저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
A Thesis
For the Degree of Master of Science
Isolation and Characterization of a
Mucin-Degrading Bacterium Attached to
the Rumen Epithelium of Korean Cattle
한우의 반추위벽으로부터 분리한 뮤신 분해
박테리아의 배양 및 특성 규명
August 2018
By
Sang Weon Na
College of Agriculture and Life Sciences
Seoul National University
Isolation and Characterization of a
Mucin-Degrading Bacterium Attached to the Rumen
Epithelium of Korean Cattle
한우의 반추위벽으로부터 분리한 뮤신 분해
박테리아의 배양 및 특성 규명
지도교수
백 명 기
이 논문을 농학석사 학위논문으로 제출함
2018 년 8 월
서울대학교 대학원 농생명공학부
나 상 원
나상원의 농학석사 학위논문을 인준함
2018 년 8 월
위 원 장 (인)
부위원장 (인)
위 원 (인)
i
Abstract
This study was performed to isolate mucin degrading bacteria attached to rumen epithelial cells. Microbial diluent was prepared by extensive homogenization of rumen epithelial cells of the ventral sacs of Korean cattle steers, and it was inoculated with a liquid medium containing a mucin as a sole carbon source in anaerobic condition. Enriched bacterial culture was serially diluted and spread on a mucin agar medium. Single colonies were picked and cultured in the same liquid mucin medium.
A novel bacterium, designated strain E39T, was isolated and its 16S
rRNA gene was sequenced. Phylogenetic analyses based on 16S rRNA gene sequences showed that strain E39T was most closely related to Alloprevotella
rava with an 87.3% sequence similarity. Strain E39T is Gram-negative,
strictly anaerobic, and non-motile coccus. Growth of strain E39T was
observed at 30–45°C (optimum, 39°C), at pH 6.5 and 8.5 (optimum, 7.5), and in the presence of 0.0–1.0% (w/v) NaCl (optimum, 0.0–0.5%). The G+C content of genomic DNA was 46.2 mol%. Enzyme profiles were identified with the Rapid ID 32A identification kit (bioMe´rieux), and this species had was arginine dihydrolase, α-galactosidase, β-galactosidase, glutamic acid decarboxylase, α-fucosidase, alkaline phosphatase, leucyl glycine arylamidase, alanine arylamidase, and glutamyl glutamic acid arylamidase activity. On the basis of the phenotypic, chemotaxonomic, and physiological properties, strain E39T represents a novel genus of the family
Prevotellaceae; the name Psuedoprevotella muciniphila gen. nov., sp. nov. is proposed; the type strain is E39T (= KCTC 15717T= JCM 32621T).
Keyword : Rumen microbiota, epimural bacteria, anaerobic, mucin,
host-gut microbiota interaction
ii
Table of Contents
Abstract ... ⅰ
Table of Contents... ⅱ
List of tables ... ⅳ
List of figures ... ⅴ
List of abbreviations ... ⅶ
Units and marks ... ⅸ
Ⅰ. Introduction ... 1
Ⅱ. Literature review ... 3
1. Rumen microbiology ... 3
1.1. Culture-dependent studies ... 3
1.2. Culture-independent studies ... 5
2. Rumen epithelial bacteria ... 9
2.1. Rumen epithelium and epimural community ... 10
2.2. Diversity of the epimural community ... 10
2.3. Functions of the epimural community ... 13
2.3.1. Tissue recycling ... 13
2.3.2. Oxygen scavenging ... 13
2.3.3. Digestion of urea ... 14
3. Mucin and mucin-degrading bacteria ... 15
3.1. Mucus and mucin ... 15
3.2. Mucin-degrading bacteria ... 16
Ⅲ. Materials and methods ... 18
1. Enrichment and isolation of mucin-degrading bacteria from the
rumen epithelium tissue ... 18
1.1. Media preparation ... 18
1.2. Sample preparation and enrichment of mucin-degrading
bacteria ... 19
1.3. Isolation of mucin-degrading bacteria ... 20
2. Identification of mucin-degrading bacteria based on 16S rRNA
iii
gene sequence ... 20
2.1. DNA extraction and 16S rRNA gene amplicon PCR ... 20
2.2. 16S rRNA gene sequencing and comparative analysis ... 21
3. Characterization of strain E39
T, a novel mucin-degrading
bacterium attached to the rumen epithelium ... 21
3.1. Analysis of 16S rRNA gene squence and construction of
phylogenetic tree ... 22
3.2. Analysis of cell morphology and Gram staining test ... 22
3.3. Growth test ... 23
3.4. Oxygen tolerance test ... 25
3.5. Enzyme profile test and analysis of cellular fatty acid
composition ... 25
3.6. Whole genome sequencing ... 26
Ⅳ. Results ... 27
1. Isolation and identification of mucin-degrading bacteria from the
rumen epithelium tissue ... 27
2. Characterization of strain E39
T, a novel mucin-degrading
bacterium attached to the rumen epithelium ... 28
2.1. Analysis of 16S rRNA gene squence and construction of
phylogenetic tree ... 28
2.2. Phenotypic and growth characteristics of strain E39
T... 33
2.3. Enzyme profile of strain E39
T... 36
2.4. Cellular fatty acid composition of strain E39
T... 38
2.5. Whole genome sequencing ... 40
Ⅴ. Discussion ... 42
Ⅵ. Literature cited ... 47
Ⅶ. Summary in Korean ... 62
iv
List of tables
Table 1. Culture-dependent and –independent methods applied to
rumen microbiology
Table 2. Bacterial composition of the epimural community
Table 3. Mucin medium compositions (per 1,000 ml)
Table 4. Medium compositions for optimum medium test
Table 5. List of isolates identified by 16S rRNA gene sequence
similarity
Table 6. Composition of phenotype characteristics of strain E39T and
closely related taxa
Table 7. Cellular fatty acid compositions (%) of strain E39T and closely
related taxa
v
List of figures
Figure 1. Three different sub-populations of rumen bacteria.
Figure 2. Functions of the epimural community.
Figure 3. Molecular structure of mucins.
Figure 4. Neighbor-joining tree showing the phylogenetic relationship
between strain E39T and closely related strains within the family
Prevotellaceae, based on 16S rRNA gene sequences.
Figure 5. Maximum-likelihood tree showing the phylogenetic
relationship between strain E39T and closely related strains within the
family Prevotellaceae, based on 16S rRNA gene sequences.
Figure 6. Maximum-parsimony tree showing the phylogenetic
relationship between strain E39T and closely related strains within the
family Prevotellaceae, based on 16S rRNA gene sequences.
Figure 7. A transmission electron micrograph (TEM) showing the
general morphology of negatively stained cells of strain E39T grown on
mucin agar medium at 39°C for 3 days.
Figure 8. Growth performance of strain E39T on different media.
vi
Figure 10. Growth performance of strain E39T at different pH units.
Figure 11. Growth performance of strain E39T at different NaCl
concentrations.
Figure 12. Growth performance of strain E39T at different oxygen
vii
List of abbreviations
rRNA : ribosomal ribonucleic acidDNA : Deoxy ribonucleic acid PCR : Polymerase chain reaction FISH : Fluorescent in situ hybridization NGS : Next-generation sequencing
PCR-DGGE : Polymerase chain reaction-denaturing gradient gel electrophoresis
PCR-TGGE : Polymerase chain reaction-temperature gradient gel electrophoresis
gDNA : Genomic deoxy ribonucleic acid GI : Gastrointestinal
VNTR : A variable number of tandem repeats GalNac : N-acetylgalactosamine
GlcNAc : N-acetylglucosamine
EDTA : Ethylenediaminetetraacetic acid VFA : Volatile fatty acid
ADS : Anaerobic dilution solution NJ : Neighbor-joining method ML : Maximum-likelihood method MP : Maximum-parsimony method OD : Optical density
BHI : Brain heart infusion TSB : Trypticase soy broth
PYG : Peptone-yeast extract-glucose TEM : Transmission electron micrograph CDS : Coding sequences
viii tmRNA : transfer-messenger RNA GAG : Glycosaminoglycan
ix
Units and marks
°C : Degree Celcius% : Percent
mol% : Mole percent g : Gram mg : Milligram ml : Milliliter N : Normality h : Hour nm : Nanometer
1
Ⅰ
. Introduction
Rumen microbiota have important roles in the digestion process in the host ruminant. Rumen microbes can be divided into three groups according to their positions in the rumen: (1) floating in the rumen fluid, (2) attached to feed particles, and (3) attached to the rumen epithelium (Cheng et al., 1979; Puniya et al., 2015). Among them, bacteria attached to the rumen epithelium (epimural bacteria) comprise only a small proportion of the total rumen microbial biomass (less than 1%) but play important roles in oxygen scavenging, digestion of urea, and epithelial tissue recycling (Cheng et al., 1979; Sadet et al., 2007).
