284 책임저자:박건영, 609-735, 부산시 금정구 장전동 산30
부산대학교 식품영양학과
Tel: 051-510-2839, Fax: 051-514-3138 E-mail: [email protected]
접수일:2008년 10월 29일, 게재승인일:2008년 11월 7일
Correspondence to:Kun-Young Park
Department of Food Science and Nutrition, Pusan National University, 30, Jangjeon-dong, Geumjeong-gu, Busan 609-735, Korea
Tel: +82-51-510-2839, Fax: +82-51-514-3138 E-mail: [email protected]
HT-29 인체 대장암세포에서 키토산의 항암 효과
1부산대학교 식품영양학과, 2(주)금호화성, 3동주대학 외식조리제과계열, 4경남정보대학 식품과학계열
반영은1ㆍ박순선1ㆍ하종길2ㆍ김소희3ㆍ문숙희4ㆍ박건영1
Anticancer Effect of Chitosan in HT-29 Human Colon Cancer Cells
Young-Eun Bahn1, Soon-Sun Bak1, Jong-Kil Ha2, So-Hee Kim3, Suk-Hee Moon4 and Kun-Young Park1
1Department of Food Science and Nutrition, Pusan National University, Busan 609-735, 2KeumHo Chemical, Seoul 134-711,
3Department of Culinary Art & Baking Technology, Dongju College, Busan 603-715, 4Subdivision of Food Science, Kyungnam College of Information and Technology, Busan 616-701, Korea
Anticancer effects of soluble and non-soluble chitosans were studied in HT-29 human colon cancer cells. Soluble chitosan S-10 (SC) inhibited 88% of the proliferation at 5 mg/ml. Non-soluble chitosan NS-8 (NSC) showed 83% anticancer effects at 1 mg/ml on the growth of the HT-29 cells. SC and NSC inhibited the growth of cells in a time-dependent manner. Chitosans induced apoptosis in the HT-29 cells and it was associated with the increase of Bax, p53, p21 and decrease of Bcl-2. The level of cIAP-1 mRNA expression decreased in NSC. These results suggest that S-10 (DA 90%, vis 5 cps) and NS-8 (DA 95%, vis 22 cps) chitosans especially showed high anticancer effects in the colon cancer cells.
(Cancer Prev Res 13, 284-291, 2008)
Key Words: Soluble/non-soluble chitosan, Anticancer, Apoptosis
서 론
최근 우리나라 사람들은 불규칙한 생활습관과 서구화 된 식습관으로 인하여 질병 및 사망 원인이 변화되고 있 다. 이 중, 만성퇴행성 질환 및 암을 앓고 있는 인구가 급속히 증가되어 가고 있으며, 특히 대장암으로 인한 사 망률의 급속한 증가가 사회 문제로 대두되고 있다. 암 발생의 원인으로 약 75∼80%가 환경적 요인에 의한 것 으로 알려져 있으며, 이 중 약 30∼40%가 식이섭취의 문 제에서 기인된다고 한다.1∼4) 암 치료에 있어서 항암제 요법은 매우 큰 비중을 차지하고 있지만 항암제 자체의 비특이성 및 독성으로 인한 정상세포의 손상은 해결해 야 될 시급한 과제로 남아있으며 현재 이에 대한 연구가
지속적으로 행해지고 있다.5∼7) 항암제의 부정적인 측면 및 암예방에 대한 사고가 높아지면서 기능성 식품 및 암 을 예방할 수 있는 식품에 대한 관심이 높아지고 있다.
키토산은 키틴을 탈아세틸화 하여 만든 동물성 식이 섬유의 일종으로 게, 새우 등의 갑각류와 곤충류의 껍질, 조개 등의 패류 등 천연적으로 자연에 풍부하게 존재하 는 성분이다. 이 키틴질은 천연 고분자 중에서 자연계에 서 셀룰로오즈 다음으로 많이 존재하며 N-acetyl-D- glucosamine이 β-1,4 결합하고 있는 구조를 가지고 있기 때문에 인체 내에서는 소화 효소가 없어서 흡수되기 힘 들다.8) 키토산과 그 유도체는 독성이 낮고, 공해 생성이 적으며, 생분해성을 가지는 것으로 보고되어 있으며, 이 외에도 제산작용, 궤양억제작용, 혈청 콜레스테롤과 중 성지질량을 감소시키며, 장내 유용세균의 발육 촉진에
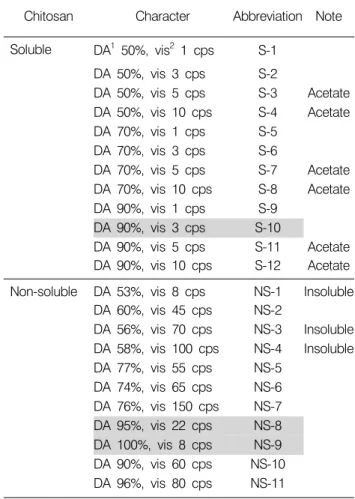
Table 1. Characteristic of chitosan used in this experiment
Chitosan Character Abbreviation Note
Soluble DA1 50%, vis2 1 cps S-1 DA 50%, vis 3 cps S-2
DA 50%, vis 5 cps S-3 Acetate DA 50%, vis 10 cps S-4 Acetate DA 70%, vis 1 cps S-5
DA 70%, vis 3 cps S-6
DA 70%, vis 5 cps S-7 Acetate DA 70%, vis 10 cps S-8 Acetate DA 90%, vis 1 cps S-9
DA 90%, vis 3 cps S-10
DA 90%, vis 5 cps S-11 Acetate DA 90%, vis 10 cps S-12 Acetate Non-soluble DA 53%, vis 8 cps NS-1 Insoluble
DA 60%, vis 45 cps NS-2
DA 56%, vis 70 cps NS-3 Insoluble DA 58%, vis 100 cps NS-4 Insoluble DA 77%, vis 55 cps NS-5
DA 74%, vis 65 cps NS-6 DA 76%, vis 150 cps NS-7 DA 95%, vis 22 cps NS-8 DA 100%, vis 8 cps NS-9 DA 90%, vis 60 cps NS-10 DA 96%, vis 80 cps NS-11
1DA (%): degree of deacethylation.
