Inhibition of Foodborne Pathogens and Spoilage Bacteria and Their Structural Changes by Ethanol Extract of Schizandra chinensis Baillon
Se Ryoung Kim1 and Mee Ra Kim2†
1Dept. of Food Science and Nutrition, Kyungpook National University, Daegu 702-701, Korea
2Dept. of Food Science and Nutrition, Center for Beautiful Aging, Kyungpook National University, Daegu 702-701, Korea
오미자 에탄올 추출물에 의한 식품위해성 세균의 증식 억제 및 세포구조 변화
김 세 령1․김 미 라2†
1경북대학교 식품영양학과, 2경북대학교 식품영양학과․장수생활과학연구소
Abstract
This study analyzed the antibacterial activity of the ethanol extract of Schizandra chinensis Baillon against food pathogenic microorganisms to determine its capabilities as a natural antimicrobial agent. A paper disc diffusion test, minimum inhibitory concentration (MIC) determination, and time-kill assay showed that the ethanol extract strongly inhibits the growth of Listeria monocytogenes, Bacillus cereus, Escherichia coli O157:H7, and Pseudomonas aeruginosa. Release of cytoplasmic β -galactosidase was detected in E. coli, E. coli O157:H7, S. aureus, and P. aeruginosa treated with the ethanol extract. An increase of outer membrane permeability caused by the ethanol extract was also observed. An outward flow of cell constituents was detected in the Gram negative strains treated with the ethanol extract. These results imply that the inner and outer membranes of cells were partially destroyed and cell constituents were released by the treatment of the S. chinensis Baillon ethanol extract. The results of this study indicate that ethanol extract of S. chinensis Baillon evidences a fairly good anti- bacterial effect.
Key words : Schizandra chinensis Baillon, antibacterial activity, MIC, β-galactosidase activity, electron microscopy.
†Corresponding author : Mee Ra Kim, Tel: +82-53-950-6233, Fax: +82-53-950-6229, E-mail: [email protected]
Introduction
Schizandra chinensis Baillon is a deciduous woody vine native to the forests of Northern China and Far East Russia (Kim et al 2009). The berries are called Omija in Korea. Its name comes from the fact that the berries possess all five tastes: salty, sweet, sour, hot, and bitter. It has been consumed as a food utilizing its pinkish-red color for traditional sweets, tea, juice, and wine. In traditional oriental medicine, it has been used as a tonic and has curative effects for bronchial problems, asthma, cardiac disorders, and wound closure.
The biological functions and benefits of S. chinensis Baillon have been studied, such as its antioxidant and anticancer properties. Jeon et al (2008) reported that the DPPH radical- scavenging activity of ethanol extract from S. chinensis Bail- lon was 57.2% at 500 μg /assay, which was higher than the activity of water and methanol extracts of Omija. Jang et al
(1996) reported that antioxidative index values of methanol and ethanol extracts during thermal oxidation (180±2℃) were greater than that of BHT. Lee and Lee (1991), Kim et al (2004), and Kim et al (2009) also reported that S. chinensis Baillon showed antioxidant effects. Jung et al (2000) demon- strated that the butanol fraction of the methanol extract of S.
chinensis seed had effective nitrite scavenging ability.
Han et al (1999) reported that tea prepared with S. chinen- sis Baillon has heavy metal removability that increases when the concentration of S. chinensis Baillon and the extraction temperature increase. Bachri et al (2009) found that a S. chi- nensis Baillon beverage was very effective at combating fati- gue, improving endurance, and increasing overall physical ac- tivity.
Several researchers have reported on the antibacterial acti- vity of the water extract of S. chinensis Baillon (Ji et al 2001, Lee et al 2001, Lee & Lim 1997), but there are scarce reports on the antimicrobial activity of the ethanol extract of S. chi- nensis Baillon. Although the yield of the water extract is rela-
tively higher than that of the ethanol extract, the quality of the water extract is somewhat impaired (Oh et al 1990, Ryu &
Cho 2004). Therefore, the antibacterial activity of ethanol extract of S. chinensis Baillon against microorganisms related to foodborne illness was evaluated by various methods, inclu- ding paper disc diffusion tests, minimum inhibitory concen- tration (MIC), and time-kill assays in this study. Additionally, the effect of the ethanol extract on the structure of the micro- organisms was determined by electron microscopy, cell consti- tuent release tests, outer membrane permeabilization assay, and β-galactosidase activity.
Materials and Methods
1. Preparation of Ethanol Extract
Dried fruits of S. chinensis Baillon were purchased from a traditional market in Daegu, Korea. Ethanol extract of S. chi- nensis Baillon was prepared by refluxing 100 g of the dried fruits in 2 L of 70% ethanol at room temperature for two days. The extract was filtered through filter paper (No. 2, Ad- ventec, Japan) and evaporated under reduced pressure. It was then freeze-dried and kept in a freezer (18℃).
2. Microbial Strains and Media
The activity of the ethanol extract was tested on 7 different microorganisms. The Gram negative bacteria included Esche- richia coli (KCTC 1682), Escherichia coli O157:H7 (ATCC 43894), Pseudomonas aeruginosa (KCTC 1750), and Salmo- nella Typhimurium (KCTC 2515), and the Gram positive strains were Listeria monocytogenes (KCTC 3569), Bacillus cereus (KCTC 1012), and Staphylococcus aureus (KCTC 1916). All strains were obtained from the Korea Biological Resource Center. The bacteria were sub-cultured on nutrient agar (Acu- media, USA).
3. Paper Disc Diffusion Assay
Antibacterial activity was measured by the paper disc di- ffusion method. The diluted inoculum suspension (100 μL) containing approximately 1×108 colony forming units (CFU) /mL was applied onto the nutrient agar plate. The ethanol extract was dissolved in distilled water to a final concen- tration range of 1.5~4.0 mg/disc and sterilized by filtering through a 0.22 μm syringe filter. The plates were placed at room temperature for 2 hr. The filter paper discs (Advantec,
Tokyo, Japan) were individually impregnated with 50 μL of ethanol extract at 1.5~4.0 mg/disc and placed on the plates.
