DOI: 10.5532/KJAFM.2013.15.4.245
ⓒ Author(s) 2013. CC Attribution 3.0 License.
CO 2 및 기온 상승이 ‘후지’/M.9 사과나무의 광합성, 신초생장 및 과실품질에 미치는 영향
권헌중1·사공동훈1*·박무용1·송양익1·정경호1·남종철2·한점화2·도경란2
1농촌진흥청 국립원예특작과학원 사과시험장,
2농촌진흥청 국립원예특작과학원 과수과
(2013년 9월 3일 접수; 2013년 11월 19일 수정; 2013년 12월 17일 수락)
Influence of Elevated CO 2 and Air Temperature on Photosynthesis, Shoot Growth, and Fruit Quality of ‘Fuji’/M.9 Apple Tree
Hun-Joong Kweon1, Dong-Hoon Sagong1*,Moo-Yong Park1, Yang-Yik Song1, Kyeong-Ho Chung1, Jong-Chul Nam2, Jeom-Hwa Han2
and Gyung-Ran Do
21Apple Research Station, National Institute of Horticultural & Herbal Science, R.D.A., Gunwi 716-812, Korea
2Fruit Research Division, National Institute of Horticulture & Herbal Science, Rural Development Administration, Suwon 440-706, Korea
(Received September 3, 2013; Revised November 19, 2013; Accepted December 17, 2013)
ABSTRACT
This study was conducted to find out the influence of elevated atmospheric CO2 concentrations and air temperature on photosynthesis and fruit quality of ‘Fuji’/M.9 apple trees and to investigate these to the effects of climate change during the last four years (2009-2012). The treatments employed were:
‘Ambient’ (ambient temperature + ambient CO2 concentration); ‘High CO2’ (ambient temperature + elevated CO2 concentration); ‘High Temp.’ (elevated temperature + ambient CO2 concentration); and
‘High CO2 + High Temp.’ (elevated temperature + elevated CO2 concentration). The elevated temperature plots were maintained at 4oC higher than ambient air temperature, while the elevated CO2 plots were maintained at 700
µmol·mol
−1. Annual treatment period was applied from end of April to beginning of November for four years. Results showed that elevated CO2 decreased stomatal conductance and leaf SPAD value, but increased photosynthetic rate, intercellular CO2 concentration (Ci), and starch content of mesophyll tissue. In the vegetative growth, elevated temperature increased total number of shoot and total shoot growth per tree, but elevated CO2 decreased average shoot length. In the fruit quality, elevated CO2 increased soluble solid content, fruit red color, and ethylene production. In conclusion, elevated CO2 increased photosynthetic rate of apples during the early growth, but effect of increased photosynthetic rate due to elevated CO2 was decreased during latter growth stage. Elevated temperature, on the other hand, tended to decrease photosynthetic rate of apples during the early growth, but that tended to increase during latter growth stage. Both elevated CO2 and temperature tended to decrease the degree of decreased photosynthetic rate due to each factor.Key words: fruit red color, intercellular CO2 concentration, Malus domestica Borkh., starch content of mesophyll tissue, stomatal conductance
* Corresponding Author : Dong-Hoon Sagong ([email protected])
I. 서 론
식물체의 광포화점은 기온과 CO2 농도에 따라 크게 변하는데, 생육 적온까지 기온이 높아질수록 광합성속 도는 높아지나 광포화점은 낮아지고, 또 CO2포화점까 지 공기 중의 CO2농도가 높아질수록 광합성속도와 광포화점은 높아진다(Chae et al., 2006). 사과나무는 15-35oC 범위에서 광합성능력의 대부분이 이루어지나 (Palmer et al., 2003), 기온이 30oC 이상 되면 체내 수분을 유지하기 위하여 기공이 닫히고, 호흡소비 (respiratory cost)가 많아지면서(Lakso et al., 1999;
Oh et al., 2004), 과실 비대 및 품질이 감소될 수 있다(Lakso, 2003; Kim et al., 2006; Tomana, 1983).
광합성속도가 30oC에서부터 감소되는 것은 광합성속도 와 정의 상관관계를 가지는 기공전도도가 29oC를 넘 으면 감소되기 때문이라고 한다(Lee et al., 2012;
Massonnet et al., 2007; Sagong et al., 2013; Tromp, 2005; Yoon and Richter, 1991).