2vis (cps): viscosity.
영향을 미친다.9,10) 키토산은 대식세포를 활성화시키는 등의 면역 강화 및 조절, 식물병원성진균에 대한 식물의 저항성 유도가 있는 것으로 보고되어 있다. 또한 여러 종류의 암세포나 종양에 대하여 항암 효과가 있는 것으 로 알려져 있다.11∼15) 그러나 수용성 및 불용성 키토산이 apoptosis의 인자들에 어떠한 작용을 하는지에 대한 연구 는 미흡한 실정이다.
또한 다양한 식품에 키토산을 첨가하였을 때, 저장성 이나 기능성이 증가하는 것으로 나타났다.16∼19) 그러나 키토산의 탈아세틸화도와 점도에 따라서 특성 및 기능 성이 달라지므로 본 연구에서는 항암효과가 높은 키토 산을 선별하여 이를 식품산업에 응용하기 위한 연구의 일환으로, 탈아세틸화도 및 점도에 따른 여러 가지 키토 산의 효과를 비교 실험하였다.
키토산의 항암 기능성을 확인하기 위해 탈아세틸화도 및 점도가 다양한 키토산(수용성 12가지, 불용성 11가지) 의 암세포 성장 저해 정도를 확인하고, 그 항암 기전에 대하여 연구하였다. 이를 위하여 여러 가지 키토산의 특 성을 알아보고, HT-29 인체 대장암 세포를 이용하여 암 세포 성장저해효과를 알아보고 apoptotic body를 확인하 였으며, apoptosis 관련 유전자들의 발현 양상을 관찰하였 다.
재료 및 방법 1. 키토산 시료
본 실험에 사용된 chitosan은 탈아세틸화도(Degree of deacethylation, DA (%))와 점도(Viscosity, cps)가 다른 수용 성 chitosan 12종류(S-1∼12)와 불용성 chitosan 11종류 (NS-1∼11)이고, chitosan의 특성은 Table 1에 나타내었다.
키토산 시료는 (주)금호화성(경북, 영덕)에서 영덕산 게 껍질을 건조시키고 칼슘을 제거하여 만든 키틴을 고 온, 강알칼리의 조건하에서 탈아세틸화(DA)하여 제조하 였다. 탈아세틸화도는 건강기능식품공전 중 키토산 분 말 품질기준에 준하여 colloid 적정법으로 측정하였고,20) 점도는 0.5% 키토산을 0.5% 초산에 용해시켜 20oC에서 2시간 용해시켜 brookfield 점도계(USA)를 이용하여 측정 하였다.21) 분자량(Molecular weight)은 Size-Exclusion Chro- matography/Muti-Angle Laser Light Scattering (Wyatt DAWN DSP, POTILAB, Wyatt Techonology, USA)을 이용하여 측 정하였다.22) 이렇게 제조한 키토산 중 수용성 키토산 (S-10)은 PBS를 용매로 불용성 키토산(NS-8)은 1% 초산을 용매로 녹여 10 mg/ml 농도의 stock을 만들어 plastic tube (SPL, 경기도)에 넣어 4oC에서 보관하면서 사용하였다.
2. 키토산의 in vitro 항암효과 및 기작 연구
1) 암세포 배양: HT-29 인체 대장암 세포는 100 units/ml 의 penicillin-streptomycin과 10%의 Fetal bovine serum (FBS) 가 함유된 RPMI 1640 (GIBCO, USA)을 사용하여 37oC, 5% CO2 incubator (Forma, model 311 S/N29035, USA)에서 배양하였다. 배양된 각각의 암세포는 일주일에 2∼3회 refeeding하고 6∼7일 만에 PBS로 세척한 후 0.05% Trypsin- 0.02% EDTA로 부착된 세포를 분리하여 원심분리한 후 집적된 암세포에 배지를 넣고 피펫으로 암세포가 골고 루 분산되도록 잘 혼합하여 75 ml cell culture flask에 10 ml씩 일정 수 분할하여 주입하고 계속 6∼7일마다 계대 배양하면서 실험에 사용하였다.