The plates were incubated at 37℃ for 24 hr and the dia- meters of the inhibition zones around the discs were mea- sured. Filter paper discs containing dimethyl sulfoxide with- out the extract were also incubated as a negative control.
Additionally, discs with ρ-hydroxybenzoic acid (Sigma, USA) (1.5~4.0 mg/disc) were incubated as a positive control. All determinations were performed in duplicate.
4. Minimum Inhibitory Concentration (MIC)
A modified micro-well dilution method was used to deter- mine the MIC (Demirci et al 2008). Stock solutions of the ethanol extract were prepared with sterilized distilled water at a final concentration range of 0.5~5.0 mg/mL. Each strain was cultured at 37℃ and the culture was diluted to 1×108 CFU/mL. The concentration of the ethanol extract solution ranged from 0.5 to 5.0 mg/mL and 100 μL of the culture was added to each well of a 96-well plate. A positive control con- tained the bacteria-inoculated medium without ethanol extract and a negative control contained medium without ethanol ex- tract. The 96-well plate was incubated at 37℃ for 24 hr. The tests were carried out in triplicate. The MIC was defined as the lowest concentration of ethanol extract at which the mi- croorganisms did not demonstrate growth.
5. Time-Kill Assay
One colony of microorganisms grown on nutrient agar was chosen and suspended in nutrient broth at 37℃ for 16 hr. The pre-incubated culture was adjusted to 1×108 CFU/mL and 100 μL of suspension was added to 50 mL of media. The ethanol extract was added to the media to a final concentration of 1,000 ppm or 3,000 ppm. The flask was incubated with sha- king at 37℃ and growth of the bacteria was analyzed by the change of absorbance at 620 nm at 0, 4, 8, 12, 16, 20, and 24 hr of incubation using a spectrophotometer (DU 800, Beck- man, USA). A flask containing inoculated medium without extract was also incubated under the same conditions, which served as a control.
6. Cell-Constituent Release
The cells were pre-incubated with nutrient broth at 37℃ for 12 hr. They were then harvested and the pellets were washed with phosphate buffer (pH 7.0) three times. The cell pellets
were resuspended in 100 mL of phosphate buffer and then the final concentration was adjusted to the same as the MIC of each strain by adding the ethanol extract. This suspension was incubated at 37℃ for 24 hr. The suspension was filtered though a syringe filter (pore size, 0.22 μm) to remove the bacteria and the absorbance was measured at 260 nm at 0 and 24 hr of incubation time. Sterile distilled water instead of the ethanol extract was used for the control. The experiments were performed in triplicate.
7. Permeability of the Outer Membrane
Permeabilization activity of the outer membrane by the etha- nol extract of S. chinensis Baillon was determined according to Liu et al.’s method (2004). The cells were pre-incubated with nutrient broth at 37℃ for 12 hr. The cell suspensions were centrifuged and the supernatants discarded. The pellets were washed with sterilized 0.5% NaCl solution three times.
The final cell suspensions were adjusted to 1×108 CFL/mL.
The ethanol extract (1 mL) was mixed with 20 μL of 1 mM N-phenyl-1-naphthylamine (NPN) (Sigma, USA). After the addition of 1 mL of cell suspension to the solution, fluore- scence intensity was immediately measured with a F-2500 fluorescence spectrophotometer (Hitachi, Japan) at an excita- tion wavelength of 350 nm and an emission wavelength of 420 nm every 20 sec for two minutes.
8. β-Galactosidase Activity
β-Galactosidase activity is easily measured with o-nitro- phenyl-β-D-galactoside (ONPG) a colorless substrate, but in the presence of β-galactosidase it turns yellow. Therefore, β-ga- lactosidase activity can be measured by its absorption at 420 nm (Miller 1972). Pre-incubated microbial suspensions were added to M9 media to adjust to an absorbance of 0.25 ± 0.003 at 620 nm. Five-hundred milliliters of suspension was mixed with 500 μL of Z-buffer and 500 μL of ethanol extract solu- tion to a final concentration of 1,000 ppm. After vortexing for 10 sec, 500 μL of toluene was added. To volatilize the to- luene, the test tubes without caps were put into a water bath at 37℃ for 40 min and then left at 28℃ for 5 min. A 200 microliter amount of ONPG (4 mg/mL) was added and held for 20 hr to develop the yellow color due to the enzyme reac- tion. The reaction was stopped by adding 500 μL of 1 M Na2CO3 solution and the absorbance of the supernatant of the solution was measured at 420 nm using a spectrophotometer
(Beckman, USA).
9. Scanning Electron Microscope (SEM)
The changes in cell morphology and functional mutation by treatment of the ethanol extract were observed using a SEM.
The microorganisms were pre-incubated in 10 mL of nutrient broth until the absorbance was 0.9. The ethanol extract in distilled water (1 mL) was added to 4 mL of nutrient broth to a final extract concentration of 1,000 ppm. The pre-incubated culture (500 μL) was added to the suspension. A control was also prepared under the same conditions except for addition of the ethanol extract. After incubation at 37℃ for 12 hr, the cells were harvested by centrifugation. The cell pellets were washed with 0.1 M phosphate buffer (pH 7.2) three times and then the cells were centrifuged again. After the cells were treated with 2% glutaraldehyde, they were kept on ice for 90 min and the supernatant was removed gently by pipetting. De- hydration of the bacterial specimens was carried out sequen- tially with 50%, 70%, 80%, 90%, and 99.99% ethanol. Then the specimens were treated with isoamylacetate and kept at room temperature overnight (Kim M 2007, Park et al 2005).
