoneda, 2002; Tominaga et al., 2005; Liao et al., 2009)
등의연구가있다.
하지만이전연구 대부분은동중국해와일본등에집중되어있어국내연안해역에서출현하는황돔에관한생태학적연구는부족한실정이다
.
우리나라연근해어장의어업환경은지속적으로변하고있으 며,
수산자원의생태특성및자원또한변화하고있어우리나라 연근해에서식하는황돔의효율적인자원관리방안과신뢰도 높은자원평가를하기위해서는생태학적인특성을파악하는 것이중요하다.
따라서본연구는우리나라연안해역에서식하 는황돔의자원생태학적정보를제공하기위해성비,
산란기,
포란수,
성숙체장(total length, TL)
등의파라메터를제공하여 황돔의성숙과산란특성을밝히고자한다.
재료 및 방법
황돔시료는남해와동해주변해역에서
2018
년1
월부터12
월까지 매월
1
회 저층트롤망(bottom trawl net)
에 의해채집 된1,062
개체를사용하였다.
채집된시료는전장0.1 cm,
체중0.01 g,
생식소0.01 g
까지측정하였다.
또한,
황돔의월별암·
수간성비차이는
Chi-square test
를 이용하여유의성을검증한국 연안해역에 출현하는 황돔(Dentex tumifrons)의 성숙과 산란
최유정·최정화 1 ·이해원 2 ·정재묵 2 ·김현지 2 ·김도균·진수연·백근욱*
경상대학교, 해양식품생명의학과/해양산업연구소/해양생물교육연구센터, 1국립수산과학원 제주수산연구소, 2국립수산과학원 수산자원연구센터
Maturation and Spawning of Yellowback Seabream Dentex tumifrons in the Coastal Waters of Korea
Yu Jeong Choi, Jung Hwa Choi
1
, Hae Won Lee2
, Jae Mook Jeong2
, Hyeon Ji Kim2
, Do-Gyun Kim, Suyeon Jin and Gun Wook Baeck*Department of Seafood & Aquaculture Science/Institute of Marine Industry/Marine Bio-Education & Research Center, College of Marine Science, Gyeongsang National University, Tongyeong 53064, Korea
1Fisheries Resources and Environment Division, Jeju Fisheries Research Institute, Jeju 63068, Korea
2Fisheries Resources Research Center, National Institute of Fisheries Science, Tongyeong 53064, Korea
In this study, we examined the maturation and spawning of yellowback seabream Dentex tumifrons using samples collected by monthly bottom trawling in Korean coastal waters from January to December 2018. We analyzed month- ly changes in gonadosomatic index (GSI), maturity stage, histology, fecundity, and total length at group maturity.
Spawning occurred in May and October. Fecundity varied with total length as F =1.3754 TL 3.2664 , with a range of 1,220 to 155,625 eggs per female. At group maturity, 50%, 75%, and 97.5% estimates of female total length were 18.4, 20.5, and 25.3 cm, respectively.
Keywords: Dentex tumifrons , Maturation, Spawning
*Corresponding author: Tel: +82. 55. 772. 9156 Fax: +82. 55. 772. 9159 E-mail address: [email protected]
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received 23 December 2019; Revised 24 January 2020; Accepted 12 February 2020 저자 직위: 최유정(대학원생), 최정화(연구관), 이해원(연구사), 정재묵(연구 사), 김현지(연구사), 김도균(대학원생), 진수연(대학원생), 백근욱(교수) https://doi.org/10.5657/KFAS.2020.0107
Korean J Fish Aquat Sci 53(1), 107-111, February 2020
최유정
ㆍ
최정화ㆍ
이해원ㆍ
정재묵ㆍ
김현지ㆍ
김도균ㆍ
진수연ㆍ
백근욱108
하였다
.
생식소의성숙단계는육안으로관찰하였고
,
육안으로판별이 불분명하거나어려운개체의경우,
조직학적관찰을통해구분 하였다.
또한,
황돔암컷의성숙도를미숙(immature),
중숙(ma- turing),
성숙(mature),
완숙(ripe),
방중및방후(spawning and spent)
의5
단계로구분하여분석하였다.