2) MTT assay: 배양된 암세포를 96 well plate에 well당 2×104 cells/ml가 되도록 180μl씩 분주하고 시료를 일정 농도로 제조하여 20μl 첨가하여 37oC, 5% CO2배양기에 서 72시간 배양하였다. 여기에 5 mg/ml의 농도로 제조한
3-(4,5-dimethyl-thiazol)-2,5-diphenyltetrazolium bromide (MTT) 용액 20μl를 첨가하여 동일한 배양 조건에서 4시간 동 안 더 배양하였다. 이때 생성된 formazan결정을 DMSO에 녹여서 ELISA reader로 540 nm에서 흡광도를 측정하였 다.23,24)
3) Hematocytometer를 이용한 세포 성장률의 측정:
세포배양용 6 well plate를 이용하여 1×105 cells/ml의 농도 로 HT-29 세포를 분주하고 24시간 동안 안정화시킨 후, 키토산김치 시료를 48시간동안 처리하였다. 준비된 세 포를 phosphate buffered saline (PBS) 용액으로 수세한 후, trypan blue로 염색하였다. 이를 hematocytometer에 옮긴 후 위상차 현미경(×200) 하에서 살아있는 세포의 수를 측정하였다.25)
4) DAPI 염색을 통한 핵의 관찰: 시료의 처리에 의한 암
세포의 apoptosis 유발 여부를 확인하기 위한 핵의 형태변 화 관찰을 위하여 시료가 처리된 세포들을 PBS로 수세 하고 3.7% paraformaldhyde로 상온에서 10분간 고정시킨 후 형광 염색물질인 4,6-diamidino-2-phenylindole (DAPI) 용액을 이용하여 10분간 염색하였다. 이들 세포를 다시 PBS로 2회 수세한 후 광학현미경(Olympus BX50, Japan) 을 이용하여 핵의 형태 변화를 정상군과 비교하였다.26) 5) Reverse transcription-polymerase chain re- action (RT-PCR) 분석: Trizol을 이용하여 total RNA를 분 리하였다. Oligo dT primer를 사용하면 분리한 RNA 중 mRNA가 모두 cDNA로 만들어지기 때문에 분리된 RNA 를 정량한 후, oligo dT primer와 AMV reverse transcriptase 를 이용하여 2μg의 RNA에서 mRNA에 상보적인 ss cDNA로 역전사 시켰다. 이 cDNA를 template로 사용하여 Bax, Bcl-2, p53, p21, cIAP-1, cIAP-2 유전자를 polymerase chain reaction (PCR) 방법으로 특정 유전자 부위를 증폭하 였다. 이때 housekeeping 유전자인 glyceraldehyde-3-phos- phate dehydrogenase (GAPDH) 유전자를 포함하여 internal control로 사용하였다. 각 PCR 산물들을 1% agarose gel을 이용하여 전기영동하고 ethidium bromide (EtBr, Sigma, USA)를 이용하여 염색한 후 UV 하에서 확인하였다.27) 6) SDS-polyacrylamide gel 전기영동 및 Western
blot analysis: 정상 및 시료가 처리된 배지에서 자란 세
포들을 lysis buffer로 용해한 후, 고속원심분리로 세포 내 잔사물을 분리시킨 다음 동량의 단백질을 SDS-polyacryl- amide gel 전기영동으로 분리하였다. 분리된 단백질을 함 유한 acrylamide gel을 nitrocellulose membrane으로 electro- blotting에 의해 전이시킨 후, 10% skim milk를 함유한 PBS-T (0.1% Tween 20 in PBS)에 4oC에서 1시간 이상 incubation하면서 비특이적인 단백질들에 대한 blocking을
실시하였다. 그리고 특정 단백질에 대한 항체를 membrane에 적용시켜 항원 항체 반응을 일으킨 후, PBS- T로 씻어내고 특정 항체에 대한 이차 항체 반응을 실시 한 후 ECL (Enhanced ChemiLuminoesence) 용액을 적용시킨 다음 X-ray film에 감광시켜 특정 단백질의 양을 분석하 였다. 본 실험에 사용된 항체들은 Santa Cruz Biotechnol- ogy Inc. (USA) 및 Calbiochem (Germany)에서 구입하였으 며, 이차 항체로 사용된 peroxidase-labeled donkey anti-rabbit immunoglobulin 및 peroxidase-labeled sheep anti-mouse immunoglobulin은 Amersham Corp. (USA)에서 구입하였 다.28)
3. 통계분석
대조군과 각 시료로부터 얻은 실험 결과들의 유의성 을 검정하기 위하여 분산분석(ANOVA)을 행한 후 p
<0.05 수준에서 Duncan's multiple range test를 실시하였 으며, 그 결과는 평균(Mean)±표준편차(Standard deviation, SD)로 표시하였다. 모든 통계 분석은 Statistic Analysis System (v9.1 SAS Institute Inc., NC, USA) 통계프로그램을 이용하여 처리하였다.