Mucin, a glycoprotein derived from epithelial cells in the host gastrointestinal tract, forms a mucosal barrier to prevent pathogens from penetrating the intestinal tract. Epithelial cells form two types of mucus barriers: secreted mucin and cell membrane-associated mucin (McGuckin et al., 2011). Although they have the potential to be pathogens, not all mucolytic bacteria are pathogens. Commensal mucolytic bacteria provide carbon and nitrogen, derived from mucin degradation, to other bacteria. Studies on Akkermansia muciniphila, a commensal mucin-degrading bacterium, revealed that this bacterium improved host metabolic disorders, such as obesity and insulin resistance, by controlling gut barrier permeability and inflammation (Derrien et al., 2004; Everard et al., 2013).
The host-gut microbiota interaction is currently being highlighted as a major issue, but there are few studies on the interaction between the rumen epithelial microbiota and the rumen epithelium. This is likely due to the relatively small contribution of rumen epithelial microbiota to feed digestion and the difficulty in their cultivation.
2
In this study, I used mucin as a novel carbon source to enrich mucin-degrading bacteria from the rumen epithelium of Korean cattle. Then, I isolated and characterized a novel mucin-degrading bacterium, called strain E39T.
3
Ⅱ
. Literature review
1. Rumen microbiology
Rumen is a unique foregut that ruminant animals have and it is an evolved symbiotic environment for various microorganisms including archaea, bacteria, protozoa, and fungi (Puniya et al., 2015). Studies on microorganisms in the rumen have been conducted to discover their roles in ruminant nutrition for a long time and experimental methods have also changed with the development of technology. Nowadays, experimental methods of rumen microbiology were classified into culture-dependent methods and culture-independent methods depending on the need of cultivation (Table 1).
1.1. Culture-dependent studies
In the mid-1900s, researchers tried to cultivate rumen microorganisms in artificial media in order to reveal their characteristics affecting fermentation and dysfunctions in the rumen (Hungate, 1944, 1950; Bladen et al., 1961). However, it was hard to cultivate rumen microbes because they required fastidious conditions such as strictly anaerobic condition, various energy sources, and unknown growth factors for growth. Many attempts have been made to cultivate strictly anaerobic rumen microbes in artificial media by imitating the rumen environment (Hungate, 1950; Smith and Hungate, 1958; Kistner, 1960). Finally, Robert E. Hungate, father of rumen microbiology, established the anaerobic culture technique called as Hungate technique (Hungate, 1969; Bryant, 1972; Krause et al., 2013). Since then many researchers like Marvin P. Bryant, have continuously
4
studied ecological and physiological characteristics of anaerobic rumen microbes using the Hungate technique with some modifications (Puniya et al., 2015). The Hungate technique is still used to cultivate anaerobic microbes from various environments as well as the rumen of ruminants. In addition to basic anaerobic culture technique of Hungate, various anaerobic techniques have been developed to completely simulate the rumen environment. For example, anaerobic jar, chamber, and pouch systems were developed to create strictly anaerobic condition.
In the late 1900s, the development of molecular techniques enabled a methodical approach to ecology, physiology, and metabolism of anaerobic rumen microbes. The application of polymerase chain reaction (PCR), cloning, gel electrophoresis and sequencing to rumen microbiology have helped to characterize the genes and enzymes involved in rumen microorganisms. Barros and Thomson (1987) cloned a cellulose gene from Ruminococcus flavefaciens in Escherichia coli and UTT et al. (1991) identified a novel bifunctional protein which has both of β-xylosidase and α-arabinofuranosidase activities from the ruminal bacterium Butyrivibrio fibrisolvens using sequencing and cloning techniques. Many studies on fiber-degrading enzymes contributed to understanding the mechanism of fiber degradation and the use of cellulosic biomass for biofuel production (Béguin and Aubert, 1994; Ohmiya et al., 2005; Christopherson and Suen, 2013).
Since Carl woese and George E. Fox (1977) used 16S rRNA gene sequences for phylogenetic analysis, classification of bacteria has been rapid and accurate. 16S rRNA analysis using the Sanger sequencing technique is an easy method that widely used to identify a cultured bacterium and also has applied to rumen microbial ecology (Whitford et al., 1998; Deng et al.,
5
2008). Although only 20–40% of total rumen microbes have been cultivated, identification and characterization of a novel rumen bacterium using cultivation-dependent methods are necessary to deeply understand physiology and metabolism of rumen microbial ecosystem (Zhang and Dong, 2009; Li et al., 2012; McCann et al., 2014; Firkins and Yu, 2015).
1.2. Culture-independent studies
Studies on rumen microbial ecology using sequence-based analysis has become possible without cultivation with the development of other molecular techniques such as fluorescent in situ hybridization (FISH), real-time PCR, genetic fingerprinting, cloning and next-generation sequencing (NGS) techniques. Comparative analysis with the sequences of rumen microbial genes enabled to design DNA probes for detecting and enumerating targeted rumen microorganisms through various methods (Amann et al., 2000).
FISH is a representative method that can detect and enumerate target cells using small DNA probes labeled with fluorescent. The fluorescent labeled DNA probes penetrate cell membrane and hydridized to a target gene (Vaughan et al., 1999; Deng et al., 2008). Yanagita et al. (2000) detected a ruminal methanogen, Methanomicrobium mobile using 16S rRNA targeted probes by FISH. Shinkai et al. (2007) used FISH and real-time PCR to localize ruminal cellulolytic bacteria including Fibrobacter succinogenes and Ruminococcus flavefaciens on plant fibrous materials.
Real-time PCR is a widely used experimental method that monitors amplification of a target DNA sequence in real time using fluorescent dyes and this technique has used to quantify the cell number of microorganisms
6
as well as the gene expression (Ludwig and Schleifer, 2000; Fraga et al., 2014). Tajima et al. (2001) designed specific primers based on the 16S rRNA gene sequences of several rumen bacteria and investigated the effect of shift to high grain diet on microbial population by real-time PCR.
Genetic fingerprint techniques including polymerase chain reaction-denaturing gradient gel electrophoresis (PCR-DGGE) and polymerase chain reaction-temperature gradient gel electrophoresis (PCR-TGGE) were used to study rumen microbial diversity without cultivation (Sadet et al., 2007; Mackie et al., 2000). Amplified ribosomal DNA fragments were separated by their denaturation characteristics when they passed through gradient of denaturant or temperature (Muyzer, 1999; Deng et al., 2008). Kocherginskaya et al. (2001) compared rumen bacterial diversity between corn diet and hay diet by PCR-DGGE method.
Before the application of NGS to researches on microbiology, researchers conducted transformation of 16S rRNA clones into culturable microorganisms to identify the diversity of the microbial community including culturable and unculturable microbes at one experiment (Deng et a., 2008). Cho et al. (2006) obtained total 113 16S rRNA clones from three different fractions of rumen and revealed that 62.2% of the total clones haven’t been cultured or identified yet.