육안관찰방법의경우,
미숙단계
(immature)
의생식소는극히작아육안으로난립을확인할수없었으며
,
중숙단계(maturing)
의생식소는비대해있고크기가다른난이혼재되어있었다
(Kim, 1997).
성숙단계(mature)
는생식소의크기가크고,
난이관찰되기시작하였으나투명난은거의확인할수없었으며
,
완숙단계(ripe)
의생식소는체강에가득하며
,
투명한난이관찰되었다.
방중및방후단계(spawning and spent)
는배를가볍게누르면난이흘러나오거 나생식소의부피가현저히줄어있고생식소내에일부남은알 들이붕괴되어있었다.
조직학적관찰은Oki and Tebeta (1998)
과Tominaga et al. (2005)
와비교하여분석하였다.
산란기추 정을위한생식소숙도지수(gonadosomatic index, GSI)
는다음 의식을이용하여계산하였다(Anderson and Gutreuter, 1983).
GSI=GW/BW×100
여기서
, GW (gonad weight)
는 생식소중량(g), BW (body weight)
는체중(g)
이다.
생식소의내부구조와생식소발달의조직학적변화를관찰하 기위하여생식소를추출하여
Bouin’s solution
에24
시간고정 하였다.
이후,
수세와탈수과정을거쳐paraplast
에포매한후, 6-7 μm
두께로연속절편하여H-E (hematoxylin-eosin)
비교 염색을하였고, marinol
로봉입하였다.
또한,
제작된생식소조 직의표본은광학현미경(LEICA DMIL LED, Leica microsys- tems, Wetzlar, Germany)
을이용하여검경하였다.
포란수는산란기간동안방란의흔적이없는성숙한개체의 난소에대하여다음의습중량법
(Bagenal, 1978)
으로구하였다.
F=
A-BC ×e여기서
, A
는난소의중량, B
는난소껍질의중량, C
는검경한 난소일부의중량, e
는C
에서계수된난의수이다.
상대포란수는성숙한개체의체중에대하여다음식의방법
(Bagenal, 1978)
으로구하였다.
F=aTL
b; F=aTW
b; F=aOW
b여기서
, TL (total length)
은전장, TW (total weight)
는체중, OW (ovary weight)
는생식소중량, a
와b
는상수이다.
성숙체장
(TL)
은산란기로추정되는기간에중숙이상의개체 를당해연도산란가능군으로판단한후,
비율을구하고Logis-
tic
식(King, 2007)
을이용하여추정하였다. P
i= 1
1+
e-b(TL-TLi)여기서
, P
i는i
전장계급에서의성숙비율, TL
i는i
전장계급의 성숙체장(TL), b
는상수이다.
일반적으로사용하는L
50보다엄 격한금지전장설정을위한과학적기준을제시하기위해개체군의
75%, 100%
가성숙하는성숙체장(TL)
을함께추정하였다
.
단,
이번연구에서사용한표본개수와측정오차의영향을 최소화하기위하여100%
에근접하면서도오차가적은것으로 분석된L
97.5를추정하였다.
결 과
성비
2018
년1
월부터12
월까지조사된개체는총1,062
개체였으 며,
그중암컷은508
개체,
수컷은554
로성비는0.9:1 (
암:
수)
로 유의한차이가없었다(P>0.05).
생식소숙도지수(GSI)와 성숙도의 월 변화
2018
년1
월부터12
월까지황돔암컷508
개체를대상으로산 란기를추정하기위하여생식소숙도지수와성숙도의월별변 화를알아본결과(Fig. 1),
생식소숙도지수는2
월에평균0.35
로가장낮은값을나타내었고,
이후,
증가하기시작하여5
월 에평균2.93
으로높은값을보였다(Fig. 1A).
또한, 5
월이후,
감소하여6
월에평균0.57
로낮은값을나타내었으며,
다시증가하여
9
월에평균4.55
로가장높은값을보이다점점감소하는양상을보였다
.