결과 및 고찰 1. 암세포성장 저해 효과
세포에 대한 키토산의 독성을 검토한 결과(결과제시 안함), 수용성 및 불용성 키토산은 LLC-PK1 (porcine renal epithelial cell) 정상세포에 대하여 수용성 키토산은 5 mg/
ml의 첨가농도까지는 99% 이상의 높은 생존율을 나타내 었다. 불용성 키토산은 1 mg/ml까지는 95% 이상의 높은 생존율을 나타내어 이 농도에서는 독성을 나타내지 않 았으나, 이 보다 높은 농도에서는 LLC-PK1 정상세포에 대한 독성이 나타났으므로 세포에 영향을 주지 않는 농 도 범위 내(수용성 키토산은 5 mg/ml 농도 이하, 불용성 키토산은 1 mg/ml 농도 이하)에서 암세포 성장 억제 효 과를 관찰하였다.
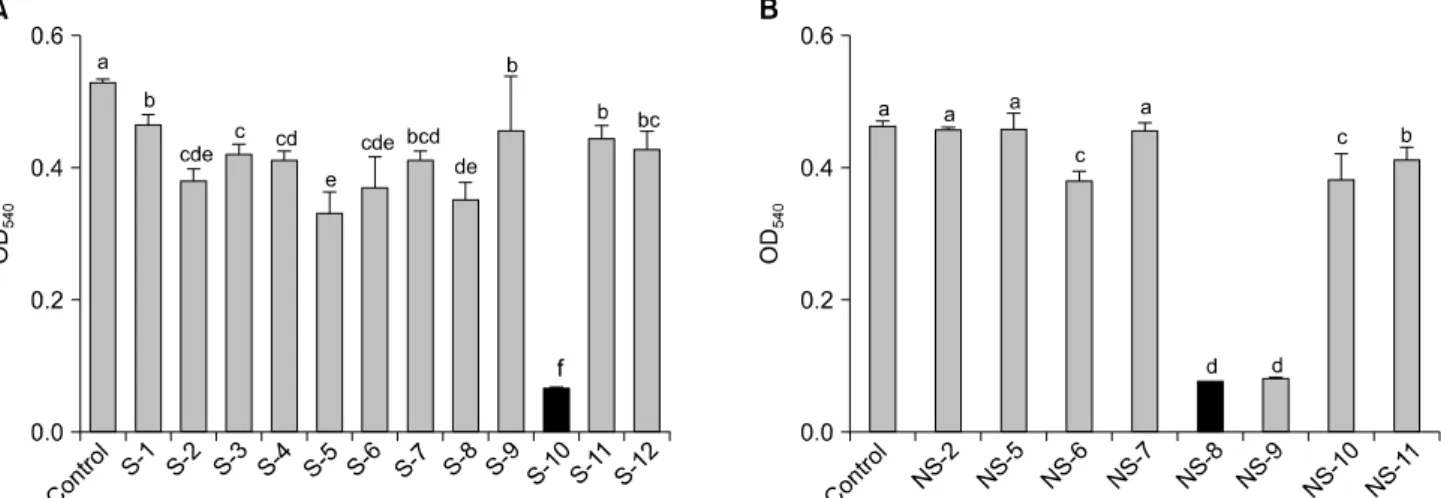
수용성 및 불용성 키토산의 암세포 성장 저해효과를 HT-29 인체 대장암세포를 이용하여 알아보았다. 수용성 키토산의 경우, 5 mg/ml의 높은 농도에서 S-10 (DA 90%, vis 5 cps)이 88%의 가장 높은 항암효과를 보였으며, 불용 성 키토산은 NS-8 (DA 95%, vis 22 cps)과 NS-9 (DA 100%, vis 8 cps)가 각각 83%로 높은 암세포 성장 저해 효과를 나타내었다(Fig. 1). 수용성 키토산은 탈아세틸화도가 90% 이상인 S-10의 효과가 가장 높았으며, 불용성 키토 산은 탈아세틸화도가 90% 이상이고 점도가 8∼22 cps로
Fig. 1. Inhibitory effect of various kinds of (A) soluble chitosan (5 mg/ml) and (B) non-soluble chitosan (1 mg/ml), on the growth of HT-29 human colon cancer cells in MTT assay. a∼fMeans with the different letters in the same column are significantly different (p<0.05) by Duncan's multiple test.
Fig. 2. Time-dependent growth inhibition by soluble (S-101) and non-soluble chitosan (NS-82) treatment in HT-29 human colon carcinoma cells. Cells were plated at 1×105 cells/60 mm plate, incubated for 24 hrs and treated with (A) 1 mg/ml (B) 2 mg/ml of chitosan for 48 hrs. The cells were trypsinized, wash with PBS and the viable cells were scored by hemocytometer counts.
Each point represents the mean±SD of three independent experiments. 1S-10: DA (degree of deacethylation) 90%, vis (viscosity) 3 cps, 2NS-8: DA 95%, vis 22 cps.