대기 중 CO2 농도가 높아지면 식물들의 광합성과 건물생산성이 증대하는데(Byun et al., 2006; Chae et al., 2006), 이는 세포 내 CO2 농도(intercellular CO2 concentration, Ci)가 높아졌기 때문이라고 한다 (Kim and Lee, 2001a). 그러나 CO2농도가 높은 조 건에서 생육이 장기화되면 생육 후기 광합성이 오히려 억제될 수 있다(Kim and Lee, 2001b).
한편 기공 바로 아래 세포간극의 CO2 농도가 높아 지면 기공이 닫히고, 반대로 낮아지면 기공이 열리는 데(Byun et al., 2006), Leymarie et al.(1999)은 대 기 중의 CO2농도가 2배로 상승되면 식물체에 삼투 스트레스(osmotic stress)가 발생하여 기공전도도가 낮 아진다고 하였다.
사과 과총잎(spur leaves)의 광합성속도는 발아기(4 월 20일)부터 급격히 증가하여 첫 홍뢰기(first pink stage)인 5월 1일경에 최고치에 달한 이후 줄어들어 큰 변화 없이 지속되다가 10월초부터 잎의 노화에 의 해 급격히 감소하기 시작하여 11월초에 잎이 황화되면 서 광합성활동을 하지 않았다는 보고(Fujii and Kennedy, 1985)가 있다. 생장이 끝난 후 잎의 광합성능력은 보통 엽록소의 파괴, 엽록체의 광인산화작용의 감퇴, 광합성 효 소인 Rubisco (ribulose 1, 5-bisphosphate carboxylase/
oxygenase)의 분해 및 활성 저하 등에 의해 감소되는 데(Kim et al., 2006), 생육기 중에 저온 및 서리가
발생하면 광합성속도가 감소되는 등 잎의 노화가 촉진 된다(Seeley and Kammereck, 1977; Spencer and Titus, 1972). 그러나 생육 후기에 질소를 추비하거나 기온을 높이면 잎의 노화를 지연시킬 수 있다(Lakso and Lenz, 1986; Kim et al., 2006)
일반적으로 대기 중의 CO2 농도가 높아지면 광합성 활동이 활발해져 잎의 전분이 많아지지만, 생육 적온 이상의 고온에서는 호흡작용이 왕성해져 양분의 소모 가 많아진다고 알려져 있다(Lee et al., 2009). 그러 나 Chae et al.(2006)은 대기 중 CO2 농도를 0.3- 1.0%로 증가시키면 콩의 경우 떡잎에서의 엽록소 함 량의 증가를 볼 수 있으나 탄산가스처리와 함께 조명 시간을 길게 하는 경우에는 생장한 잎의 엽록소 함량 이 감소하고, 오이는 생육 후반에 잎의 노화가 진행되 어 빨리 시든다고 하였다.
기온이 높아지면 수체생장은 증가하고, 대기 중의 CO2 농도가 증가되면 식물의 탄소 흡수, 광합성, 수분 이용효율, 지상부 및 지하부의 생장이 증가한다고 알 려져 있다(Chae et al., 2006; Chung et al., 2012;
Huang et al., 2007). Ro et al.(2001)은 기온을 상 승시킨 곳에서 자란 사과나무의 주간직경 및 신초의 길이와 수는 대기온도 처리구들보다 높았고, 기온 및 CO2 농도를 동시에 높였던 처리구가 처리구들 중에서 가장 높았다고 하였다. 그러나 사과나무는 수분 유지 가 어려워지면 feedforward 기작으로 기공을 닫아 수 분의 손실을 최소화하면서 신초생장을 억제시킨다 (Lakso, 2003; Yoon and Richter, 1991; Yoon et al., 2005). Sagong et al.(2013)은 일 평균기온이 30oC를 넘었던 날이 10일이나 발생했던 해의 신초길이가 하루 도 없었던 해보다 짧았다고 하였다.