월별성숙도의경우, 1
월과2
월에는모두미 숙개체로나타났으며,
이후,
점점감소하는양상을보이다6
월 에증가하였고,
다시감소한후, 12
월에높은비율로출현하였 다(Fig. 1B).
중숙단계는3
월부터관찰되기시작하여11
월에높 은비율을나타낸후, 12
월부터는관찰되지않았다.
완숙단계는1-3
월을제외하고연중관찰되었으며, 6
월에급격히감소한후,
다시증가하여8
월에가장높게출현하였다.
방중및방후단계 는4-12
월까지출현하였으며,
특히, 5
월과10
월에비교적높은 비율을나타내었다.
따라서생식소숙도지수와성숙도의월변 화를통한황돔의산란기는4-12
월이며,
주산란기는5
월과10
월로1
년에2
회의산란기를가지는것으로추정된다.
난소의 조직학적 관찰
황돔암컷의생식소를조직학적으로살펴본결과
(Fig. 2),
난 소가활성화되기시작하여초기성장기(early growing stage)
에 난소소엽내에서난원세포와초기난모세포가출현하였다(Fig.
2A).
난소소엽내의난모세포들이성장하여대부분후기성장기(late growing stage)
의난소를가지며,
난소내에는세포질의피 질층에공포를갖는난황포기및난황과립을갖는난모세포들황돔의 성숙과 산란
109
이나타나기시작하여난소소엽을채웠다
(Fig. 2B).
이후,
성숙기
(mature)
의난이출현하였는데,
이시기에는난소의발달이급격히진행되어대부분의난소내의난모세포들은성장하여 다량의난황이축적되었고
,
난모세포핵막부위의핵내에서는 인이나타나는난황형성개시기의성숙난모세포들이출현하였 다(Fig. 2C).
완숙기(ripe)
의난소소엽내에는완숙난으로가득 채워져있었으며,
일부호산성과립과공포형의과립이혼재되 어있었다(Fig. 2D).
또한,
방중및방후(spawning and spent)
의 난소에서는산란후,
배란흔적이보이는난모세포의퇴화및흡 수가되는것을확인할수있었다(Fig. 2E).
포란수
황돔의재생산력을알아보기위해산란기로추정되는기간동 안산란하지않은성숙된개체를대상으로포란수를측정한결 과
(Table 1),
전장에따른절대포란수는전장18.6 cm
개체에서 최소1,220
개,
전장33.7 cm
개체에서최대155,625
개로측정되 었으며,
전장과포란수관계식은F=1.3754TL
3.2664(R
2=0.4646)
로나타났다.
상대포란수는(eggs/cm)
는전장16.0-17.9 cm
크 기군에서평균149
개로가장적게나타났으며,
전장30.0-31.9
cm
크기군에서3,213
개로가장많게나타났다. 성숙체장(TL)
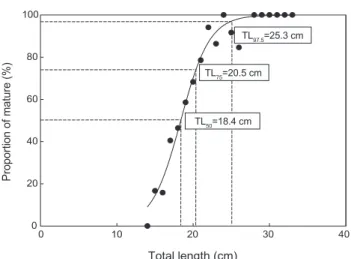
생식에참여가능한개체의크기를알아보기위해산란기로 추정되는기간사이에중숙이상의암컷을대상으로산란하는 것으로간주하여성숙개체의출현비율을나타낸결과
(Fig. 3),
전장
15.0 cm
미만에서는성숙한개체들이출현하지않았으며
,
전장15.8 cm
부터산란에참여가능한성숙한개체들이출현하기시작하였다
.
또한, 28.0 cm
이상의개체들부터는모든 개체들이산란에참여가능한성숙한개체로나타났다.
따라서Logistic
식에적용시킨결과,
황돔암컷의50%, 75%, 97.5%
성 숙체장(TL)
은각각18.4 cm, 20.5 cm, 25.3 cm
로추정되었다.
고 찰
황돔의성비는
0.9:1 (
암:
수)
로암·
수간유사한결과를나타냈 으나,
암컷이수컷에비해비교적낮은성비를보였다.