NGS, also called as high-throughput sequencing, means a new sequencing technology that can sequence a large number of DNA fragments at a time and this technology has been used for different two purposes: 1) whole genome shotgun sequencing (metagenomics) and 2) 16S rRNA amplicon sequencing (metataxonimcs) (McCann et al., 2014; Marchesi and Ravel, 2015). Various microbial communities from various environments such as soil, sea water, human feces, rumen fluid, and fossils
7
could be used as samples for metagenomics or metataxonomics analysis without cultivation. Genomic DNA (gDNA) is extracted and then, libraries are constructed in different ways depending on the purpose. For whole genome sequencing, gDNA is fragmented followed by adapter ligation in the library construction process whereas 16S rRNA variable regions of metagenome are amplified for 16S rRNA amplicon sequencing (Mardis, 2008; Sangschagrin and Yergeau, 2014). Whole genome shotgun sequencing provides the gene and genome profiles of the microbial community and 16S rRNA amplicon sequencing provides microbial abundance and composition (Breitwieser et al., 2017). In rumen microbiology, Hess et al. (2011) identified 27,755 putative carbohydrate-active genes and obtained 15 genomes of uncultured microbes from plant-attached rumen microbes using metagenomic genome sequencing and Myer et al. (2016) investigated the diversity of the rumen bacterial community using 16S rRNA amplicon sequencing technology with two different sequencing platforms. In addition to many researchers have used NGS technologies for overall understanding of the rumen microbial community and its influence on host metabolism including nutrition and immune system (Morgavi et al., 2012; McCann et al., 2014; Firkins and Yu 2015)
8
Table 1. Culture-dependent and -independent methods applied to rumen microbiology
Methods Techniques Analysis Uses References
Culture-dependent methods
Hungate technique Anaerobic culture technique Cultivation of an anaerobic microorganism Hungate, 1969; Zhang and Dong, 2009 Cloning Bacterial gene cloning Isolation and characterization of
a microbial enzyme Barros and Thomson, 1987; UTT et al., 1991 Sanger sequencing 16S/18S rRNA sequencing Identification of a microorganism Zhang and Dong, 2009
Culture-independent methods
FISH DNA hydridization Detection and enumeration of a microorganism Yanagita et al. 2000; Shinkai et al., 2007 Real-time PCR 16S rRNA gene target PCR Detection and enumeration of a microorganism Tajima et al., 2001;
Shinkai et al., 2007
PCR-DGGE/TGGE Genetic finger printing Identification of a microbial diversity Kocherginskaya et al., 2001; Sadet et al., 2007 Clone library 16S rRNA gene library Identification of a microbial diversity Cho et al., 2006
Bacterial gene library Isolation of microbial enzymes Ferrer et al., 2005 NGS 16S rRNA gene sequencing Identification of a microbial diversity Li et al., 2012; Myer et
al., 2016 Genome sequencing Determining the genome sequence of
9
2. Rumen epithelial adherent bacteria
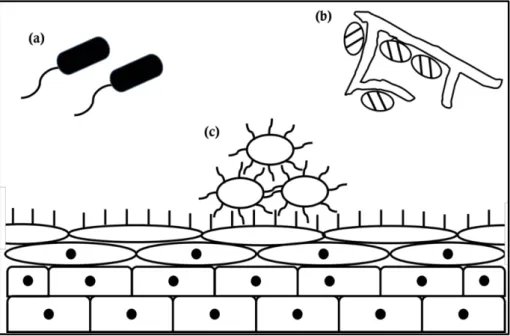
The rumen ecosystem has a complex environment in which the epithelium tissues of host ruminants are enclosed and rumen fluid and feed particles fill the inside of rumen (Puniya et al., 2015). Rumen bacteria are grouped in places that fit their characteristics (Figure 1). Most rumen bacteria float in rumen fluid or attach to feed particles. However, only 1–2% of total rumen bacteria attach to rumen epithelial tissues and they are called as rumen epithelial adherent bacteria or epimural bacteria (Cheng et al., 1979; Sadet et al., 2007; Li et al., 2012).
Figure 1. Three different sub-populations of rumen bacteria. Diagram
shows three different bacterial sub-populations in the rumen: (a) planktonic bacteria, (b) feed attached bacteria, and (c) rumen epithelial adherent bacteria (Cheng et al., 1979).
10
2.1. Rumen epithelium and epimural community
Early studies on the epimural community have mainly conducted from 1970s to 1980s using electron microscopy and culture-dependent methods (Li et al., 2012). Lavker et al. (1969) investigated the morphology of bovine rumen epithelial tissues using a transmission electron microscope and Tamate et al. (1971) observed bovine rumen epithelial tissues and adherent bacteria using a scanning electron microscope. The outer layer of rumen epithelium (called as stratum corneum) consist of partially keratinized and mucus-containing horny cells and epimural bacteria attach to the glycocalyx of horny cells using their fibrous carbohydrate coats (Lavker et al., 1969; McCowan et al., 1978). Epimural bacteria were observed not only on epithelial surfaces but also in cavities within the epithelial cells (Dinsdale et al., 1980). McCowan et al. (1978) observed that epimural bacteria had morphological heterogeneity and consisted of the large number of Gram-positive bacteria whereas Gram-negative bacteria are predominant in rumen fluid.
2.2. Diversity of the epimural community
Researchers have isolated adherent bacteria from rumen epithelium with culture-dependent methods and the isolates were identified as several species involved in the genera Acidaminococcus, Bacteroides, Bifidobacterium, Butyrivibrio, Clostridium, Corynebacterium, Eubacterium, Fusobacterium, Lachnospira, Lactobacillus, Micrococcus, Propionibacterium, Ruminococcus, Staphylococcus, Selenomonas, Streptococcus, Succinivibrio, and Treponema (Cheng et al., 1979; Dehority and Grubb, 1981; Mead and Jones, 1981;
11
Mueller et al., 1984). However, these early studies using culture-dependent methods underestimate the diversity of the epimural community because only culturable bacteria were identified (Li et al., 2012).
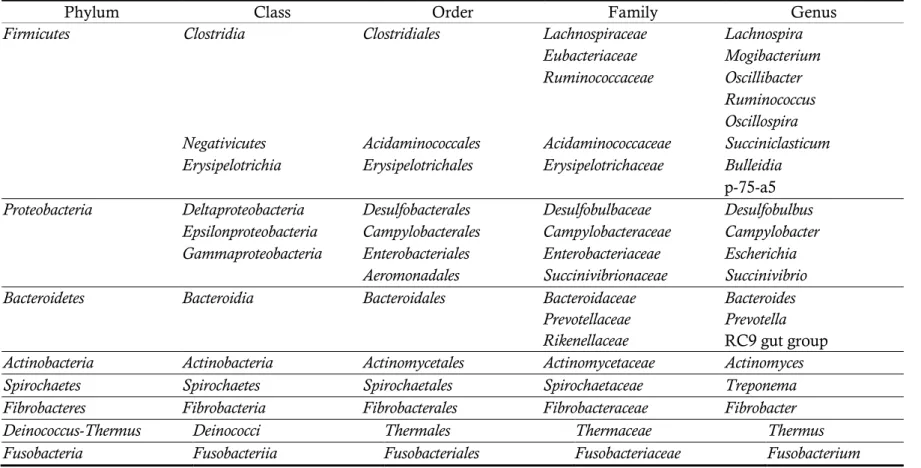
These days, the diversity of the epimural community has been identified with culture-independent methods such as real-time PCR, PCR-DGGE, and NGS technologies (Sadet et al., 2007, 2010; Li et al., 2012; Petri et al., 2013) (Table 2). Li et al. (2012) compared the diversity of the epimural bacterial community to the diversity of the rumen content bacterial community and revealed that the diversity index of epimural bacterial communities were lower than rumen content bacterial communities and bacteria species involved in the phylum Firmicutes, the genera Mogibacterium, and Oscillibacter were predominant in epimural bacterial communities.
12
Table 2. Bacterial composition of the epimural community (Li et al., 2012; Petri et al., 2013)
Phylum Class Order Family Genus
Firmicutes Clostridia Clostridiales Lachnospiraceae Lachnospira
Eubacteriaceae Mogibacterium
Ruminococcaceae Oscillibacter
Ruminococcus Oscillospira
Negativicutes Acidaminococcales Acidaminococcaceae Succiniclasticum
Erysipelotrichia Erysipelotrichales Erysipelotrichaceae Bulleidia
p-75-a5
Proteobacteria Deltaproteobacteria Desulfobacterales Desulfobulbaceae Desulfobulbus
Epsilonproteobacteria Campylobacterales Campylobacteraceae Campylobacter
Gammaproteobacteria Enterobacteriales Enterobacteriaceae Escherichia
Aeromonadales Succinivibrionaceae Succinivibrio
Bacteroidetes Bacteroidia Bacteroidales Bacteroidaceae Bacteroides
Prevotellaceae Prevotella
Rikenellaceae RC9 gut group
Actinobacteria Actinobacteria Actinomycetales Actinomycetaceae Actinomyces
Spirochaetes Spirochaetes Spirochaetales Spirochaetaceae Treponema
Fibrobacteres Fibrobacteria Fibrobacterales Fibrobacteraceae Fibrobacter
Deinococcus-Thermus Deinococci Thermales Thermaceae Thermus
13
2.3. Functions of the epimural community
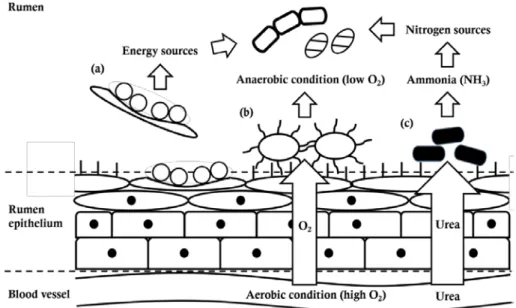
The rumen epithelium is composed of stratified and squamous epithelial cells and exposed to oxygen and urea from blood vessels (Lavker et al., 1969; Cheng and Wallace, 1979; Cheng et al., 1979). These distinctive physiological properties of the rumen epithelial environment are related with three functions of the epimural community: 1) tissue recycling, 2) oxygen scavenging, and 3) digestion of urea (Cheng et al., 1979) (Figure 2).