낮은 것이 효과가 좋았다. 본 실험에서 선택된 키토산의 분자량(Molecular weight: Mw)을 측정한 결과 S-10은 4,000
∼6,000, NS-8은 400,000∼600,000으로 나타나 불용성 키 토산의 분자량이 수용성에 비하여 매우 큰 것으로 나타 났다. 키토산 올리고당(3,000<Mw<5,000)이 대장암 세 포주에서 암세포의 성장을 억제한다는 선행연구는 본 실험의 수용성 키토산(S-10: 4,000<Mw<6,000)의 결과 와 비슷한 경향을 보였다.29) 또한 NS-8 (DA 95%, vis 22 cps)의 결과는 탈아세틸화도가 96% 이상이고 점도가 1.53 cps인 불용성 키토산이 혈액 종양세포에 대해서 효 과가 있다는 선행연구와 관련성이 있는 것으로 보인
다.30)
MTT assay 결과를 바탕으로 효과가 가장 높았던 S-10 과 NS-8을 이용하여 HT-29 인체 대장암 세포의 증식에 미치는 영향을 조사하기 위하여 trypan blue 시약을 이용 하여 시간과 농도에 따라 살아있는 세포수를 측정하였 다(Fig. 2). (A)와 (B)를 비교해 보면 알 수 있듯이 키토산 의 첨가 농도가 높을수록 암 세포 증식 억제 효과가 높 고, 불용성 키토산보다는 수용성의 암세포 증식률 및 생 존율이 높은 것으로 보아 불용성 키토산의 효과가 더 높 은 것을 알 수 있었다. 이는 앞서 행한 MTT 실험과 비슷 한 경향을 보인다(Fig. 1). 특히, NS-8은 1 mg/ml의 농도로
Fig. 3. Induction of apoptosis by soluble (S-101) and non-soluble (NS-82) chitosan treatment in HT-29 human colon carcinoma cells. Cells were incubated with soluble and non-soluble chitosan (1 mg/ml) for 48 hr and then stained with DAPI. After 10 min incubation at room temperature, the cells were washed with PBS and photographed with a fluorescence microscope using blue filter. Magnification, ×400. 1S-10: DA (degree of deacethylation) 90%, vis (viscosity) 3 cps, 2NS-8: DA 95%, vis 22 cps.
Fig. 4. Induction of Bax and inhibition of Bcl-2 by chitosan treatment in HT-29 human colon carcinoma cells. (A) RT-PCR: HT-29 cells were incubated with chitosan for 48 hr, Total RNA was isolated using an RNA Zol B reagent and RT-PCR was performed using indicated primers. The amplified PCR products were run in an 1% agarose gel and visualized by EtBr staining. GAPDH was used as a house-keeping control gene. (B) Western blot: Lysed and cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the anti-Bcl-2 and anti-Bax antibodies.
Proteins were visualized using ECL detection system. β-actin was used as a house-keeping control gene. CON: control, SC:
soluble chitosan (S-10), NSC: non-soluble chitosan (NS-8).
처리한 군에서는 24시간 후에 거의 모든 대장암 세포의 성장이 저해되었다. 또한, 높은 농도(2 mg/ml)에서는 S-10 과 NS-8 처리군이 12시간 만에 거의 모든 HT-29 세포의 성장을 저해하였으며, 따라서 S-10과 NS-8은 대장암 세 포의 증식을 억제할 뿐만 아니라 사멸효과가 있는 것으 로 나타났다.
2. Apoptosis 유도 효과
암세포의 성장억제 기전으로 중요하게 연구되고 있는 분야는 apoptosis에 의한 암세포의 세포사멸 기전이다.30) Apoptosis는 세포막과 소기관의 변화와 함께 빠른 세포 축소가 일어나 세포막에 크고 작은 돌기가 많이 생긴 후
불규칙하게 변형되면서 많은 apoptotic body가 생성된 다.31) 수용성 키토산과 불용성 키토산의 암세포 증식 억 제는 apoptosis 유발의 결과로 추측되므로 키토산에 의한 apoptosis 가능성을 조사하였다. DAPI는 DNA에 강력하 게 결합하는 형광염료이고 cell membrane에 손상을 주지 않고 통과하므로 살아있는 cell과 고정된 cell 모두에서 사 용될 수 있으며, 신속하게 cell로 들어가 DNA와 결합한 다.32) 이러한 이유로 DAPI staining을 통한 암세포 핵의 형태적 특징을 관찰하였다(Fig. 3). 정상 배지에서 자란 HT-29 대장암세포(control)는 모두 정상적인 핵의 형태를 띠고 있었으나, 키토산이 함유된 배지에서 자란 세포의 핵은 전형적인 apoptosis가 유발되었을 경우 관찰되는
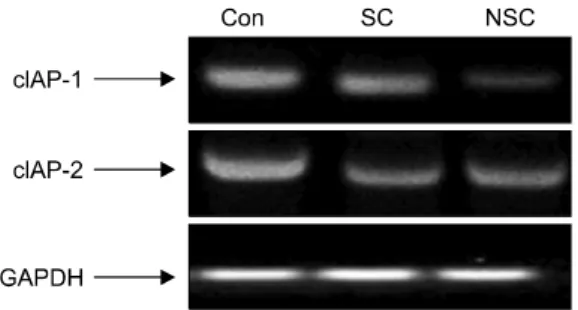
Fig. 5. Effect of chitosan on the expression of cIAP-1 and cIAP-2 mRNAs in HT-29 human colon carcinoma cells. Fol- lowing treatment with chitosan for 48 hrs, total RNAs were isolated and RT-PCR was performed using cIAP-1 and cIAP-2 primers. The amplified PCR products were run in agarose gel and visualized by EtBr staining. CON: control, SC: soluble chitosan (S-10), NSC: non-soluble chitosan (NS-8).