과실의 성숙은 대체로 20-25oC를 중심으로 하여 이 보다 낮으면 늦어지고, 약간 높아 27oC 전후가 되면 빨라지며, 30oC 이상이 되면 과실의 성숙은 오히려 늦어지는 것으로 알려져 있다(Kim et al., 2006). 또 한, Chae et al.(2006)은 CO2를 시용하면 수체생육이 지연되면서 과실의 성숙이 지연되고, CO2를 시용하였 을 때 과실이 커지면 상대적으로 가용성 고형물 함량 이 저하된다고 하였다. 그러나 Lee et al.(2006)은 CO2농도 상승구들의 과중은 대기 CO2농도 처리구 들보다 낮았으며, 에틸렌 발생량은 CO2농도 상승구 들이 더 높았다고 하였고, Han et al.(2012)은 CO2
농도 상승구들의 과중, 가용성 고형물 함량, 착색 정
도가 대기 CO2 농도처리구들보다 높았다고 하였다.
산업혁명 이전(1790년대)의 대기 중 CO2 농도는 세계적으로 약 280µmol·mol−1이었으나 1999년에는 376µmol·mol−1로 높아졌고(Park et al., 2005), 현재 기상청(Korea Meteorological Administration, KMA) 에서 조사한 안면도의 2011년 연 평균 CO2 농도는 396µmol·mol−1이라고 한다. 이러한 추세라면 2100년 에는 CO2 농도가 약 2배(540-970µmol·mol−1)로 높아 지고, 기온은 지금보다 1.4-6.0oC 정도 상승될 것으로 예측되고 있다(Chung et al., 2007; IPCC, 2007;
Kim and Lee, 2001b). Oh et al.(1994)은 CO2 농 도와 기온이 상승하게 되면 우리나라의 연강수량은 약 15-30% 증가(약 300-400mm)할 것으로 예측하였고 특히, 여름철 강우량이 증가할 것이라고 하였다. 이와 같이 최근 널리 논의되고 있는 지구온난화는 광합성능 력에 영향을 주는 기후요소(광량, CO2농도, 기온, 상 대습도)를 변화시킬 것으로 예상되는데, 국내에서는 월 별 및 시간 별로 변화하는 기후요소에 따른 사과나무 광합성능력의 변화를 조사한 보고가 없다.
따라서 본 시험에서는 CO2 농도 및 기온상승이 사 과나무의 생리생태에 미치는 영향을 구명하고자, 기후 변화 정부간 위원회(Inter-governmental Panel on Climate Change, IPCC)의 미래 기후변화 시나리오들 (Special Report on Emissions Scenarios, SRES)중 CO2 농도가 약 2배(720µmol·mol−1)로 높아지면, 우리 나라 2100년의 평균기온은 지금보다 4.0oC 정도 상승 될 것이라는 A1B 시나리오(IPPC, 2007)를 기초로 하여, 기상환경이 조절 가능한 시설에서 ‘후지’/M.9 사과나무의 광합성능력, 신초생장 및 과실품질을 2009 년부터 2012년까지 4년간 조사하였다.
II. 재료 및 방법
2.1. 시험재료 및 관리방법
본 시험은 경기도 수원시 소재 국립원예특작과학원 의 기상환경 조절이 가능한 라이조트론에서 실시하였 다. 시험에 사용된 사과나무는 접목부 상단 10cm 부 위의 주간직경이 14-15mm, 수고는 2.0-2.2m, 30cm 이상 측지수가 5-10개인 ‘후지’/M.9 1년생 묘목이었다.
재식방법은 2009년 4월초에 식토, 육묘용 상토, 퇴비 를 5:3:2 비율로 혼합한 토양을 90L 포트에 채우고, 각 포트마다 1년생 묘목 1주를 대목이 25±1cm 정도
노출되게 심었다.
처리구는 대기온도+대기 CO2 농도(Ambient), 대기 온도+CO2 농도 700µmol·mol−1 (High CO2), 대기온 도보다 4oC 상승+대기 CO2농도(High Temp.), 대기 온도보다 4oC 상승+CO2 농도 700µmol·molσ1(High CO2+High Temp.)로, 포트 1개씩 배치하였다. 라이조 트론의 CO2 농도 및 기온 처리기간은 4개년 모두 4 월말부터 11월초까지로, 내부의 CO2 농도와 기온은 외부에 설치된 데이터로거(21X, Campbell Scientific, USA)에서 실시간으로 수집된 기상정보를 바탕으로 적외 선 가스 분석기(ZRH model, Fuji Electric CO., Japan) 와 중앙 분산식 제어시스템(POREX6800, POSCON Co., Korea)으로 조절하였다(Ro et al., 2001). 다만 기온 상승구들의 내부 기온은 40oC를 넘지 않도록 조 절하였고, 모든 처리구들의 상대습도는 인위적으로 조 절하지 않았다.