2.3.1. Tissue recycling
The dead or highly keratinized epithelial cells are continuously sloughed off from the rumen epithelial tissues (Dinsdale et al., 1980). Epimural bacteria colonize on not only the lumenal surface of the epithelial cells but also the undersurface of them and digest dead or wounded epithelial cells (McCowan et al., 1978). Digestion of dead or wounded epithelial cells by epimural bacteria promote absorptive ability of rumen epithelial tissue through the conversion of old cells into new ones and recycle of dead cells as nutrient for the bacteria and finally host ruminant (Cheng et al., 1979).
2.3.2. Oxygen scavenging
The epithelial cells of the host ruminant need oxygen for respiration and blood vessels provide oxygen to rumen epithelial cells. For that reason, most of epimural bacteria have oxygen tolerance or oxygen scavenging activity and their oxygen scavenging abilities contribute to maintain strictly
14
anaerobic conditions of the rumen (Cheng et al., 1979; Mead and Jones, 1981; Mueller et al., 1984).
2.3.3. Digestion of urea
The urea in the rumen comes from feed intake, endogenous urea in saliva, and influx from blood vessels across the rumen epithelium and the digestion of the urea by ureolytic bacteria provides nitrogen sources for synthesis of bacterial proteins (Cheng and Wallace, 1979). Wallace et al. (1979) revealed that facultative epimural bacteria had major role in degradation of urea and ureolysis in the rumen fluid was mainly derived from epimural bacteria attached to sloughed epithelial cells.
Figure 2. Functions of the epimural community. Epimural community has
three functions: (a) tissue recycling, (b) oxygen scavenger, and (c) digestion of urea (Cheng et al., 1979).
15
3. Mucin and mucin-degrading bacteria
3.1. Mucus and mucin
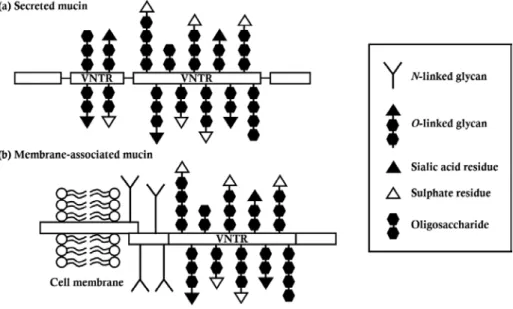
The epithelial cells of animal organs including oral cavity, gastrointestinal (GI) tract, respiratory tract, and reproductive organs are generally covered by viscous and highly hydrated mucus layer. The mucus layer is composed of heavy glycoproteins, called mucins, large amount of water, and other lipids or proteins and it plays many roles as a protective barrier against harmful materials like pathogens, a selective barrier that only pass small nutrient, and a lubricant to help pass food through. Furthermore, components of mucus layers are utilized as energy sources for bacteria (Derrien et al., 2010).
The mucins, constituents of mucus, consist of oligosaccharide branches and protein backbone. The central region of the protein backbone contains a variable number of tandem repeats (VNTR) attached by lots of O-linked glycans and the region called as PTS-region (proline, threonine, and serine rich region). Galactose, acetylgalactosamine (GalNac), N-acetylglucosamine (GlcNAc), and other residues like sialic acid or sulfate constitute O-linked glycans and they have various combinations (Hoorens et al., 2011). The mucins are made in two forms: 1) secreted mucins or 2) membrane-associated mucins (Figure 3). The secreted mucins such as MUC2 were secreted by specific cells of epithelium tissues like goblet cells and they form gel-like filamentous structures through disulfide bonds with each other. The bottom of membrane-associated mucins such as MUC1 is fixed to the membrane of epithelial cells and they act as cell surface receptors and both of protective barriers against pathogens and adhesion
16
spots for adherent bacteria (Derrien et al., 2010).
In case of rumen epithelium, the rumen epithelial cells only express membrane-associated mucins including MUC1, MUC16, and MUC20 whereas other GI tracts like abomasum, large and small intestines are covered by mucus layers consisted of secreted mucins (Hoorens et al., 2011).
Figure 3. Molecular structure of mucins. There are two types of mucins:
(a) secreted mucin and (b) membrane-associated mucin (Derrien et al., 2010; McGuckin et al., 2011).
3.2. Mucin-degrading bacteria
Although mucins protect underlying epithelial cells from pathogens, they also provide energy sources and a place to attach for bacteria. Miller and Hoskins (1981) reported that 1% of the total human fecal microbiota was able to degrade mucins. Researchers have isolated mucin-degrading bacteria from human digestive tract including oral cavity and GI tract and
17
the mucin-degrading bacteria were involved in the genera Akkermansia, Bacteroides, Bifidobacterium, Clostridium, Helicobacter, Prevotella, Ruminococcus, Streptococcus, Streptomyces, and Vibrio (Derrien et al., 2010).
The mucin-degrading bacteria have been considered as potential pathogens because of their ability to degrade mucin. However, mucin-degrading bacteria also positively affect to the host. For example, they stimulate hyperplasia and hypertrophy of goblet cells which secret mucins and prevent abnormal accumulation of mucus in GI tract through degradation of secreted mucins (Kandori et al., 1996). In addition, commensal mucin-degrading may compete with pathogens for attachment to mucins (Derrien et al., 2010). Akkermansia muciniphila, a commensal mucin-degrading bacterium, was isolated from human feces and researchers has observed that a decrease in the abundance of Akkermansia muciniphila in patients with metabolic disorders such as obesity, type 2 diabetes, and cardiovascular diseases (Derrien et al., 2004, 2017). Although Akkermansia species degrade mucins, they also stimulate to secrete mucins from goblet cells in intestinal epithelium (Shin et al., 2014). Also, they strengthen the host-gut immunity by enhancing the production of antimicrobial peptide and the proliferation of regulatory T cells-mediated anti-inflammatory effects (Everard et al., 2013; Shin et al., 2014; Derrien et al., 2017).
About the rumen microbial studies, a few mucin-degrading bacteria were isolated from the rumen and their role in bloat were last studied during the mid-1900s (Fina et al., 1961; Mishra et al., 1967, 1968).
18
Ⅲ
. Materials and Methods
1. Enrichment and isolation of mucin-degrading bacteria from the
rumen epithelium tissue
1.1. Media preparation
The mucin defined medium was prepared with some modification of medium 10 (Caldwell and Bryant, 1966) and consisted of 2 g peptone; 0.5 g yeast extract; 2.5 g hog gastric mucin (Type Ⅲ; Sigma); 50 ml mineral solution 1 (6 g K2HPO4 per liter); 50 ml mineral solution 2 (12 g NaCl, 6 g
KH2PO4, 1.2 g CaCl2.2H2O, 1.2 g MgSO4.7H2O, 12 g (NH4)2SO4 per liter);
10 ml Pfenning’s solution (0.5 g EDTA, 0.1 g ZnSO4.7H2O, 0.03 g
MnCl2.4H2O, 0.03 g H3BO3, 0.2 g CoCl2.6H2O, 0.01 g CuCl2.2H2O, 1.5 g
FeCl2.4H2O, 0.02 g NiCl2.6H2O, 0.03 g Na2MoO4.2H2O, 0.01 g Na2SeO3
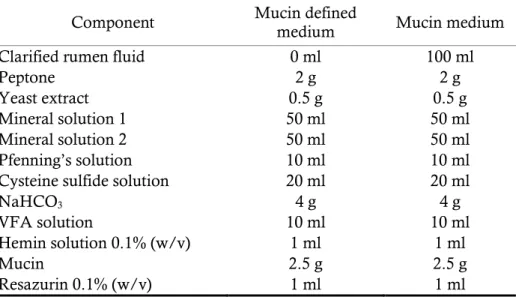
per liter); 10 ml VFA solution (700 ml 0.2 N NaOH, 17 ml acetic acid, 6 ml propionic acid, 4 ml butyric acid, 1 ml iso-butyric acid, 1 ml 2-metylbutyric acid, 1 ml valeric acid, 1 ml iso-valeric acid, pH 7.5 per liter); 1 ml hemin solution (0.5 g hemin, 10 ml 1 N NaOH per liter); 1 ml resazurin (0.1%, w/v); 20 ml cysteine sulfide solution (6.25 g NaOH, 25 g cysteine-HCl, 25 g Na2S.9H2O per liter); 4 g NaHCO3 per liter (Table 3).