Finally, the supernatant was removed by pipetting, and after platinum coating under 20 mA for 90 s, the morphology of the bacterial specimens was observed using a FE-SEM (S- 4300, Hitachi, Japan).
10. Transmission Electron Microscope (TEM) Cell morphology was also observed using a TEM. The pro- cedures for incubation and cell harvest were the same as for SEM. The cells were washed with 0.1 M phosphate buffer and treated with 2% glutaraldehyde and then post-fixed in 1%
OsO4 at room temperature for 2 hr. The supernatant was re- moved and the fixed cells were mixed with 2% agar solution at a ratio of 1.5:1, which would become a homogenated agar block. The agar block was dehydrated with ethyl alcohol and propylene oxide, embedded in Epon 812, and cured at 60℃ for 48 hr. Ultramicrothin sections were obtained by an ul- tramicrotome (MT-X, RMC, USA) and samples were placed on a 300-mesh grid. They were double- stained with uranyl acetate and lead citrate and observed with a TEM (model 7600, Hitach, Japan) at a voltage of 80.0 kV.
11. Fast-Atom Bombardment/Mass Spectroscopy (FAB/MS) Positive and negative mode FAB/MS were measured up to
the molecular weight of the ethanol extract of S. chinensis Baillon. The extract was dissolved into m-nitrobenzylalcohol.
The solution was coated onto the FAB probe tip prior to bombardment. A JMS700 mass spectrometer (Jeol Co., Japan) was used at an accelerating voltage of 4 kV and fitted with a 10 kV post-acceleration detector. Fast-atom bombardment was performed using an Ion Tech saddle-field FAB gun ope- rated at 5 kV and with Xe gas.
Results
1. Paper Disc Diffusion Test
The results of the agar disc diffusion test of the ethanol ex- tract from S. chinensis Baillon are shown in Table 1. The dia- meter size of the inhibition zone for L. monocytogenes was the largest (23.03 mm), which implies that the growth of L.
monocytogenes was the most sensitive to the ethanol extract.
On the plates of Bacillus cereus, Escherichia coli O157:H7, and Pseudomonas aeruginosa, inhibition zones were relatively large indicating that the ethanol extract had good antimicro- bial activity against these three strains. As the concentration of the ethanol extract increased, the diameters of the inhibi- tion zones increased for almost strains. Choi et al (2002) reported that ethanol extract from S. chinensis Baillon some- what inhibited the growth of E. coli, but methanol extract showed no inhibition of E. coli growth. Additionally, it was reported that the essential oil of S. chinensis Baillon did not inhibit growth of P. aeruginosa, S. Typhimurium, or E. coli (Lee et al 2003). Compared to these previous studies, the
Table 1. Diameter of clear zones for each microorganism by various concentrations of Schizandra chinensis Baillon etha- nol extract
Microorganism
Concentration (μg/disc)
1,500 2,000 2,500 3,000 3,500 4,000
Bacillus cereus 13.601) 15.05 16.65 17.90 19.25 20.35
Escherichia coli 10.00 10.00 10.20 10.20 10.35 11.20
Escherichia coli O157:H7 16.30 17.10 14.93 15.78 15.80 16.33
Listeria monocytogenes 15.63 16.95 19.03 19.98 20.96 23.03
Pseudomonas aeruginosa 11.00 12.05 13.38 13.95 14.63 15.33
Staphylococcus aureus 10.00 10.50 10.50 11.03 11.53 12.85
Salmonella Typhimurium 10.00 10.10 10.18 10.30 10.60 11.38
1) Sizes are expressed in mm.
ethanol extract from S. chinensis Baillon showed fairly good antimicrobial activity for the strains tested in this study.
2. Minimum Inhibitory Concentration
The results of MIC performed by a micro dilution method are shown in Table 2. L. monocytogenes and E. coli showed the lowest MIC (1.0 mg/mL), whereas S. aureus, P. aerugi- nosa, and S. Typhimurium showed the highest MIC (2.0 mg/
mL). The MIC values of all tested bacteria were in the 1.0~
2.0 mg/mL of range. The ethanol extract affected growth in- hibition of both the Gram positive and Gram negative bacteria.
3. Time-Kill Assay
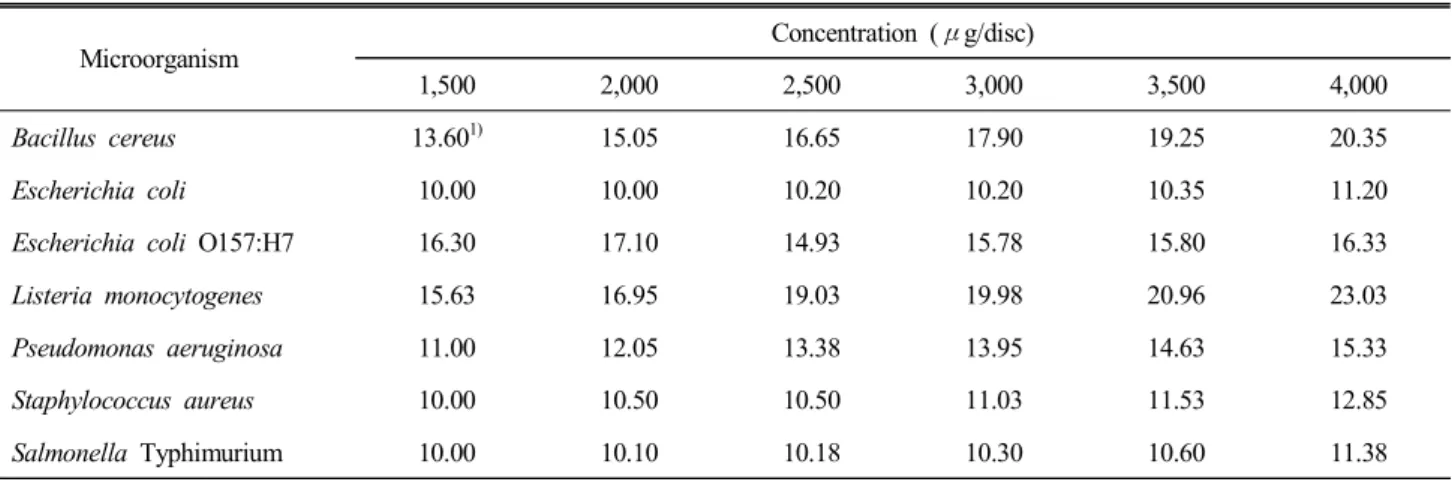
The time-kill curves of some strains are shown in Fig. 1.