시험수 관리방법에 있어, 수형은 세장방추형을 목표 로 하였고, 시험 2년 차(2010년)부터 매년 2월말에 주당 복합비료 100g(N:P:K=12:6:8)을 시비하였다. 복 토는 2년 차에 대목이 15±1cm 정도 노출되게 하였 다. 관수방법은 1년 차(2009년)의 경우 3일 간격으로 살수분무기를 이용하여 토양수분장력이 0kPa에 도달할 정도로 충분히 관수하였고, 2년 차부터는 토양수분장 력계(Watermark, Irrometer, USA)를 포트 지면에서 30cm 깊이에 설치하여 포트의 토양수분장력이 -50kPa 이하로 내려가지 않도록 관수하였다. 착과량은 2년 차 의 경우 3월말의 주간황단면적(trunk cross sectional area, TCA)을 기준으로 하여 5월 중순에 1cm2당 4과 로 조절하였고, 3년 차부터 주당 15개 정도 착과가 되 게 조절하였는데, 3년 차에 ‘Ambient’와 ‘High Temp.’
처리구들에서는 꽃이 생성되지 않았다.
시험기간 동안 수원지역의 일별 최고, 평균, 최저기 온은 2009년만 기상청의 수원 관측소(수원시 권선구 서둔동 관측소)의 자료를 이용하였고, 이외 연도는 국 립원예특작과학원에 설치되어 있는 자동기상장치 (Automatic weather system, Campbell Scientific, USA)로 관측된 자료를 이용하였다.
2.2. 평균 광합성능력
기온 및 CO2 농도에 따른 생육기 동안의 사과나무 의 광합성능력을 비교하고자, 기상청의 일기예보를 참 고하여 일사량이 충분한 날을 택하였다. 1차년도(2009
년)에는 시험구별로 5개의 정단신초(지면에서 약 1.3m 높이)를 미리 선정하고, 7월부터 10월까지 매달 10일 전 후에 인공광 챔버가 부착된 광합성 측정기(LI-6400, LI- COR, USA)로 중간 부위 잎의 광합성속도, 기공전도도 및 세포 내 CO2 농도(intercellular CO2 concentration, Ci)를 6시부터 18시까지 1시간 간격으로 조사하였다.
각 처리구의 광합성 측정 시 광량 조절은 매시간 휴 대용 광량 측정기(Field scout, Spectrum, USA)로 측정한 노지의 광량을 LI-6400의 인공광 챔버에 매시 간 대입하였고, 광합성 측정 시 CO2 농도 조절은
‘Ambient’ 및 ‘High Temp.’ 처리구들의 경우 매시간 LI-6400의 CO2농도 조절 밸브를 완전히 열어 측정 한 노지 대기의 CO2농도를 매시간 LI-6400에 대입 하였다. ‘High CO2’과 ‘High CO2+High Temp.’ 처 리구들은 700µmol·mol−1로 고정하여 광합성능력을 조 사하였다.
2차년도(2010년)에는 5월 하순에 처리구별로 5개의 과대지(지면에서 약 1.3m 높이)를 선정하고, 6월부터 10월까지 매달 10일 전후에 중간 부위 잎을 대상으로 일출 때(광량 30µmol·m−2·s−1 이상)부터 일몰 직전(광 량 30µmol·m−2·s−1 이하)까지 1시간 간격으로 광합성 능력(광합성속도, 기공전도도, Ci)을 조사하였다. 측정 방법은 2009년과 동일하였다.
3차년도(2011년)에는 ‘Ambient’와 ‘High Temp.’
처리구에 과실이 없어, 5월 하순에 각 처리구별로 7개 의 정단신초(지면에서 약 1.3m 높이)를 선정하고 6월 부터 10월까지 매달 20일 전후에 중간 부위 잎을 대 상으로 광합성능력을 9시부터 12시까지 1시간 간격으 로 3회 조사하였다. 측정방법은 2009년과 동일하였다.