Fig. 6. Induction of p53 and p21 by chitosan treatment in HT-29 human colon carcinoma cells. (A) RT-PCR: HT-29 cells were incubated with chitosan for 48 hr, Total RNA was isolated using an RNA Zol B reagent and RT-PCR was performed using indicated primers. The amplified PCR products were run in an 1% agarose gel and visualized by EtBr staining. GAPDH was used as a house-keeping control gene. (B) Western blot: Lysed and cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the anti-p53 and anti-p21 antibodies. Proteins were visualized using ECL detection system. β-actin was used as a house-keeping control gene. CON: control, SC: soluble chitosan (S-10), NSC: non-soluble chitosan (NS-8).
apoptotic body의 출현을 관찰할 수 있었다. 특히 NS-8을 처리한 군에서 S-10을 처리한 군보다 apoptotic body의 출 현이 잘 관찰되었으며 앞서 행한 암세포 성장 저해 효과 실험의 결과와 비슷한 경향을 보였다. 따라서 키토산에 의한 암세포 성장 억제 중 한 가지 기작은 apoptosis 유발 에 의한 것으로 사료되며, 불용성 키토산(NS-8)이 수용성 키토산(S-10)에 비해서 높은 효과를 보였다.
3. Bcl-2와 Bax mRNA, protein 발현
Apoptosis 과정에는 세포외 인자들이 signal로서 전달되 어 관련유전자들의 발현과 조절에 관여하는 것도 알려 지고 있으며 많은 단백질이 apoptosis 기전에 관련되어 있 다. 특히 Bcl-2, Bax 등의 유전자에 의해 발현된 단백질은 서로 상호 작용하여 apoptosis를 유도하는 자극에 대한 세 포의 민감 정도를 조절 한다. Bcl-2는 anti-apoptotic 분자 로서 apoptosis의 유발을 억제하고, Bax는 pro-apoptosis 분 자로서 apoptosis 유발과 관련이 있다. 이들 유전자는 mitochondria로부터의 cytochrome c를 유리시켜 종양억제 유전자인 p53, caspase, DNA 단편화와 연관된 endonuclease 등의 활성을 조절한다.33,34) 키토산에 의한 apoptosis가 이 들 유전자의 발현 조절과 연관이 있는지를 조사하였다 (Fig. 4). mRNA 발현의 경우, Bax의 발현이 대조군에 비하 여 키토산을 처리한 군에서 증가되는 것을 볼 수 있는 반면, Bcl-2는 큰 차이를 보이지 않았다. 하지만 Western blot을 이용하여 단백질을 비교해 본 결과, 대조군에 비 하여 키토산을 첨가하였을 때 Bcl-2의 발현이 현저히 감 소하는 것을 알 수 있었으며, 특히 불용성 키토산이 수용 성에 비하여 낮은 발현 정도를 보였다. 따라서 키토산은 이들 유전자의 발현을 조절하여 apoptosis를 유도하여 암 세포의 증식을 억제한 것으로 나타났으며, 특히 수용성
키토산보다 불용성키토산이 효과가 높았다.
4. cIAP-1와 cIAP-2 mRNA 발현
IAP (inhibitor of apoptosis proteins) family는 apoptosis 유발 에 핵심적인 역할을 하는 caspase와 직접 결합하여 이들 의 활성을 억제한다.35) 따라서 키토산에 의한 인체 대장 암세포의 apoptosis 유발과 연관된 IAPs의 관련 여부를 조 사하였다(Fig. 5). IAP family 중 cIAP-2의 경우 대조군에 비하여 키토산에서 전사 수준의 유의적인 발현의 차이 를 관찰할 수 있었으나 수용성과 불용성간의 차이는 없 었다. cIAP-1의 경우, 대조군에 비하여 키토산에서 mRNA의 발현이 유의적으로 감소되었으며 특히 불용성 키토산의 발현 정도가 수용성에 비하여 현저히 감소하 였다. 따라서 키토산에 의한 cIAP-1의 발현 저하는 caspase의 활성화에 영향을 미쳐 apoptosis에 연관성이 있
는 것으로 생각되며, 이에 대한 지속적인 연구가 요구된 다.