Table 2. Minimal inhibition concentration (MIC) of Schi- zandra chinensis Baillon ethanol extract
Microorganisms MIC (mg/mL)
Gram (+)
Bacillus cereus 1.5
Listeria monocytogenes 1.0
Staphylococcus aureus 2.0
Gram ()
Escherichia coli 1.0
Escherichia coli O157:H7 1.5
Pseudomonas aeruginosa 2.0
Salmonella Typhimurium 2.0
Fig. 1. Time-kill curves of microorganisms cultured with Schizandra chinensis Baillon ethanol extract. A: B. cereus, B:
L. monocytogenes, C: E. coli, D: S. aureus. ◆: control , ●: 1,000 ppm, ▲: 3,000 ppm.
A time-kill curve is an instance of bactericidal activity expressed as a progressive change in bacterial growth by a fixed concen- tration of an antimicrobial agent.
The time-kill assays were based on the MIC results. L. mo- nocytogenes and E. coli had a MIC value of 1.0 mg/mL and bacterial growth was completely hindered at 1,000 ppm and 3,000 ppm. The growth of S. aureus, having a MIC value of 2.0 mg/mL, was completely suppressed at 3,000 ppm, which was higher than its MIC value.
4. Cell-Constituent Release
The amount of nucleotides released from cytoplasm can be determined using a UV-visible spectrophotometer at 260 nm, which is widely utilized in the determination of membrane integrity (Chen & Cooper 2002). Fig. 2 shows the variation of absorbance by cell-constituent release after 24 hr of incu-
Fig. 2. Absorbance for nucleotide release of control (white bar) and food pathogenic microorganisms treated with the MIC of Schizandra chinensis Baillon ethanol extract (black bar). A:
B. cereus, B: E. coli, C: E. coli O157:H7, D: L. monocytogenes, E: P. aeruginosa, F: S. aureus, G: S. Typhimurium.
bation. The largest difference in absorbance was between the E. coli control (0.0243) and E. coli treated with S. chinensis Baillon ethanol extract (0.1445). The other Gram negative bac- teria including E. coli O157:H7, P. aeruginosa, and S. Typhi- murium also showed significant differences between the con- trol and treated samples. However, L. monocytogenes, B. cereus, and S. aureus, the Gram positive strains, showed small varia- tions between the control and treated samples.
5. Permeability of Outer Membrane
The uptake of NPN fluorescence is presented in Fig. 3. The fluorescence intensity of the Gram negative strains treated with S. chinensis Baillon increased dependent on time, while that of the control treated with distilled water showed little or just a very low increase. On the other hand, the Gram positive strains, which do not have an outer membrane, showed a ten- dency of maximum fluorescence intensity in one minute and then a sudden drop (data not shown).
6. β-Galactosidase Activity
The activity of β-galactosidase could provide the degree of cell membrane damaged by treatment with the S. chinensis Baillon ethanol extract. The absorbance of the solution treated with distilled water instead of ethanol extract was adjusted to 0%, and the absorbance of the solution treated with toluene was adjusted to 100%. E. coli, E. coli O157:H7, S. aureus, and P. aeruginosa showed that β-galactosidase reacted with ONPG and had relatively high percentages for the reaction (Table 3). E. coli showed the highest percentage of β-galac- tosidase activity (90.4%) and P. aeruginosa and E. coli O157:
H7 also showed over 80%. This result implies that the cell membranes of these strains were severely destructed by treat- ment of the S. chinensis Baillon ethanol extract.
Fig. 3. N-phenyl-1-naphthylamine (NPN) uptake fluorescence after treatment with Schizandra chinensis Baillon ethanol ex- tract for the Gram negative strains. A: E. coli, B: E. coli O157:H7, C: P. aeruginosa, D: S. Typhimurium. ◆: Control, ■:
S. chinensis Baillon ethanol extract.
7. Structure of Cells
The morphologies of the microorganisms treated or untreated with S. chinensis Baillon ethanol extract were observed using a SEM and a TEM. A coarse surface and swelling of cells were found in electron micrographs of E. coli and S. aureus treated with the ethanol extract (Fig. 4). The cells treated with the ethanol extract were larger than the control cells and de- formed by destruction of the cell membrane (Fig. 5). The cells treated with the ethanol extract showed holes or white spots in their membranes, and constituents of E. coli seemed to be released.
8. Fast-Atom Bombardment/Mass Spectroscopy Since positional isomers and stereoisomers of some lignans have the same molecular mass, they may be co-eluted. There- fore, three additional peaks based on their intense [M+H]+ and
Table 3. β-Galactosidase activity of bacterial cell mem- branes treated with Schizandra chinensis Baillon ethanol extract on reaction with o-nitrophenyl-β-D-galactoside
Percentage (%)
Distilled water 0
Toluene 100
E. coli 90.41)
E. coli O157:H7 83.8
S. aureus 72.3
P. aeruginosa 86.3
1) Each value is a relative value to the destructive power of to- luene.