19
Table 3. Mucin medium compositions (per 1,000 ml)
Component Mucin defined medium Mucin medium
Clarified rumen fluid 0 ml 100 ml
Peptone 2 g 2 g
Yeast extract 0.5 g 0.5 g
Mineral solution 1 50 ml 50 ml
Mineral solution 2 50 ml 50 ml
Pfenning’s solution 10 ml 10 ml
Cysteine sulfide solution 20 ml 20 ml
NaHCO3 4 g 4 g
VFA solution 10 ml 10 ml
Hemin solution 0.1% (w/v) 1 ml 1 ml
Mucin 2.5 g 2.5 g
Resazurin 0.1% (w/v) 1 ml 1 ml
1.2. Sample preparation and enrichment of mucin-degrading bacteria
Strain E39T was isolated from the rumen epithelium of Korean cattle.
The rumen epithelium tissue samples were excised from ventral sac of the rumen immediately after Korean cattle were slaughtered at the abattoir in Bucheon (37°31'48.4"N 126°45'46.3"E) of South Korea and transported in a sterile container with rumen fluid. On arriving in laboratory, samples were moved into anaerobic chamber and washed several times with anaerobic dilution solution (ADS; 3 g K2HPO4, 6 g NaCl, 3 g KH2PO4, 0.6
g CaCl2.2H2O, 0.6 g MgSO4.7H2O, 6 g (NH4)2SO4, 0.5 g cysteine-HCl, 0.5
g Na2S.9H2O, 0.625 g NaOH,1 mg resazurin per liter) to remove rumen
contents and non-adherent bacteria (Mead and Jones, 1981; Muller et al., 1984). Five grams of epithelial surfaces stripped from muscle layer were homogenized in 30 ml of ADS and serially diluted (10-fold). Each 0.3 ml of dilution was inoculated into 30 ml of mucin medium in butyl rubber stoppered serum bottle and incubated at 39°C in anaerobic atmosphere (95%
20 CO2, 5% H2) for 24 h for enrichment.
1.3. Isolation of mucin-degrading bacteria
Enrichments were serially diluted with ADS and streaked onto mucin agar medium and incubated at 39°C for 4 days under anaerobic condition. Each single colony was picked, inoculated into 5 ml of mucin defined medium in hungate tube and incubated at 39°C for 24–48 h. Streaking, colony picking and incubation were repeated until isolate was pure. The isolates were preserved with 15% (v/v) glycerol stock solution at –80°C.
2. Identification of mucin-degrading bacteria based on 16S rRNA gene sequence
2.1. DNA extraction and 16S rRNA gene amplicon PCR
Genomic DNAs of the isolates were extracted using DNeasy PowerSoil Kit (Qiagen) following manufacturer’s protocol. The 16S rRNA genes of the isolates were amplified by PCR using AccuPower® PCR PreMix (Bioneer) and the universal primers 10F (5′-AGT TTG ATC ATG GCT CAG ATT G-3′) and 1507R (5′-ACC TTG TTA CGA CTT CAC CCC AG-3′) (Lane, 1991) with following step : initial denaturing at 94°C for 3 min, 27 cycles of denaturing (at 94°C for 1 min), annealing (at 55°C for 1 min), and elongation (at 72°C for 1 min), and final extension at 72°C for 7min. PCR products were purified using MEGAquick-SpinTM Total
21 protocol.
2.2. 16S rRNA gene sequencing and comparative analysis
The resulting 16S rRNA amplicons were sequenced at NICEM (Seoul National University, Seoul, South Korea) using the universal primers 10F (5′-AGT TTG ATC ATG GCT CAG ATT G-3′) and 1507R (5′-ACC TTG TTA CGA CTT CAC CCC AG-3′). The sequencing quality was checked and the sequences were assembled using the program Geneious (version 11.0.4) and the 16S rRNA gene sequences were compared with those of all validated type strains using the Nucleotide Similarity Search program in EzBioCloud server (http://www.ezbiocloud.net/identify/) (Yoon et al., 2017).
3. Characterization of strain E39T, a novel mucin-degrading bacterium
attached to the rumen epithelium
I selected isolate E39 for characterization among previously uncultured bacteria which were isolated in this study because isolate E39 grew the best on mucin defined medium and had the lowest 16S rRNA gene sequence similarity (87.3%) with a closely related bacterium. I performed phylogenetic analysis, phenotypic, physiological and biochemical tests to identify characteristics of the isolate E39.
22
3.1. Analysis of 16S rRNA gene sequence and construction of phylogenetic tree
The 16S rRNA gene of strain E39T was sequenced again for obtaining
complete sequence using the universal primers 340F (5′-CCT ACG GGA GGC AGC AG-3′), 518R (5′-ATT ACC GCG GCT GCT GG-3′), and 805F (5′-GAT TAG ATA CCC TGG TAG TC-3′). Sequencing analysis and comparative analysis were conducted with the same methods. Phylogenetic trees were constructed using 16S rRNA gene sequences of strain E39T and closely related taxa and the program MEGA (version 7.0.26)
with neighbor-joining (NJ), likelihood (ML), and maximum-parsimony (MP) methods (Kumar et al., 2016).
3.2. Analysis of cell morphology and Gram staining test
Cell morphology was investigated using phase-contrast microscopy and transmission electron microscopy (Talos L120C; FEI) at 120 kV. For negative staining, cells grown on mucin defined agar medium at 39°C for 3 days were applied onto 200 copper mesh Formvar/carbon-support film grids (Ted Pella Inc.) and stained with phosphotungstic acid. Gram staining was performed using the Sigma Gram staining kit following manufacturer’s protocol.
23
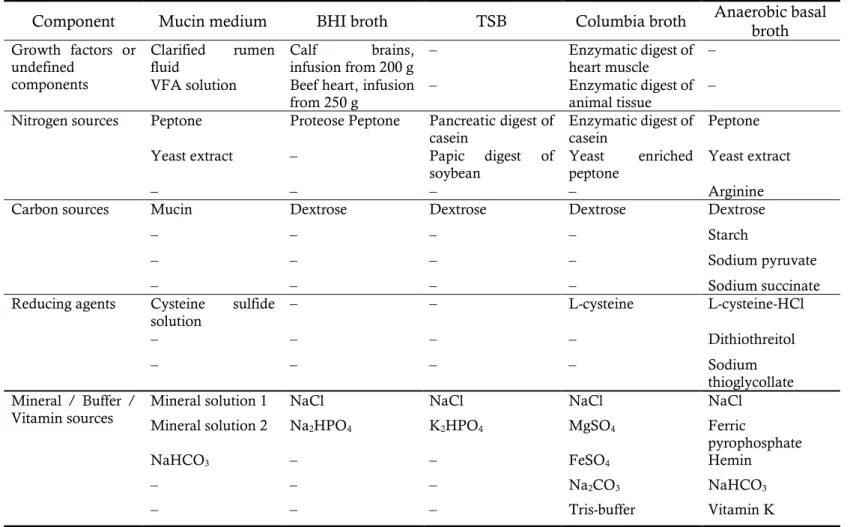
3.3. Growth tests
The growth of strain E39T, as determined from the optical density at a
wavelength of 600 nm, was evaluated by culturing the cells in mucin medium, brain heart infusion (BHI) broth (BD), trypticase soy broth (TSB; BD), Columbia broth (Acumedia), and anaerobe basal broth (Oxoid) (Table 4) at 39°C for 48 h. The optimum temperature, pH, and NaCl concentration for growth were determined by culturing the cells on mucin defined medium for 48 h at different temperatures (5–45°C, at 5°C intervals), pH (5.0–9.0 at 0.5 pH unit intervals), and NaCl concentrations (0.0–2.0% at 0.5% intervals). To determine the optimum pH, different pH buffers were used in the appropriate pH range (Na2HPO4-NaH2PO4 buffer
at pH 5.0–7.5; Tris-HCl buffer at pH 8.0–9.0) and the pH values were adjusted before and after autoclaving (121°C, 15 min) (Gomori, 1955).
24
Table 4. Medium compositions for optimum medium test
Component Mucin medium BHI broth TSB Columbia broth Anaerobic basal broth Growth factors or
undefined components
Clarified rumen
fluid Calf brains, infusion from 200 g – Enzymatic digest of heart muscle – VFA solution Beef heart, infusion
from 250 g – Enzymatic digest of animal tissue – Nitrogen sources Peptone Proteose Peptone Pancreatic digest of
casein Enzymatic digest of casein Peptone
Yeast extract – Papic digest of
soybean Yeast enriched peptone Yeast extract
– – – – Arginine
Carbon sources Mucin Dextrose Dextrose Dextrose Dextrose
– – – – Starch
– – – – Sodium pyruvate
– – – – Sodium succinate
Reducing agents Cysteine sulfide
solution – – L-cysteine L-cysteine-HCl
– – – – Dithiothreitol
– – – – Sodium
thioglycollate Mineral / Buffer /
Vitamin sources Mineral solution 1 Mineral solution 2 NaCl Na NaCl NaCl NaCl
2HPO4 K2HPO4 MgSO4 Ferric
pyrophosphate
NaHCO3 – – FeSO4 Hemin
– – – Na2CO3 NaHCO3
25
3.4. Oxygen tolerance test
Oxygen tolerance was investigated by measuring growth (OD at 600 nm) in the absence of a reducing agent (cysteine sulfide solution) or aerobic condition of headspaces on mucin defined medium.