5. p53와 p21 mRNA, protein 발현
키토산에 의한 암세포의 증식억제가 Cdk (cyclin depen- dent kinase) inhibitor의 발현과 상관성이 있는지의 여부를 조사하기 위하여 현재까지 알려진 Cdk inhibitor 중 많은 연구가 이루어져있는 p21과 세포증식 조절에 중요한 조 절인자에 해당하는 종양억제인자인 p53의 발현에 미치 는 키토산의 영향을 Western blotting 및 RT-PCR법으로 조사하였다(Fig. 6). mRNA와 단백질 발현 모두에서 대조 군에 비하여 p53이 유의적으로 증가되는 것을 관찰할 수 있었으며, 특히 불용성 키토산(NS-8)이 수용성(S-10)에 비 하여 현저하게 증가하였다. p21의 경우, 수용성 키토산 의 mRNA 발현이 불용성에 비하여 높은데 반하여, 단백 질의 발현정도는 서로 비슷하였다. 이 실험의 결과로 키 토산 처리에 의한 대장암 세포의 증식 억제는 p53 및 p21의 유전자가 관여하는 것을 확인할 수 있었다. 최근 의 연구에서 p21은 세포주기 상 G1 기의 조절뿐만 아니 라 G2/M 기를 포함한 세포주기 전반에 걸친 조절자로 서, 세포증식과 분화, 노화 등에 관여하는 중요한 조절인 자로 인식되어져 오고 있다.36∼39)
결 론
키토산의 항암 기능성의 차이를 확인하기 위해 탈아 세틸화도 및 점도가 다양한 키토산(수용성 12가지, 불용 성 11가지)의 암세포 성장 저해 정도를 확인하고, 그 중 가장 효과가 높은 키토산의 항암 기전을 밝히고자 하였 다. 키토산의 암세포 성장 저해효과를 HT-29 인체 대장 암세포를 이용하여 알아본 결과, S-10 (수용성 키토산, DA 90%, vis 5 cps, Mw: 4,000∼6,000)과 NS-8 (DA 95%, vis 22 cps, Mw: 400,000∼600,000, 불용성 키토산)의 효과 가 높았다. 수용성 및 불용성 키토산 중 항암 기능성이 높은 것은 탈아세틸화도가 90% 이상인 것으로 특히 불 용성 키토산은 점도가 8∼22 cps로 낮은 것이 효과가 좋 았다. DAPI 염색을 통하여 키토산에 의한 암세포의 apoptotic body 출현을 관찰할 수 있었으며, anti-apoptotic 분자인 Bcl-2와 caspase와 결합하여 apoptosis를 저해하는 인자인 cIAP-1 발현이 감소하는 것을 알 수 있었다. 특히 NS-8에 의해 Bcl-2와 cIAP-1의 발현이 현저히 감소하였 다. 또한 키토산 처리에 의해 종양억제 인자인 p53이 증 가되었으며, 특히 NS-8에 의해서 현저하게 증가하였다.
따라서 키토산은 apoptosis 관련인자인 Bcl-2와 Bax 및
cIAP-1의 발현과 cell cycle에 관여하는 p53, p21을 조절하 여 암세포의 생존 및 증식을 저해시킨 것으로 사료된다.
감사의 글
이 논문은 부산대학교 자유과제 학술연구비(2년)에 의 하여 연구되었으며 이에 감사드립니다.
참 고 문 헌
1) Doll R, Peto R. The causes of cancer: quantitative estimates of avoidable risks of cancer in the United States today. J Natl Cancer Inst 66, 1191-1308, 1981.
2) Ornstein DL, Pipas JM, Rigas JR. Recent advances in cancer chemotherapy. Acad Radiol 6, 696-705, 1999.
3) Lee KY, Lee YC, Park YS, Yoon KH, Kim BS. A study of relation between dietary vitamin A intake and serum vitamin A levels and cancer risk in Korea. Korean J Nutr 18, 301-312, 1985.
4) Watson RR, Leonard TK. Selenium and Vitamin A, E and C; nutrients with cancer preventive properties. J Am Diet Assoc 86, 505-510, 1986.
5) Ahn BZ, Lee YH, Kim SI. Isolation of cytotoxicity potentia- ting substances from red ginseng. J Korean Cancer Assoc 24, 795-806, 1992.
6) Hirose A, Sato E, Fujii H, Sun B, Nishioka H, Aruom OI.
The influence of active hexose correlated compound (AHCC) on cisplatin-evoked chemotherapeutic and side effects in tumor-bearing mice. Toxicol Appl Pharmacol 222, 152-158, 2007.
7) Min SH, Pyo MY. Effect of chitosan on the cytotoxicity of anticancer drugs in vitro. J Environ Toxicol 22, 263-269, 2007.
8) Kim SK, Jeon YJ. Bioactivities of chitin and chitosan(2)- antitumor activities by immunological function of chitin, chitosan and their oligosaccharides. J Chitin Chitosan 2, 3-15, 1997.
9) Jeon YJ, Lee EH, Kim SK. Bioactivities of chitin and chitosan (1)-antimicrobial function, hypertension control function and cholesterol control function. J Chitin Chitosan 1, 4-14, 1996.
10) Sugano M, Watanabe S, Kishi A, Izume M, Ohtakara A.
Hypocholesterolemic action of chitosan and other polysaccha- rides. Lipids 23, 187-191, 1988.
11) Nishimura K, Nishimura S, Azuma I. Immunological activity of chitin and its derivatives. Vaccine 2, 93-99, 1984.
12) Peluso G, Petillo O, Ranieri M, Santin M, Ambrosio L, Calabro D, Avallone B, Balsamo G. Chitosan-mediated sti- mulation of macrophage function. Biomaterials 15, 1215-1220, 1994.
13) Tokoro A, Tatewaki K, Suzuki T, Mikami S, Suzuki M.
Growth inhibitory effect of hexa-N-acetyl chitohexaose and chitohexaose against meth A solid tumor. Chem Pharm Bull 36,
784-790, 1988.
14) Suzuki K, Mikami T, Okawa Y, Tokoro A, Suzuki S, Suzuki M. Antitumor effect of hexa-N-acetyl-chitohexaose and chitohexaose. Carbohydr Res 151, 403-408, 1986.