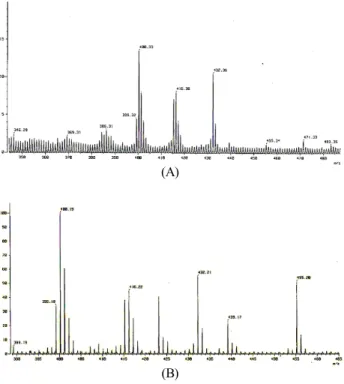
[M+Na]+ ions could be the same tentatively identified com- pounds. The mass chromatograms of the ethanol extract of S.
chinensis Baillon are shown in Fig. 6. Three peaks were ten- tatively identified and the [M]+, [M+H]+, and [M+Na]+ values of the peaks are listed in Table 4. The tentatively identified compounds are Schizandrin, Gomisin A, and Gomisin N. Be- cause the first peak has an intense protonated molecule [M+
H]+ at m/z 433 and an adduction [M+Na]+ at m/z 455, the tentatively identified compound of this peak was presumed to
(A) (B)
(C) (D)
Fig. 4. Scanning electron micrographs of bacteria treated with 1,000 ppm of Schizandra chinensis Baillon ethanol ex- tract (magnification ×50,000). (A): S. aureus control (not-treated), (B): S. aureus treated with 1,000 ppm of Schizandra chinensis Baillon ethanol extract, (C): E. coli control (not-treated), (D): E.
coli treated with 1,000 ppm of Schizandra chinensis Baillon ethanol extract.
(A) (B)
(C) (D)
Fig. 5. Transmission electron micrographs of bacteria treated with 1,000 ppm of Schizandra chinensis ethanol extract. (A):
E. coli control (not-treated, ×7,000), (B): E. coli control (not- treated, ×12,000), (C): E. coli (treated, ×7,000), (D): E. coli (treated,
×12,000).
(A)
(B)
Fig. 6. Simultaneous FAB/MS chromatograms of lignan reference compounds in Schizandra chinensis ethanol extract.
(A): scan range 344.29~487.40 m/z, (B): scan range 388.87~
465.10 m/z.
be Schizandrin. The second peak shows an intense protonated molecule [M+H]+ at m/z 417 and an adduction [M+Na]+ at
Table 4. Peak assignments for analysis of ethanol extract from Schizandra chinensis Baillon
No. [M]+ (m/z)
[M+H]+ (m/z)
[M+Na]+
(m/z) Tentative identification
1 432 433 455 Schizandrin
2 416 417 439 Gomisin A (Schizandrol B)
3 400 401 423 Gomisin N
m/z 439. Therefore, this peak could be Gomisin A. The last peak has an intense protonated molecule [M+H]+ at m/z 401 and an adduction [M+Na]+ at m/z 423, which suggests that this peak could be Gomisin N.
Discussion
The ethanol extract of S. chinensis Baillon exhibited high antimicrobial activity for food pathogenic microorganisms. The ethanol extract showed large growth inhibition zones against Bacillus cereus, Escherichia coli O157:H7, and Listeria mo- nocytogenes in paper disc diffusion tests (Table 1). Moreover, notable growth inhibition by the ethanol extract was shown for L. monocytogenes. According to the results of antimicro- bial experiments, the lowest MIC (1.0 mg/mL) and the largest inhibition zone were detected for L. monocytogenes (Table 2).
Effective growth inhibition against L. monocytogenes by the ethanol extract was also found in a time-kill test (Fig. 1).
These results imply that the ethanol extract of S. chinensis Baillon has an especially strong inhibition effect on the growth of L. monocytogenes. Some previous studies reported that S. chinensis Baillon extract inhibited the growth of S.
aureus and B. subtilis (Chung et al 2001, Kim et al 2003).
In addition, Lee et al (2003) illustrated that S. chinensis contains citronellol and terpineol, which can act as antimicro- bial compounds. On one hand, the essential oil of S. chinensis Baillon did not show antibacterial activity toward Gram ne- gative bacteria (Lee et al 2003), while the ethanol extract of S. chinensis Baillon presented antibacterial activity toward Gram negative bacteria and Gram positive bacteria in this study.
This result could be due to differences in components between the essential oil and ethanol extract of S. chinensis Baillon.
The MIC was lowest for L. monocytogenes and E. coli, followed by B. cereus and E. coli O157:H7. The MIC (1.0 mg/mL) against E. coli and L. monocytogenes was the same but the inhibition zone against E. coli was the smallest. Do-
ming et al (2007) reported on antibiotic susceptibility by broth microdilution and an agar disc diffusion assay. They suggested that there is generally a consistent relationship between broth tests and agar tests, but complete correspondence of results between the two tests does not always occur because of tech- nical limits and different antimicrobial susceptibility for the liquid and solid media. Therefore, they also asserted that broth microdilution assays and agar disc diffusion assays are useful alternative methods.
Cell constituent release, outer membrane permeability, β-ga- lactosidase activity, and electron microscopy were examined to observe how the ethanol extract of S. chinensis Baillon inhibits microbial growth. The release of cytoplasmic β-galac- tosidase was detected in E. coli, E. coli O157:H7, S. aureus, and P. aeruginosa treated with the ethanol extract (Table 3).
This indicates that the permeability of the inner membrane of these bacterial strains increased by the ethanol extract. An increase of outer membrane permeability was also ascertained by measuring fluorescence intensity when the ethanol extract was added to the cell suspensions (Fig. 3). An outward flow of cell constituents was detected in the Gram negative strains treated with the ethanol extract (Fig. 2). Therefore, it was confirmed that the inner and outer membranes of the cells were partially destroyed and the cell constituents containing nucleic acids were released by treatment of the ethanol extract of S. chinensis Baillon. Previously, Helander et al (2001) also found this kind of phenomena during incubation of E. coli, P.
aeruginosa, and S. Typhimurium with chitosan, which has been recognized as a natural antimicrobial agent. Liu et al (2004) reported that the addition of chitosan to E. coli sus- pensions in the presence of NPN led to a time-dependent in- crease in fluorescence. Loh et al (1984) obtained similar re- sults from the incubation of Pseudomonas aeruginosa treated with gentamicin. Therefore, the antibacterial activity of S. chi- nensis Baillon ethanol extract can be explained by the destruc- tion of the outer membrane and the release of cell constituents.