3.5. Enzyme profile test and analysis of cellular fatty acid composition
Alloprevotella tannerae ATCC 51259T, Alloprevotella rava 81/4-12T, and
Prevotella melaninogenica ATCC 25845T, which is the type species of the
genus Prevotella, were used as reference strains to compare enzyme profiles and cellular fatty acid composition. The freeze-dried cells of Alloprevotella
tannerae ATCC 51259T, Alloprevotella rava 81/4-12T, and Prevotella
melaninogenica ATCC 25845T were recovered on modified chopped meat
medium (ATCC medium 1490), Columbia blood medium (DSMZ medium 693), and modified peptone-yeast extract-glucose (PYG) medium (KCTC medium 282) at 37°C and pH 7.0 for 24–48 h in anaerobic conditions.
The enzyme profiles were determined using an API Rapid ID 32A identification kit (bioMérieux) following the manufacturer’s instructions. Analysis of cellular fatty acids was performed according to a standard MIDI protocol. All of the strains were cultivated in PYG broth, except strain E39T,
which was cultivated in mucin defined medium. Cells were harvested at the late exponential phase and cellular fatty acids were extracted from the cells following four steps (saponification, methylation, extraction, and base wash). Fatty acid methyl esters were analyzed by gas chromatography (Hewlett Packard 6890) and identified using the RTSBA6 database of the Microbial Identification System (Sherlock ver. 6.0B) (Sasser, 1990).
26
3.6 Whole genome sequencing
De novo genome sequencing was performed using an Illumina HiSeq platform at Macrogen (Seoul, Korea; http://www.macrogen.com). A library was prepared using an Illumina TruSeq DNA PCR-Free kit and the library quality was checked before sequencing. After sequencing, reads were assembled using SOAPdenovo2 method (http://soap.genomics.org.cn) (Luo et al., 2012) and functional annotation analysis was performed using the annotation tool in BlastKOALA server (http://www.kegg.jp/blastkoala/) (Kanehisa et al., 2016).
27
Ⅳ
. Results
1. Isolation and identification of mucin-degrading bacteria from the
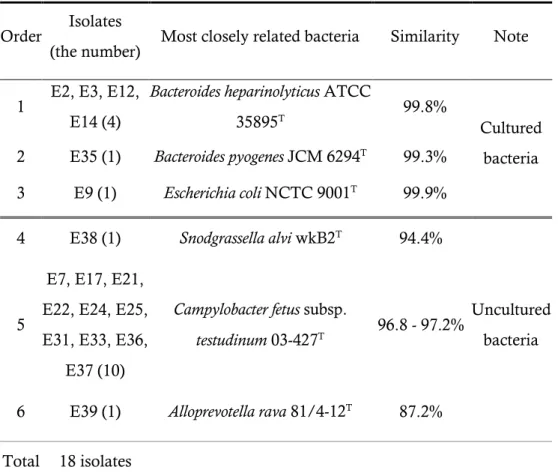
rumen epithelium tissue
Total 39 colonies were picked from mucin agar medium and only 18 isolates were grown on mucin liquid medium. The six of the isolates were most closely related to 3 cultured bacteria (Bacteroides heparinolyticus ATCC 35895T, Bacteroides pyogenes JCM 6294T, and Escherichia coli NCTC 9001T)
with 99.8%, 99.3%, and 99.9% 16S rRNA gene sequence similarities, respectively. And the others were most closely related to 3 uncultured bacteria (Uncultured rumen bacterium clone L102RT, Uncultured rumen bacterium clone L206RT, and Uncultured rumen bacterium clone ZP040) with 99.8, 98.5-99.1, and 89.6% 16S rRNA gene sequence similarities, respectively (Table 5).
28
Table 5. List of isolates identified by 16S rRNA gene sequence similarity
Order Isolates
(the number) Most closely related bacteria Similarity Note 1 E2, E3, E12,
E14 (4)
Bacteroides heparinolyticus ATCC
35895T 99.8%
Cultured bacteria 2 E35 (1) Bacteroides pyogenes JCM 6294T 99.3%
3 E9 (1) Escherichia coli NCTC 9001T 99.9%
4 E38 (1) Snodgrassella alvi wkB2T 94.4%
Uncultured bacteria 5
E7, E17, E21, E22, E24, E25, E31, E33, E36,
E37 (10)
Campylobacter fetus subsp.
testudinum 03-427T 96.8 - 97.2%
6 E39 (1) Alloprevotella rava 81/4-12T 87.2%
29
2. Characterization of strain E39T, a novel mucin-degrading bacterium
attached to the rumen epithelium
2.1. Analysis of 16S rRNA gene sequence and construction of phylogenetic tree
I obtained the almost complete 16S rRNA gene sequence of strain E39T (1,476 nucleotides) and the sequence was submitted to the NCBI
GenBank under the accession number of MG763147. Comparative analysis of the 16S rRNA gene sequences revealed that strain E39T was
closely related to the genera Alloprevotella, Paraprevotella, Prevotella, and Bacteroides. Alloprevotella rava 81/4-12T, Paraprevotella clara YIT 11840T, and
Bacteroides gallinarum JCM 13658T were most closely related to strain E39T
with 87.3%, 87.0%, and 86.4% 16S rRNA gene sequence similarities among cultured bacteria, respectively. And I constructed phylogenetic trees using 16S rRNA gene sequences of strain E39T and species involved in the genera
Alloprevotella, Paraprevotella, Prevotella and Bacteroides with the NJ (Figure 4), ML (Figure 5), and MP (Figure 6) methods.
30
Figure 4. Neighbor-joining tree showing the phylogenetic relationship
between strain E39T and closely related strains within the family
Prevotellaceae, based on 16S rRNA gene sequences. Bootstrap values are shown on nodes in percentages of 1,000 replicates; only values over 70% are shown. Sphingobacterium spiritivorum NCTC 11386T (EF090267) was
used as an outgroup. The scale bar equals 0.02 changes per nucleotide position.
31
Figure 5. Maximum-likelihood tree showing the phylogenetic relationship
between strain E39T and closely related strains within the family
Prevotellaceae, based on 16S rRNA gene sequences. Bootstrap values are shown on nodes in percentages of 1,000 replicates; only values over 70% are shown. Sphingobacterium spiritivorum NCTC 11386T (EF090267) was
used as an outgroup. The scale bar equals 0.1 changes per nucleotide position.
32
Figure 6. Maximum-parsimony tree showing the phylogenetic relationship
between strain E39T and closely related strains within the family
Prevotellaceae, based on 16S rRNA gene sequences. Bootstrap values are shown on nodes in percentages of 1,000 replicates; only values over 70% are shown. Sphingobacterium spiritivorum NCTC 11386T (EF090267) was
used as an outgroup. The scale bar equals 50 changes per nucleotide position.
33
2.2. Phenotypic and growth characteristics of strain E39T
Figure 7. A transmission electron micrograph (TEM) showing the general
morphology of negatively stained cells of strain E39T grown on mucin agar
medium at 39°C for 3 days. Bar, 200nm.
The cells of strain E39T were non-motile, Gram-negative coccus
without flagella (680–820 nm in diameter) and filamentous structures were observed from the cell surface (Figure 7). Strain E39T could grow on only
mucin medium among various media, including BHI broth, TSB, Columbia broth, and anaerobic basal broth (Figure 8). Cells grew at 30– 45°C (optimum, 39°C; Figure 9), pH 6.5–8.5 (optimum, pH 7.5; Figure 10), and 0.0–1.0% NaCl (optimum, 0.0–0.5%; Figure 11), respectively. Cells did not grow in an aerobic atmosphere regardless of whether a reducing agent was present or not. When the headspaces were filled with anaerobic gas, growth was observed in the absence of a reducing agent, but a lower rate than in its presence (Figure 12).
34
Figure 8. Growth performance of strain E39T on different media.
Figure 9. Growth performance of strain E39T at different temperatures.
35
Figure 11. Growth performance of strain E39T at different NaCl
concentrations.
Figure 12. Growth performance of strain E39T at different oxygen
concentrations. NROX: no reducing agent and aerobic headspace; NRAN: no reducing agent and anaerobic headspace; PROX: presence of reducing agent and aerobic headspace; PRAN: presence of reducing agent and anaerobic headspace.