암·
수간 의성비에서차이를나타내는이유는여러가지로설명될수있 Fig. 1. Monthly changes in gonadsomatic index (GSI; A) and ma-turity stages (B) of female yellowback seabream Dentex tumifrons collected from January to December 2018 in the coastal waters of Korea.
0 20 40 60 80 100
Jan. Feb. Mar. Apr. May Jun. Jul. Aug. Sep. Oct. Nov. Dec.
Fre qu ency
Month
Immature Maturing Mature Ripe Spawning and spent 0
2 4 6 8 10
G SI
(A)
(B)
A B
C D
E
Oc N
Yv
Pof
Total length (cm)
0 10 20 30 40
Pr opo rti on o f m at ur e (%)
0 20 40 60 80 100
TL
75=20.5 cm
TL
50=18.4 cm
TL
97.5=25.3 cm
Fig. 2. Photomicrographs of ovarian development phases of fe- male yellowback seabream Dentex tumifrons in the coastal waters of Korea. A, Early growing stage; B, Late growing stage; C, Ma- ture stage; D, Ripe stage; E, Spawning and spent stage. Abbrevia- tion; N, nucleus; Oc, oocyte; Yv, yolk vesicle, Pof, Post-ovulatory follicles. Scale bars=100 μm.
0 20 40 60 80 100
Jan. Feb. Mar. Apr. May Jun. Jul. Aug. Sep. Oct. Nov. Dec.
Fre qu ency
Month
Immature Maturing Mature Ripe Spawning and spent 0
2 4 6 8 10
G SI
(A)
(B)
A B
C D
E
Oc N
Yv
Pof
Total length (cm)
0 10 20 30 40
Pr opo rti on o f m at ur e (%)
0 20 40 60 80 100
TL
75=20.5 cm
TL
50=18.4 cm
TL
97.5=25.3 cm
최유정
ㆍ
최정화ㆍ
이해원ㆍ
정재묵ㆍ
김현지ㆍ
김도균ㆍ
진수연ㆍ
백근욱110
는데
,
알에서부화한개체에서암컷과수컷의개체수차이,
성 장에따른암컷과수컷의성장률,
사망률,
수명의차이,
성장에 따른성전환등이있다(Wenner, 1972; Sturm and Salter, 1989).
황돔의경우
,
자성선숙을하는종으로암컷에서수컷으로성전 환이이루어진다는연구결과가보고되어있다(Aoyama, 1955;
Oki and Tebeta, 1998).
본연구에서황돔의성비가다소다르 게나타난이유는성비의차이를나타내는여러가지이유중에 서성장에따른성전환산란특성이중요한이유중에하나일것 이라생각된다.
암컷의생식소숙도지수
(GSI)
는5
월에높은값을나타내다가 급격히감소하여6
월에낮은값을나타내었다.
이후,
점차증가 하여9
월에가장높은값을보인후,
다시감소하는결과를나타내었다
.
또한,
암컷의성숙단계의월변화를알아본결과,
성숙 단계를지난개체는4-5
월에높은비율로나타내다6
월에급격 히감소하였으며,
다시8
월에가장높은비율을나타낸이후,
다 시감소하는경향을나타냈다.
이와같은생식소숙도지수(GSI)
와성숙단계의월변화분석결과를보았을때,
황돔의산란기 는4-12
월이며,
주산란기는5
월과10
월이고, 1
년에2
회의산란 기를가지는것으로추정된다.
선행연구결과,
타이완주변해역 에서출현하는황돔은4
월과10
월에산란을하며(Liao et al., 2009),
동중국해에출현하는황돔(Oki and Tebeta, 1998)
은4
월과9
월에산란한다고보고되어져본연구의결과와산란기,
산란횟수(1
년에2
회)
는유사하게나타났지만,
본연구의산란 기가더길게나타나서식해역및해양환경에따라산란기의차 이가있는것으로나타났다(Cha et al., 2007).
산란기는위도가 높을수록산란기가길어지고늦게시작되는데,
이러한조사해 역의환경차이로인하여산란기가다르게나타난것이라생각 된다.