Antibacterial agents can inhibit bacteria growth via several mechanisms. First, some antimicrobial agents can obstruct the formation of cell membranes (Madigan & Martinko 2005).
These types of antimicrobial agents are penicillin, vancomy- cin, bacitracin, cephalosporin, and so on. Second, some can inter- rupt protein production by binding with ribosomes. In addi- tion, other antibacterial agents can inhibit the synthesis of nucleic acids or interfere with cell metabolism by working as
antimetabolites. These antibacterial actions may cause gross membrane damage and instigate cell lysis. Some studies have found these kinds of antibacterial actions. It was reported that essential oils extracted from oregano, rosewood, and thyme caused damage to cell membranes and destroyed bacterial cells (Denyer & Hugo 1991, Hammer et al 1997, Horne et al 2001). Ogunlana et al (1987) also reported that cytoplamic loss occurred in E .coli treated with lemongrass oil. Gustafson et al (1998) reported that autolysis was stimulated and loss of electron dense material was shown in E. coli treated with tea tree oil. In this study, the cells treated with the ethanol extract of S. chinensis Baillon showed alterations in their cell mem- branes, which could have affected membrane permeability.
This speculation is supported by the results of the outer mem- brane permeability assay.
The observation of the cells using SEM and TEM informed that the cells treated with the ethanol extract of S. chinensis Baillon had swelled (Fig. 4) and the constituents of the cells dispersed out (Fig. 5). This microstructural observation demon- strates that the cell membrane structure was severely affected by the extract. The white spots in the electron micrographs of E. coli treated with the ethanol extract could be attributed to the depletion of the cell contents. Cell membrane destruction was also found in several studies on the antimicrobial activity of extracts of natural plants such as Aristolochia contorta Bge (Lee & Cho 2000), Stachys sieboldii MIQ leaf (Ryu & Park 2002), and Gardenia jasminoides (Rye & Cho 2004). Jung et al (2000) reported that the butanol fraction of S. chinensis Baillon destroyed cell walls and the cells swelled.
Cell-constituent release was observed when the S. chinensis Baillon ethanol extract was added to the Gram negative cell culture in this study. Some previous studies also reported that cytoplasmic release by the treatment of antibacterial agents was time-dependent (Bennis et al 2004, Carson et al 2002).
In addition, several studies reported that cytoplasmic release was correlated with the amount of antibacterial agent added (Liu et al 2004, Oussalah et al 2006). NPN fluorescence shows weakly in aqueous environments but becomes very strong in nonpolar or hydrophobic environments (Loh et al 1984, Hancock & Wong 1984). NPN fluorescence increases with an increase of glycerophospholipid (Loh et al 1984, Helander et al 2001). Glycerophospholipid is a component of membranes and located outside of peptidoglycan in Gram negative strains.
NPN is largely exclusive to Gram negative bacteria. When
antimicrobial agents disrupt the outer membrane, glyceropho- spholipids might be released from the cell interior and then interact with NPN. Therefore, it was demonstrated that the S.
chinensis Baillon ethanol extract was effective against Gram negative bacteria to destroy the outer membranes of the cells.
On the other hand, a fluorescence increase did not occur in the S. aureus culture in this study. A similar result was obtained in an experiment of S. aureus treated with chitosan (Loh et al 1984, Chen & Cooper 2002). Additionally, E. coli treated with chitosan showed similar absorbance at different chitosan concentrations, but S. aureus showed that absorbance was correlated with the amount of chitosan added (Liu et al 2004, Oussalah et al 2006). These investigations suggest that Gram positive bacteria may be more sensitive to the concen- tration than treatment time of antimicrobial agents.
Three tentatively identified compounds, Schizandrin, Gomi- sin A, and Gomisin N, were identified in the S. chinensis Baillon ethanol extract by FAB/MS analysis. It was pre- viously reported that S. chinensis Baillon contains these com- pounds (Sohn et al 1989). The contents of Schizandrin, Gomi- sin A, and Gomisin N were 46.9%, 9.2%, and 33.6% of the total lignan in S. chinensis Baillon, respectively. Lignans are known to interrupt nucleotide transport and DNA synthesis and to be specific inhibitors of certain enzymes (Macrae &
Towers 1984). Moreover, lignans have antitumor, antimitotic, antiviral, and antimicrobial activities. The lignans in S. chi- nensis Baillon have been known to have effects on several diseases. Lee et al (1997) gave scientific evidence that lignans in S. chinensis Baillon could affect platelet-activating factor (PAF) and the immune system. It was reported that Schizan- drin, Gomisin A, and Gomisin N inhibited nerve cell extinc- tion. In addition, Schizandrin and Gomisin A were effective on initiation, and Schizandrin worked as an anti-inflammatory and anti-allergic (Hanck et al 1999, Nomura et al 1994). How- ever, because the antibiotic activity of these lignin compounds has not been studied, more research will be needed in this area in the future.
Conclusions
The results of this study demonstrated that the ethanol ex- tract of S. chinensis Baillon has fairly good antibacterial ac- tivity against 7 kinds of food pathogenic microorganisms. The large inhibition zones and the low MIC were shown in L.
monocytogenes, B. cereus, and E. coli O157:H7. The ethanol
extract effectively inhibited the growth of L. monocytogenes in time-kill assay. Furthermore, it was confirmed that the etha- nol extract caused the change of membrane permeability and partially destroyed cell membranes, which could contribute to the antibacterial activity of S. chinensis Baillon. These results suggest that ethanol extract of S. chinensis Baillon is a po- tentially good natural antimicrobial agent.