4차년도(2012년)에는 5월 하순에 처리구별로 지면에 서 약 1.3m 높이 이하에 있는 3개의 과대지와 3개의 정단신초를 선정한 뒤, 6월부터 10월까지 2주 간격으 로 중간 부위 잎을 대상으로 광합성능력을 10시부터 14 시까지 1시간 간격으로 4회 측정하였는데, 처리구에 과 실이 없을 경우에는 꽃이 달린 흔적이 있는 신초를 대 상으로 측정하였다. 측정방법은 2009년과 동일하였다.
광합성능력의 통계분석은 연도별 평균치(6-10월의 평 균치, 2009년은 7-10월의 평균치)를 1반복으로 하였다.
2.3. 광합성능력의 일변화
기후요소들의 일변화에 따른 광합성능력의 일변화 조사는 월별 하루 동안 기상요소의 변화에 따른 광합
성능력 변화를 살펴보고자 2차년도(2010년)에 조사한 자료를 이용하였다.
2.4. 광합성능력의 시기별 변화
CO2농도 및 기온 상승에 따른 시기별 광합성 능 력의 변화 및 sink의 유무에 따른 광합성속도를 비교 하고자 4차년도(2012년)에 조사한 자료를 이용하였다.
과대지와 정단신초 간의 광합성속도 비교는 착과가 정 상적으로 된 ‘Ambient’ 처리구와 ‘High CO2’ 처리구 를 대상으로 하여 과대지와 정단신초 잎의 광합성속도 를 비교하였다.
2.5. 잎의 특성
대기 중의 CO2농도 상승에 따른 사과 잎의 노화 발생 여부 및 그 시기를 밝히고자, 본 시험에서는 잎 의 녹색 정도를 나타내는 SPAD 값 및 질소함량을 4 개년 동안 매년 10월 중순에 포트 지표면에서 1.5m 이상 높이에 있는 정단신초의 중간 부위 잎을 처리구 별로 20개씩 채취하여 조사하였다. 2012년에는 6월말 에 처리구별로 정단신초의 중간 부위 잎을 10개 선정 하여 SPAD 미터(SPAD-502, Minolta, Japan)를 이용 하여 6월말부터 10월말까지 2주 간격으로 잎의 SPAD값을 측정하였다.
10월경 잎의 SPAD 값과 질소함량의 통계분석은 연 도별 측정치를 1반복으로 한 4반복이었고, 시기별 잎 의 SPAD값은 1개의 측정치를 1반복으로 한 10반복 이었다.
엽육조직 내 전분함량은 2010년 7월초와 11월초 16시에 시험구별로 5개의 과대지 중간 부위에서 1매씩 5매를 채취하여 Lee et al.(2009)의 방법으로 관찰하 였다.
2.6. 과실품질 및 에틸렌 발생량
시험구별 나무당 화총수는 4월 중부터 5월초 사이 에 조사하였고, 시험구별 과중, 가용성 고형물 함량 및 산 함량, 착색 정도, 전분지수 등의 과실품질은 11 월초에 과실 별로 조사하였다. 전분조사는 Yoo et al.(2006)의 조사방법에 따라 0(미숙)-6(완전소실)까지 7단계로 구분하였다. 과실의 에틸렌 발생량은 2012년 수확(11월초) 당시 시험구별로 4과씩 선정하여, Kweon et al.(2010)의 방법으로 조사하였다. 통계처리 는 각 연도별로 평균치와 표준오차로 나타내었다.
2.7. 수체생장
주간비대량은 매년 생장 개시 전(3월)과 생장 종료 후(11-12월)에 접목부 상단 10cm에서의 줄기직경을 방향을 바꾸어 2회 측정 평균하여 TCA 차이를 cm2 으로 표시하였다. 신초생장은 매년 11월초에서 12월말 사이에 전체 신초를 대상으로 모두 조사하였다. 주간 횡단단면적 증가, 신초 생장의 통계분석은 연도별 측 정치를 1반복으로 하여 4반복으로 하였다.