36
2.3. Enzyme profile of strain E39T
In the API Rapid ID 32A panel, strain E39T had positive enzyme
activities for arginine dihydrolase, α-galactosidase, galactosidase, β-galactosidase-6-phosphate, N-acetyl-β-glucosaminidase, glutamic acid decarboxylase, α-fucosidase, alkaline phosphatase, leucyl glycine arylamidase, alanine arylamidase, and glutamyl glutamic acid arylamidase (Table 6).
37
Table 6. Comparison of phenotype characteristics of strain E39T and
closely related taxa. Taxa: 1, strain E39T (this study); 2, Alloprevotella
tannerae ATCC 51259T (Moore et al., 1994); 3, Alloprevotella rava 81/4-12T
(Downes et al., 2013); 4, Prevotella melaninogenica ATCC 25845T (Shah and
Collins, 1990). All strains are positive for the following characteristics: activity* of β-galactosidase, alkaline phosphatase, and alanine arylamidase. All strains are negative for the following characteristics: activity* of urease, β-glucosidase, α-arabinosidase, β-glucuronidase, reduction of nitrates, indole production, proline arylamidase, phenylalanine arylamidase, leucine arylamidase, pyroglutamic acid arylamidase, tyrosine arylamidase, glycine arylamidase, histidine arylamidase, and serine arylamidase. Symbols: +, positive; –, negative; ND, not determined.
Characteristic 1 2 3 4
Morphology Coccus Rod Rod Rod
Optimal temperature (°C) 39 ND 35 37
Optimal pH 7.5 ND 7 ND
Enzyme activity (API Rapid ID 32a) * of
Arginine dihydrolase + − − − α-Galactosidase + − + − β-Galactosidase 6 phosphate + − + + α-Glucosidase − + − + N-Acetyl-β-glucosaminidase + − + + Mannose fermentation − − + + Raffinose fermentation − − + +
Glutamic acid decarboxylase + + − +
α-Fucosidases + + − +
Arginine arylamidase − − − +
Leucyl glycine arylamidase + + − +
Glutamyl glutamic acid arylamidase + + − + DNA G+C content (mol%) 46.2† 46.6† 45.5† 40.9†
*Data were obtained from this study.
38
2.4. Cellular fatty acid composition of strain E39T
The major cellular fatty acids (>5% of the total fatty acids) of strain E39T were C
16:0, C18:0, C18:1ω9c, iso-C15:0 and anteiso-C15:0. Among strain
E39T and the reference taxa of the family Prevotellaceae, iso-C
15:0 and
39
Table 7. Cellular fatty acid compositions (%) of strain E39T and closely
related taxa. Taxa: 1, strain E39T; 2, Alloprevotella tannerae ATCC 51259T;
3, Alloprevotella rava 81/4-12T; 4, Prevotella melaninogenica ATCC 25845T. All
data were obtained from this study. Data are expressed as percentages of the total fatty acids and fatty acids amounting to less than 0.5% in all strains are not shown. Major components (>5.0%) are highlighted in bold. tr, trace amount (<0.5%); –, not detected.
Fatty acid 1 2 3 4 Saturated: C12:0 0.9 tr tr tr C13:0 0.9 – – – C14:0 4.5 3.0 3.2 1.9 C16:0 24.9 4.9 14.5 8.7 C17:0 1.2 tr 1.7 tr C18:0 18.3 1.3 6.7 0.9 C20:0 0.8 tr 0.6 tr Unsaturated: C13:1 at 12-13 0.6 tr 2.2 2.2 C18:1 ω9c 10.3 0.9 6.7 23.1 Hydroxy: C15:0 3-OH 0.9 tr tr tr C16:0 3-OH 3.7 2.0 0.8 3.5 C17:0 2-OH tr tr 0.7 tr iso-C17:0 3-OH 2.1 9.4 13.3 3.6 Branched: iso-C13:0 1.6 3.51 1.0 0.8 iso-C14:0 – 8.8 – 12.2 iso-C15:0 9.0 26.6 8.4 5.6 iso-C16:0 0.5 2.2 2.4 1.0 iso-C17:0 3.0 4.2 6.7 1.4 iso-C18:0 – tr 1.0 0.1 iso-C19:0 – tr 1.1 0.8 anteiso-C13:0 tr 0.8 tr tr anteiso-C15:0 5.4 25.2 15.5 21.7 anteiso-C17:0 0.9 1.9 2.4 1.4 Summed feature*: 3 0.9 tr 1.0 3.1 5 2.7 tr – – 8 3.5 tr 2.6 3.4
*Summed features represent groups of two fatty acids that cannot be separated by GLC with the MIDI system. Summed feature 3, C16:1ω7c
and/or C16:1ω6c; summed feature 5, C18:0 ante and/or C18:2 ω6,9c;
40
2.5. Whole genome sequencing
Total 2,184,440,594 bases and 14,466,494 read counts were obtained and the reads were assembled into 36 contigs. The genomic DNA G+C content of strain E39T was approximately 46.2 mol%. Annotation analysis
revealed that the genome of strain E39T comprised 2,880,897 bases, 2,428
genes, 2,378 coding sequences (CDSs), 47 tRNA, 1 transfer-messenger RNA (tmRNA), and 2 rRNA. 969 CDSs of total 2,378 CDSs (40.7%) were annotated and classified into many metabolisms involved in 6 KEGG categories (metabolism, genetic information processing, environmental information processing, cellular processes, organismal systems, and human diseases) (Table 8). Genome of strain E39T was submitted to the NCBI Genome under the
41
Table 8. Genome annotation results of strain E39T
No. Categories Sub-catergories Pathways Genes
1 metabolism
Carbohydrate metabolism 15 157
Energy metabolism 7 82
Lipid metabolism 7 21
Nucleotide metabolism 2 80
Amino acid metabolism 14 119 Metabolism of other amino acids 8 22 Glycan biosynthesis and metabolism 8 43 Metabolism of cofactors and vitamins 11 74 Metabolism of terpenoids and
polyketides 6 17
Biosynthesis of other secondary
metabolites 9 23
Xenobiotics biodegradation and
metabolism 1 9
2 information Genetic processing
Transcription 1 3
Translation 3 78
Folding, sorting and degradation 4 24 Replication and repair 5 60 3 Environmental information processing Membrane transport 2 23 Signal transduction 6 15 4 processes Cellular
Transport and catabolism 2 6
Cell growth and death 4 17
Cellular community-prokaryotes 4 22 Cell motility 2 2 5 Organismal systems Immune system 4 5 Endocrine system 7 7 Digestive system 2 2 Nervous system 2 2 Aging 2 4 Environmental adaptation 2 2 6 diseases Human Cancers: Overview 4 4
Cancers: Specific types 1 1
Immune diseases 1 1
Neurodegenerative diseases 1 1
Cardiovascular disease 1 3
Endocrine and metabolic diseases 2 2 Infectious diseases: Bacterial 3 7 Drug resistance: Antimicrobial 3 16 Drug resistance: Antineoplastic 1 5 Total 6 categories 37 sub-categories 157 pathways genes 959
42
Ⅴ
. Discussion
Researches about rumen epithelial adherent bacteria (=epimural bacteria) were conducted mainly from 1970s to 1980s using culture dependent methods and electron microscopy (McCowan et al., 1978; Cheng et al., 1979; Wallace et al., 1979; Mead et al., 1981; Mueller et al., 1984). Since then, however, there were few researches about epimural bacteria until novel microbiology research techniques were developed. Application of cloning, PCR-DGGE and 16S rRNA sequencing techniques has facilitated to identify diversity and composition of a ruminal epithelial community, which were previously underestimated because of limitation of cultural methods (Cho et al., 2006; Sadet et al., 2007; 2010; Li et al., 2012). Despite of recent development of the experimental techniques, many things related with the epimural bacteria including their functions haven’t been completely discovered yet. Li et al. (2012) reported that 67.6% of the 16S rRNA gene sequences from both of rumen tissue samples and rumen contents were uncultured rumen bacteria at the genus level. For deep understanding a ruminal epithelial community, it is needed to isolate and characterize novel epimural bacteria using culture-dependent methods as well as to identify functions of a ruminal epithelial community using culture-independent methods like metagenomics.