국문초록
본 연구에서는 오미자 에탄올 추출물의 천연 항균제로서 의 이용 가능성을 살펴보기 위하여 식품 위해성 세균에 대한 항균활성을 측정하였다. Paper disc diffusion test와 최소저해 농도(MIC) 측정에서는 오미자 에탄올 추출물이 Listeria mono- cytogenes, Bacillus cereus, Escherichia coli O157:H7에 대해 큰 생육저해환과 낮은 MIC를 나타내었다. Time-kill assay에 서는 L. monocytogenes의 생육이 오미자 에탄올 추출물에 의 해 가장 저해되는 것으로 나타났다. 또한 오미자 에탄올 추출 물을 처리한 E. coli, E. coli O157:H7, Staphylococcus aureus, Pseudomonas aeruginosa에서는 β-galactosidase와 o-nitrophenyl- β-D-galactoside가 높은 반응을 나타냄으로써 오미자 에탄올 추출물로 인해 세포막 손상이 유발됨을 유추할 수 있었다.
또한 E. coli, E. coli O157:H7, P. aeruginosa, Salmonella Ty-
phimurium 등 그람음성균에서는 오미자 에탄올 추출물의 처
리 농도가 높을수록 세포구성물의 유출과 세포 외막의 투과 성이 증가하는 현상이 나타났다. 시차주사현미경(SEM)과 투 과전자현미경(TEM)을 이용하여 관찰한 세포구조에서도 오미 자 에탄올 추출물 처리 시 세포막의 부분적인 파괴와 세포 팽윤이 일어난 것을 확인할 수 있었다. 따라서 이들 결과는 오미자 에탄올 추출물이 식품위해성 세균에 대해 높은 항균 활성을 가지고 있으며, 천연 항균소재로서의 이용 가능성이 있음을 보여주었다.
Acknowledgments
This Research was supported by Kyungpook National Uni- versity Research Fund, 2009. We thank Dr. Sung Hong Kim at Basic Science Institute Daegu Branch for FAB/MASS ana- lysis.
References
Bachri S, Woo MH, Lee HW, Choi JW, Kim HS (2009) Effects of herbal sports drinks with Omija, Maesil and Molasses on the endurance and energy metabolites of experimental
animals. J Life Science 19: 219-227.
Bennis S, Chami F, Chami N, Bouchikhi T, Remmal A (2004) Surface alteration of Saccharomyces cerevisiae induced by thymol and eugenol. Lett App Microbiol 38: 454-458.
Carson C, Mee BJ, Riley TV (2002) Mechanism of action of Melaleuca alternifolia (tea tree) oil on Staphylococcus au- reus determined by time-kill, lysis, leakage, and salt tole- rance assays and electron microscopy. Antimicrob Agents Ch 46: 1914-1920.
Chen CZ, Cooper SL (2002) Interaction between dendrimer biocides and bacterial membranes. Biomaterials 23: 3359- 3368.
Choi OK, Kim YS, Cho GS, Sung CK (2002) Screening for antimicrobial activity from Korean plants. J Food Sci Nutr 15: 300-306.
Chung KH, Lee SH, Lee YC, Kim JT (2001) Antimicrobial activity of Omija (Schizandra chinensis) extracts. J Korean Soc Food Sci Nutr 30: 127-132.
Demirci F, Guven K, Bemirci B, Dadandi MY, Baser KHC (2008) Antibacterial activity of two Pholmis essential oils against food pathogens. Food Cont 19: 1159-1164.
Denyer SP, Hugo WB (1991) Biocide-induced damage to the bacterial cytoplasmic membrane. In S.P. Denyer, & W.B.
Hugo, (Eds.), Mechanisms of action of chemical biocides:
their study and exploitation. Blackwell Scientific Publica- tions, United Kingdom pp 171-187.
Doming KJ, Mayrhofer S, Zits U, Mair C, Petersson A, Amt- mann E (2007) Antibiotic susceptibility testing of Bifido- bacterium thermophilum and Bifidobacterium pseudolon- gum strains: Broth microdilution vs. agar disc diffusion assay. Int J Food Microbiol 120: 191-195.
Gustafson JE, Liew YC, Chew S, Markham J, Bell HC, Wyllie SG (1998) Effects of tea tree oil on Escherichia coli. Lett App Microbiol 26: 194-198.
Hammer KA, Carson CF, Riley TV (1997) In vitro susceptibi- lity of Malassezia furfur to the essential oil of Melaleuca alternifolia. J Med Vet Mycol 35: 375-377.
Han SH, Shin MK, Kim YW (1999) Effects of Korea Lycium chinense Miller, Schizandra chinensis Baillon, Pueraria thunbergiana Benth, Eucommia ulmoides Oliv tea beverage on the removal of heavy metal. J East Asian Soc Dietary Life 9: 363-369.
Hancke JL, Burgos RA, Ahumada F (1999) Schisandra chi- nensis (Turcz.) Baill. Fitoterapia 70: 451-471.
Hancock RE, Wong PG (1984) Compounds which increase
the permeability of the Pseudomonas aeruginosa outer mem- brane. Antimicrobial Agent and Chemotherapy 26: 48-52.
Helander IM, Nurmiaho-Lassila EL, Ahvenainen R, Fhoades J, Roller S (2001) Chitosan disrupts the barrier properties of the outer membrane of Gram-negative bacteria. Int J Food Microbiol 71: 235-244.
Horne D, Holm M, Oberg C, Chao S, Young DG (2001) An- timicrobial effects of essential oils on Streptococcus pneu- monia. JEOR 13: 387-392.
Jang EH, Pyp YH, Ahn MS (1996) Antioxidant effect of Omija (Schizandra chinensis Baillon) extracts. J Korean Soc Food Sci Nutr 12: 372-376.