III. 결과 및 고찰
3.1. 기상환경
4년(2009-2012년)동안 수원지역의 4월 25일부터 10 월 31일까지의 평균기온은 20.5oC, 평균 최고기온은 25.7oC, 평균 최저기온은 15.8oC 정도였고, 기온상승구 들(‘High Temp.’, ‘High CO2+High Temp.’)은 4.0oC가 높은 24.5oC, 29.7oC, 19.8oC이었다. 특히, 2012년에 는 예년과 달리 4월 30일(라이조트론 가동일)부터 5월 5일(개화 후 7일)까지 수원지역의 일 최고기온이
26oC(기온상승구들: 30oC)를 넘었던 날이 3일(5월 1 일: 28.2oC, 5월 2일: 27.1oC, 5월 3일: 27.8oC) 연 속으로 있었다(Fig. 1).
Oh et al.(2004)은 사과나무의 광합성 최적온도를 28- 30oC로 추정하였고, 32-35oC 이상이 되면 기공이 닫히 면서 광합성속도가 감소된다고 하였다. 4년 동안 수원지 역에서 일 최저기온이 21oC를 넘어 기온상승구들에서 열대야가 발생한 날(일 최저기온이 25oC를 넘은 날)은 평균적으로 49일 정도 있었고, 일 평균기온이 26oC를
Fig. 1. Change of maximum, average, and minimum air temperature at Suwon region in 2009 (A), 2010 (B), 2011 (C), and 2012 (D).
Table 1. Accumulated days of specific air temperature from 25 April to 31 October in Suwon region for 4 years
Year
Accumulated days of specified temperature Over 21oC
at Min. Temp.
Over 26oC at Avg. Temp.
Over 31oC at Max. Temp.
2009 43 15 24
2010 60 33 28
2011 47 21 16
2012 45 30 27
AVERAGE 49 25 24
넘었던 날(기온상승구들: 30oC)은 평균적으로 25일 정 도, 일 최고기온이 31oC를 넘었던 날(기온상승구들:
35oC)은 평균적으로 24일 정도 있었다(Table 1).
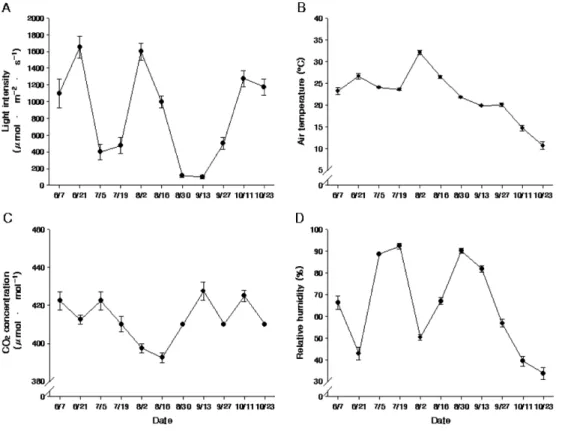
광합성 일변화를 측정한 2010년에 평균광량이 가장 높았던 날은 6월 4일로 1,800µmol·m−2·s−1까지 올라갔 고, 가장 낮았던 날은 비가 와 흐렸던 8월 12일로 400µmol·m−2·s−1밖에 올라가지 않았다(Fig. 2A). 광합성 측정 시 평균기온이 가장 높았던 날은 7월 8일과 9월 14일로 최고기온은 28.4oC 정도였고, 기온이 가장 낮았 던 날은 10월 7일로 최고, 최저기온은 각각 23.0oC, 9.8oC였다(Fig. 2B). 대기 중 평균 CO2 농도가 가장 높았던 날은 10월 7일로 8시까지 500µmol·mol−1를 유 지하였고, 가장 낮았던 날은 8월 12일로 9시부터 20시 까지 390µmol·mol−1에서 변화가 없었다(Fig. 2C). 대기 중 평균 상대습도가 가장 높았던 날은 비가 온 8월 12 일로 평균 상대습도가 80%를 넘었고, 가장 낮았던 날 은 광량이 가장 높았던 6월 4일로 36.7%까지 낮아졌다 (Fig. 2D).