따라서황돔의산란기차이가해양환경변화에따른것인 지혹은,
다른원인으로인한것인지에대해명확히규명하기위 해서는앞으로황돔의생태에관한연구가지속적으로이루어 져야할것으로판단된다.
선행연구결과들과포란수를비교하였을때
, Oki and Tabeta (1998)
는약13,000-35,000
개, Yoda and Yoneda (2002)
는약1,200-29,300
개, Liao et al. (2009)
는약19,772-80,530
개로나 타나선행연구들에비해본연구에서많은포란수의범위를가 지는것으로나타났다.
또한,
성장에따라포란수가증가하는것 을확인할수있었다.
포란수는같은종에있어서도크기,
연령 및영양상태에따라상당히다르며(Kim and Zhang, 1994),
먹 이의분배량과먹이의질,
수온및물리적인환경요인,
서식밀 도,
생존경쟁의정도,
그외성장에영향을주는여러요인들에의해결정된다
(Simpson, 1951).
일반적으로어류는영양상태가좋으면생식소의크기가커지고
,
알의크기를작게하여포란 수가많아진다.
반면,
먹이생물의농도가낮아영양이좋지못할 Fig. 3. A logistic relationship between total length and proportionof mature of female yellowback seabream Dentex tumifrons col- lected from January to December 2018 in the coastal waters of Korea.
0 20 40 60 80 100
Jan. Feb. Mar. Apr. May Jun. Jul. Aug. Sep. Oct. Nov. Dec.
Fre qu ency
Month
Immature Maturing Mature Ripe Spawning and spent 0
2 4 6 8 10
G SI
(A)
(B)
A B
C D
E
Oc N
Yv
Pof
Total length (cm)
0 10 20 30 40
Pr opo rti on o f m at ur e (%)
0 20 40 60 80 100
TL
75=20.5 cm
TL
50=18.4 cm
TL
97.5=25.3 cm
Table 1. Absolute and relative fecundity according to total length of yellowback seabream Dentex tumifons collected from January to De- cember 2018 in the coastal waters of Korea
Total length (cm) Absolute fecundity (eggs) Relative fecundity (eggs/cm)
Range Mean Range Mean n
16.0-17.9 2,670 2,670 149 149 1
18.0-19.9 1,220-56,350 23,575 63-2,923 1,223 12
20.0-21.9 8,520-81,600 36,194 408-3,904 1,732 26
22.0-23.9 13,280-96,740 57,437 576-4,196 2,491 23
24.0-25.9 11,430-97,600 63,655 462-3,942 2,571 13
26.0-27.9 30,960-99,050 70,553 1,134-3,628 2,584 11
28.0-29.9 26,240-132,870 81,748 905-4,584 2,820 26
30.0-31.9 58,690-141,980 98,682 1,911-4,622 3,213 19
32.0-33.9 86,280-155,625 105,270 2,608-4,705 3,182 5
황돔의 성숙과 산란
111
때에는부화율이낮은알을만들거나
,
알의크기가다양해지는 것으로알려져있다(Kim and Zhang, 1994).
따라서황돔또한 전장이커질수록포란수도증가하는것이확인되어다른일반 경골어류와유사한경향을나타내는것으로판단된다.
황돔에관한군성숙도변화에대한기존연구를살펴보면
,
대 마난류가흐르는동중국해,
와카사만,
타이완주변해역에출현 한황돔의성숙체장(50%)
은각각14.7 cm (FL), 13.7 cm (FL), 15.5 cm (FL)
이었다(Oki and Tabeta, 1998; Tominaga et al., 2005; Liao et al., 2009).
본연구의성숙체장과비교하기위해Huh et al. (2017)
을적용하여체장을전장으로환산하였을때,
각각약17.6 cm (TL), 16.4 cm (TL), 18.5 cm (TL)
로환산되었 으며,
본연구와약0.1-2.0 cm
차이를나타냈다.
본연구의결과
,
황돔의50%
성숙체장은18.4 cm (TL)
로선 행연구결과와비슷하게나타났지만,
다소차이를보이는것은 조사시기와관련한해양환경의변화,
개체수및계군의차이등 과도관계가있는것으로생각된다.