Jeon YH, Kil JH, Lim SM, Kim MH, Kim MR (2008) Analysis of antioxidative activity and antimutagenic effect of ethanol extract from Schizandra chinensis Baillon. J East Asian Soc Dietary Life 18: 746-752.
Ji WD, Jeong MS, Chung HC, Choi UK, Jeong WH, Kwoen DJ, Kim SY, Chung YG (2001) Growth inhibition of water extract of Schizandra chinensis Bullion on the bac- teria. J Fd Hyg Safety 16: 89-95.
Jung GT, Ju IO, Choi JS, Hong JS (2000) The antioxidative, antimicrobial and nitrite scavenging effect of Schizandra chinensis Ruprecht (Omija) seed. Korean J Food Sci Tech- nol 32: 928-935.
Kim HK, Na GM, Ye DH, Han HS (2004) Extraction cha- racteristics and antioxidative activity of Schizandra chinen- sis extracts. Korean J Dietary Culture 19: 484-490.
Kim M (2007) Characterization of antimicrobial substance pro- duced by Lactobacillus paraplantarum KNUC25 isolated from Kimchi, MS Thesis Kyungpook National University, Daegu. p 8.
Kim SH, Joo MH, Yoo SH (2009) Structural identification and antioxidant properties of major anthocyanin extracted from Omija (Schizandra chinensis) fruit. J Food Sci 74:
134-140.
Kim YS, Park YS, Lim MH (2003) Antimicrobial activity of Prunus mume and Schizandra chinensis H-20 extracts and their effects on quality of functional Kochujang. Korean J Food Sci Technol 35: 893-897.
Lee IE, Cho SH (2000) The model of antimicrobial mecha- nism of Aristolochia contorta Bge. extract. J Korean Soc Food Sci Nutr 29: 1125-1115.
Lee IS, Jung KY, Oh SR, Kim DS, Kim JH, Lee JJ (1997) Medicinal chemistry; Platelet -activating factor antagonistic activity and 13C NMR assignment of pregomisin and cha-
migrenal from Schisandra chinensis. Arch Pharm Res 20:
633-636.
Lee JS, Lee SW (1991) The studies of composition of fatty acids and antioxidant activities in parts of Omija (Schizan- dra chinensis Baillon). Korean J Dietary Culture 6: 147- 153.
Lee JY, Min YK, Kim HY (2001) Isolation of antimicrobial substance from Schizandra chinensis Baillon and antimi- crobial effect. Korean J Food Sci Technol 33: 389-394.
Lee SH, Lee YC, Yoon SK (2003) Isolation of the antimicro- bial compounds from omija (Schizandra chinensis) extract.
Korean J Food Sci Technol 35: 483-487.
Lee SH, Lim YS (1997) Antimicrobial effects of Schizandra chinensis extract against Listeria monocytogenes. Korean J Microbiol Biotechnol 25: 442-447.
Liu H, Du Y, Wang X, Sun L (2004) Chitosan kills bacteria through cell membrane damage. Int J Food Microbiol 95:
147-155.
Loh B, Grant C, Hancock RE (1984) Use of the fluorescent probe 1-N-phenylnaphthylamine to study the interactions of aminoglycoside antibiotics with the outer membrane of Pseudomonas aeruginosa. Antimicrob Agents Ch 26: 546-551.
Macrae WD, Towers GHN (1984) Biological activities of lig- nans. Phytochemistry 23: 1207-1220.
Madigan M, Martinko JM (2005) Brock biology of micro- organisms 11th ed. Prentice Hall. United States. pp 79-80.
Miller JH (1972) Experiments in molecular genetics. Cold Spring Harbor Laboratory. Cold Spring Harbor Laboratory Press. United States. pp 352-355.
Nomura M, Nakachiyama M, Hida T, Ohtaki Y, Sudo K, Aizzwa T (1994) Gomisin A, a lignan component of Schi- zandora fruits, inhibits development of preneoplastic lesions in rat liver by 3'-methyl-4-dimethylamino-azobenzene. Cancer Lett 76: 11-18.
Ogumlana EO, Höglund S, Onawunmi G, SkÖld O (1987) Effects of lemongrass oil on the morphological characteristics and pepridoglycan synthesis of Escherichia coli cells. Micro- bios 50: 43-59.
Ogunlana EO, Höglund S, Onawunmi G, Sköld O (1987) Effects of lemongrass oil on the morphological characteristics and peptidoglycan synthesis of Escherichia coli cells. Micro- bios 50: 43-59.
Oh SY, Kim SS, Min BY, Chung DH (1990) Composition of free sugars, free amino acid, non-volatile organic acids and tannins in the extracts of L. chinensis M., A. acutiloba K., S. chinensis B. and A. sessiliflorum S. Korean J Food Sci Technol 22: 76-81.
Oussalah M, Caillet S, Lacroix M (2006) Mechanism of ac- tion of Spanish oregano, Chinese cinnamon, and savory essential oils against cell membranes and walls of Escheri- chia coli O157:H7 and Listeria monocytogenes. J Food Prot 69: 1046-1055.
Park KH, Park YM, Seul KJ, Ghim SY (2005) The indirect effects of the near infra-red light-treated materials on mi- crobial growth. Korean J Microbiol Biotechnol 33: 222-225.
Ryu BH, Park BG (2002) Antimicrobial activity of the hexane extract of Stachys sieboldii MIQ leaf. Korean J Life Science 12: 803-811.
Ryu EJ, Cho SH (2004) Antimicrobial characteristics and safety test of Gardenia jasminoides extract. Journal of Agri- culture & Life Science 38: 11-19.
Sohn HJ, Bock JY, Baik SO, Kim YH (1989) Determination of lignan compounds in fruits of Schisandra chinensis Bail- lon by capillary-GC (FID). J Korean Agric Che Soc 32:
350-356.
접 수:
최종수정:
채 택:
2012년 1월 12일 2012년 2월 22일 2012년 2월 23일