15) Qi L, Xu Z. In vivo antitumor activity of chitosan nano- particles. Bioorg Med Chem Lett 16, 4243-4245, 2006.
16) Youn SK, Kim YJ, Ahn DH. Antioxidative effects of chitosan in meat sausage. J Korean Soc Food Sci 30, 477-481, 2001.
17) Lee HY, Park MS, Ahn DH. Effect of storage properties of pork dipped in chitosan solution. J Korean Soc Food Sci 32, 519- 525, 2003.
18) Kim BK, Rhee SH, Park KY. Chemopreventive effects of chitosan added kimchi. J Korean Assoc Cancer Prevent 9, 162- 1702, 2004.
19) Li H, Cho JY, G TC, Choi CR, Lee KD, Cho JE, Cho GS, Ham KS. Increment of physiologically active compounds in germinated brown rice treated with chitosan and its effect on obesity of rat fed a high fat diet. J Korean Soc Food Sci 37, 985-991, 2008.
20) Kim HK, Lee JS, Kim SK, Cho SY, Choi HG, Lee EH. In vitro adsorption of radiostrontium by various phosphorylated chitosans by changing the molecular weight, the degree of deacetylation and the degree of substitution of phosphated groups. J Chitin Chitosan 2, 45-56, 1997.
21) Yi SD, Yang JS, Chang KS, Oh MJ. Viscometric and pulsed photostimulated luminescence properties of irradiated glutinous rice. J Food Sci & Nutr 9, 133-137, 2004.
22) Vold IM, Christensen BE. Periodate oxidation of chitosans with different chemical compositions. Carbohydr Res 340, 679- 84. 2005.
23) Park JG, Kramer BS, Steinber CJ, Collins JM, Minna JD, Gazdar AF. Chemosensitivity testing of human colorectal carcinoma cell lines using a tetrazolium-based colorimetric assay. Cancer Res 47, 5875-5879, 1987.
24) Skehan P, Storeng R, Monks SA, McMahon J, Vistica D, Warren JT, Bokesch H, Kenney S, Boyd MR. New colori- metric cytotoxicity assay for anticancer-drug screening. J Natl Cancer Inst 82, 1107-1112, 1990.
25) Miller J. Experiments in molecular genetics, cold spring harbor laboratory. New York, Cold Spring Harbor, 113-185, 1972.
26) Choi YH, Baek JH, Yoo M, Chung H, Kim ND, Kim KW.
Induction of apoptosis by ursolic acid though activation of caspases and down-regulation of c-IAPs in human prostate epithelial cells. Int J Oncol 17, 565-571, 2000.
27) Chomczynski P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques 15, 532-537, 1993.
28) Hempelmann E, Schirmer RH, Fritsch G, Hundt E, Gröschel- Stewart U. Studies on glutathione reductase and methemo- globin from human erythrocytes parasitized with Plasmodium falciparum. Mol Biochem Parasitol 23, 19-24, 1987.
29) Nam KS, Cho HJ, Shon YH. Effect of chitosan oligosaccha- ride on chemopreventive enzymes of colon cancer. J Chitin Chitosan 11, 171-174, 2006.
30) Jeong YW, Kim KS, Oh JY, Park JC, Lee JC, Yoo K, Kim JW, Chang SJ, Baek WK, Suh SI, Suh MH, Cho JW.
Chitosan-oligosaccharides inhibits HL-60 cell growth through induction of apoptosis. J Chitin Chitosan 5, 74-78, 2000.
31) Kokal WA. The impact of antitumor therapy on nutrition.
Cancer 55, 273-278, 1985.
32) Lee KI, Park KY. The effects of anticarcinogenic activity of several green-yellow vegetables. J Life Science 3, 143-152, 1993.
33) Antonsson B. Bax and other pro-apoptotic Bcl-2 family 'killer proteins' and their victim the mitochondrion. Cell Tissue Res 306, 347-361, 2001.
34) Zamzami N, Kroemer G. The mitochondrion in apoptosis:
how Pandora's box opens. Nat Rev Mol Cell Biol 2, 67-71, 2001.
35) Kil JH. Studies on Development of Cancer preventive and Anticancer Kimchi and Its anticancer mechanism. Department of Food Sceince and Nutrition, Ph D, Pusan National University. 2004.
36) Elledge SJ, Harper JW. Cdk inhibitors: on the threshold of checkpoints and development. Curr Opin Cell Biol 6, 847-852, 1994.
37) Harper JW, Adami GR, Wei N, Keyomarsi K, Elledge SJ.
The p21 Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell 75, 805-816, 1993.
38) El-Deiry WS, Harper JW, O'Connor PM, Velculescu VE, Canman CE, Jackman J, Pietenpol JA, Burrell M, Hill DE, Wang Y, Wiman KG, Mercer WE, Kastan MB, Kohn KW, Elledge SJ, Kinzler KW, Vogelstein B. WAF1/CIP1 is induced in p53-mediated G1 arrest and apoptosis. Cancer Res 54, 1169-1174, 1994.
39) Zeng YX, El-Deiry WS. Regulation of p21WAF1/CIP1 expression by p53-independent pathways. Oncogene 12, 1557- 1564, 1996.