황돔자원을보다체계적으 로관리하기위해서는지속적인모니터링을통하여해양환경 변화에따른황돔자원의산란생태변화를지속적으로연구해 야할것이다.
또한,
이번연구는우리나라황돔자원의평가와 관리방안을마련하기위한과학적근거자료를제공하여황돔 의자원관리에있어중요한자료가될것으로생각된다.
사 사
본연구는국립수산과학원수산과학연구사업
(
근해어업자원조사
, R2020021)
의지원에의해수행되었습니다.
References
Anderson RO and Gutreuter SJ. 1983. Length, weight and as- sociated structural indices in Nielson LA and Johson DL (des.) Fisheries Techniques. American Fisheries Society, Bethesda, MD, U.S.A., 468.
Aoyama T. 1955. On the hermaphroditism in the yellow sea bream, Taius tumifrons. Nippon Suisan Gakkaishi 22, 679- 684. https://doi.org/10.11369/jji1950.4.119.
Bagenal TB. 1978. Methods for assessment of fish production in freshwaters. Blackwell scientific publications, Oxford, U.K., 300.
Cha HK, Lee SI, Yoon SC, Kim YS, Chun YY, Chang DS and Yang JH. 2007. Maturation and spawning of the pacific cod,
Gadus macrocephalus TILESIUS in East Sea of Korea. J
Korean Soc Fish Ocean Technol 43, 320-328. https://doi.org/10.3796/KSFT.2007.43.4.320.
Chyung MK. 1977. The fishes of Korea. Ilji-sa, Seoul, Korea, Huh SH, Jeong JM and Park JM. 2017. Length-weight and 360.
length-length relationships for six commercial fishes from southern Korean waters. J Appl Ichthyol 33, 649-651.
https://doi.org/10.1111/jai.13345.
Kim S and Zhang CI. 1994. Fish ecology. Seoul Press, Seoul, Korea, 274.
Kim YS. 1997. Fishery biology. Shingeung Press, Busan, Ko- rea, 57.
King MG. 2007. Fisheries biology, assessment and manage- ment: 2nded. Blackewll Publication, Oxford, U.K., 382.
Kim HW, Beack GW, Park JM and Huh SH. 2012. Feeding hab- its of yellowback seabream, Dentex tumifrons, in the coastal waters of Busan, Korea. Korean J Ichthyol 24, 35-40.
Liao CH, Pong M, Chyn SS and Su WC. 2009. Reproductive biology of the yellow seabream (Dentex tumifrons) in the Northern Waters off Taiwan. J Fish Soc Taiwan 36, 233-250.
http://dx.doi.org/10.29822%2fJFST.200912.0001.
Kim YS, Han KH, Kang CB and Kim JB. 2004. Commercial fishes of the coastal and offshore waters in Korea, 2nd de.
Natl Fish Ress Dev Ins, Busan, Korea, 1-333.
Oki D and Tebeta O. 1998. Age, growth and reproductive characteristics of the yellow seabream Dentex tumifrons in the East China Sea. Fish Sci 64, 191-197. https://doi.
org/10.2331/fishsci.64.191.
Simpson AC. 1951. The fecundity of plaice. Fish Invest London Ser 17, 1-27.
Sturm MG and Salter P. 1989. Age, growth, and reproduction of the king mackerel Scomberomorus cavalla (Cuvier) in Trinidad waters. Fish Bull 88, 361-370.
Tominaga O, Inoue M, Kamata M and Seikai T. 2005. Reproduc- tive cycle of yellow sea bream Dentex tumifrons in Wakasa Bay, the Sea of Japan off central Honshu. Fish Sci 71, 1069- 1077. http://dx.doi.org/10.1111/j.1444-2906.2005.01065.x.
Wenner A. 1972. Sex ratio as a function of size in marine cruste- cea. Amer Natur 106, 310-350.
Yoda M and Yoneda M. 2002. Assessment of spawning fre- quency and batch fecundity in the yellow sea bream, Den-
tex tumifrons. Fish Sci 68, 443-444. https://doi.org/10.2331/
fishsci.68.sup1_443.