In this study, I isolated mucin-degrading bacteria attached to the rumen epithelium using medium contained mucin as a sole carbon source. Total 18 isolates were identified as 3 cultured bacteria and 3 uncultured bacteria (Table 5). Four isolates were most closely related to Bacteroides
43
were most closely related to Bacteroides pyogenes JCM 6294T and Escherichia
coli NCTC 9001T with 99.3% and 99.9% similarity, respectively. Ten
isolates were most closely related to Campylobacter fetus subsp. testudinum 03-427T with 96.8 to 97.2% similarity and two isolates each were most
closely related to Snodgrassella alvi wkB2T and Alloprevotella rava 81/4-12T
with 94.4% and 87.2% similarity, respectively. All of the isolates were belong to the Gram-negative phyla Bacteroidetes (the genera Bacteroides and Alloprevotella) and Proteobacteria (the genera Campylobacter, Escherichia, and Snodgrassella) whereas the Gram-positive phylum Firmicutes is most dominant in a rumen epithelial community (Li et al., 2012). Derrien et al. (2010) reported that mucin-degrading bacteria isolated from human digestive tract belong to the phyla Firmicutes (the genera Streptococcus, Clostridium, and Ruminococcus), Bacteroidetes (the genera Bacteroides and Prevotella), Proteobacteria (the genera Vibrio and Hellicobaceter), Actinobacteria (the genera Streptomyces and Bifidobacterium), and Verrucomicrobia (the genus Akkermansia). It is well known that several Bacteroides species including Bacteroides heparinolyticus and Bacteroides pyogenes have glycosaminoglycans (GAGs; mucopolysaccharides) degrading enzymes such as sulfatase, chondroitinases or herparinase (Nakamura et al., 1988; Ahn et al., 1998; Ulmer et al., 2014). Escherichia coli, well known as intestinal bacteria and pathogens, also have mucin-degrading enzymes (Ulmer et al., 2014). Most Campylobacter species are pathogens and they can bind to mucin and degrade it using mucin-degrading enzymes (Sylvester et al., 1996; Tu et al., 2008). Snodgrassella alvi wkB2T is a bacterium isolated from the gut of honey bees and little is
known about their mucolytic ability (Kwong and Moran, 2013). The genus Alloprevotella was isolated from human oral cavity and they are closely
44
related to the genus Prevotella (Downes et al., 2013). Prevotella sp. RS2, a bacterium belong to the genus Prevotella, has mucin-degrading enzymes including sulfoglycosidase and glycosulfatase (Wright et al., 2000; Rho et al., 2005; Derrien et al., 2010). These results showed that ruminant have their own mucin-degrading bacterial community on the surface of rumen epithelium. Studies with identifying composition and diversity of mucin-degrading epimural community using 16S rRNA sequencing and isolating and characterizing novel mucin-degrading epimural bacteria are required to understand the role of a mucin-degrading epimural community.
Among the 18 isolates, I selected and characterized the isolate E39 because it had the lowest 16S rRNA gene sequence similarity (87.2%) with the most closely related bacterium, Alloprevotella rava 81/4-12T.
The phylogenetic analysis using the NJ algorithm showed that strain E39T was affiliated with the family Prevotellaceae and closed to the genus
Alloprevotella (Downes et al., 2013; Moore et al., 1994) (Figure 4). However, strain E39T formed a distinct phylogenic lineage from the genus
Alloprevotella. The phylogenetic trees based on the MP and ML algorithms also showed that strain E39T formed a distinct phyletic lineage within the
family Prevotellaceae (Figure 5 and 6). The 16S rRNA gene sequence of strain E39T had low similarities with Alloprevotella tannerae ATCC 51259T
(85.1%) and Alloprevotella rave 81/4-12T (87.3%). These molecular and
phylogenetic analysis suggests that strain E39T can represent a novel genus
of the family Prevotellaceae.
Cells of strain E39T were Gram-negative, non-motile coccus whereas
Alloprevotella tannerae and rava are bacilli (Downes et al., 2013). A transmission electron micrograph revealed that filamentous structures of strain E39T (Figure 7). It is assumed that these filamentous structures are
45
bacterial glycocalyx which is polysaccharide containing structure for adhesion to other bacteria or epithelial cells of the rumen (Costerton et al., 1981). And similar structures were observed on the cells of Akkermansia muciniphila, mucin-degrading bacterium isolated from human feces (Derrien et al., 2004).
The results of enzyme profile test and cellular fatty acid analysis showed that small but clear differences in several items between strain E39T
and reference strains. Especially, strain E39T had positive activities on
mucin-degrading enzyme including α-galactosidase, β-galactosidase, N-acetyl-β-glucosaminidase, and α-fucosidases (Derrien et al., 2010).
The whole genome sequencing and annotation analysis showed that strain E39T had many genes which were classified into 165 pathways
involved in 6 categories including metabolism, genetic information processing, environmental information processing, cellular processes, organismal systems, and human diseases (Table 8). Strain E39T had genes
involved in carbohydrate metabolisms (including fructose, mannose, galactose, starch, and sucrose) and VFA metabolisms (including propionate and butyrate). Regarding glycan degradation, strain E39T had
glycan-degrading enzymes including N-acetyl-β-hexosaminidase (EC 3.2.1.52), hyaluronoglucosaminidase (EC 3.2.1.35), N-acetyl-α-glucosaminidase (EC 3.2.1.50), sialidase-1 (EC 3.2.1.18), β-galactosidase (EC 3.2.1.23), and α-fucosidase (EC 3.2.1.51) and these enzymes may contribute to degradation of glycans like hyaluronan, chondroitin sulfate, dermatan sulfate, heparan sulfate, keratan sulfate, and mucin. In addition, strain E39T had also a
putative protease which is used to digest a mucus layer in Helicobacter pylori infection. Common mucin-degrading enzyme activities were detected by enzyme profile test and these results demonstrate that strain E39T has
46
mucolytic ability. The ability to degrade mucin is necessary for pathogens to penetrate the host GI tract (McGuckin et al., 2011). In the rumen of ruminant, it is known that the mucolytic activity of mucin-degrading bacteria cause bloat by degradation of bovine salivary mucin (Fina et al., 1961). However, there are also commensal mucin-degrading bacteria like Akkermansia muciniphila and they prevent pathogens to penetrate host GI tract through competing with pathogens or stimulating the host’s immune system (Derrien et al., 2010, 2017). Strain E39T has all of the potential as a
pathogen or a commensal bacterium. To identify the function of strain E39T,
further studies should be conducted using the combination of culture-dependent and culture-inculture-dependent methods.
In conclusion, the phylogenetic, physiological and chemotaxonomic features clearly support that strain E39T represents a novel genus of the
family Prevotellaceae, for which the name Pseudoprevotella muciniphila gen. nov., sp. nov. is proposed. In the future, it is expected that studies on mucin-degrading bacteria attached to the rumen epithelium may contribute to the improvement in ruminal diseases such as bloat and ruminal acidosis, and a better understanding of host-rumen microbial interactions.
47
Ⅵ
. Literature cited
Ahn MY, Shin KH, Kim DH, Jung EA, Toida T, Linhardt RJ, Kim YS.
Characterization of a Bacteroides species from human intestine that degrades glycosaminoglycans. Can J Microbiol 1998;44:423–429.
Amann R, Ludwig W. Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology. FEMS Microbiol Rev 2000;24:555–565.
Barros MEC, Thomson JA. Cloning and expression in Escherichia coli of a cellulase gene from Ruminococcus flavefaciens. J Bacteriol 1987;169:1760– 1762.
Béguin P, Aubert JP. The biological degradation of cellulose. FEMS Microbiol Rev 1994;13:25–58.
Bladen HA, Bryant MP, Doetsch RN. A study of bacterial species from
the rumen which produce ammonia from protein hydrolysate. Am Soc Microbiol 1961;9:175–180.
Breitwieser FP, Lu J, Salzberg SL. A review of methods and databases for
metagenomics classification and assembly. Brief Bioinform 2017;doi: 10.1093/bib/bbx120.
48
anaerobic bacteria. Am j Clin Nutr 1972;25:1324–1328.
Caldwell DR, Bryant MP. Medium without rumen fluid for nonselective
enumeration and isolation of rumen bacteria. Appl Microbiol 1966;14:794–801.
Cheng KJ, McCowan RP, Costerton JW. Adherent epithelial bacteria in
ruminants and their roles in digestive tract function. Am J Clin Nutr 1979;32:139–148.
Cheng KJ, Wallace RJ. The mechanism of passage of endogenous urea
through the rumen wall and the role of ureolytic epithelial bacteria in the urea flux. Br J Nutr 1979;42:553–557.
Cho SJ, Cho KM, Shin EC, Lim WJ, Hong SY, Choi BR, Kang JM, Lee SM, Kim YH, Kim H, Yun HD. 16S rDNA analysis of bacterial
diversity in three fractions of cow rumen. J Microbiol Biotechnol 2006;16:92–101.
Christopherson MR, Suen G. Nature’s bioreactor: the rumen as a model
for
biofuel production. Biofuels 2013;4:511–521.