2012년에 비가 와서 평균 광량이 500µmol·m−2·s−1 을 넘지 못했던 흐린 날은 7월 5일, 7월 19일, 8월 30
일, 9월 13일이었고, 광량이 1,000µmol·m−2·s−1 이상이었 던 맑은 날은 6월 7일, 6월 21일, 8월 2일, 8월 16일, 10월 11일, 10월 23일이었다(Fig. 3A). 광합성측정 시 평균기온은 8월 2일에 32.1oC까지 높아졌다가 10월 23 일에 10.7oC까지 떨어졌다(Fig. 3B). 대기 중 평균 CO2
농도는 6월 7일부터 7월 5일까지 420µmol·mol−1 이상을 유지하다가 8월 16일에 392µmol·mol−1까지 떨어졌고, 이후로 다시 410µmol·mol−1 이상을 유지하였다(Fig.
3C). 평균 상대습도는 흐린 날의 경우 80% 이상을 유지 하였고, 맑은 날에는 10월 이전까지 40% 이상을 유지하 다가 10월 이후로 40% 이하로 떨어졌다(Fig. 3D).
이상의 결과를 종합해보면, 대기온도 처리구들 (‘Ambient’, ‘High CO2’)에서는 수분스트레스가 발생 할 정도로 기온이 높지 않았으나 기온상승구들(‘High Temp.’, ‘High CO2+High Temp.’)에서는 매년 49일 이상 열대야가 발생하고, 일 평균기온이 30oC를 넘었 던 날이 매년 25일 정도 발생하는 등 고온 스트레스 가 발생할 수 있는 여건이 되었다(Table 1). 특히, 예 년과 달리 2012년 개화기에는 기온상승구들의 최고 기온이 30oC를 넘어 낙화 및 착과 불량이 발생할 위 Fig. 2. Diurnal change of light intensity (A), air temperature (B), CO2 concentration (C), and relative humidity (D) of Suwon region in 2010.
Fig. 3. Seasonal change of light intensity (A), air temperature (B), CO2 concentration (C), and relative humidity (D) of Suwon region in 2012. Vertical bars indicate standard errors of four-hour (10:00-14:00) replication.
Table 2. Average of photosynthetic rate, stomatal conductance, transpiration rate, leaf-to-air water vapor pressure deficit on leaf temperature (VpdL), and intercellular CO2 concentration (Ci) of ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature for 4 years
Treatmentz Photosynthetic ratey (µmol·m−2·s−1)
Stomatal conductance (mol·m−2·s−1)
Ci (µmol·mol−1)
Ambient 10.3 bcx 0.16 a 257 b
High CO2 12.7 a 0.11 b 422 a
High Temp.
0 8.9 c
0.14 ab 255 bHigh CO2 + High Temp. 11.8 ab 0.11 b 429 a
ANOVAw
CO2 (A) * Ns ***
Temp. (B) Ns Ns Ns
A × B Ns Ns Ns
zAmbient (ambient temperature + ambient CO2); High CO2 (ambient temperature + 700µmol·mol−1 CO2); High Temp., (4oC higher than ambient temperature + ambient CO2); High CO2+ High Temp., (4oC higher than ambient temperature + 700µmol·mol−1 CO2). Sample leaf for checking photosynthetic rate in 2009 and 2011 was the terminal shoot leaf and that of 2010 was bourse shoot leaf. That of 2012 was both the terminal shoot leaf and bourse shoot leaf.
yFour-year average value (2009-2012); one replicate was from one-year average (June to October).
xMeans followed by the same letter are not significantly different using Duncan's multiple range test, P=0.05.
wCorrelation between: CO2 and all parameters tested except between Ci; air temperature and all parameters tested;
and CO2 and air temperature, were not significant (Ns) at P=0.05, 0.05(*), 0.001(***).
험이 있었는데(Fig. 1D), Lee(2008)는 사과 개화기 기 온이 15-30oC 범위에 있을 때에는 화분의 발아와 신 장에 문제가 없으나 35oC까지 올라가면 화분이 발아되 지 않아 수정이 불가능할 수 있다고 하였다.
3.2. 평균 광합성능력
4년간의 평균 광합성능력을 비교해 본 결과(Table
2), 광합성속도는 CO2 농도 상승구들(‘High CO2’,
‘High CO2+High Temp.’)이 대기 CO2 농도 처리구 들(‘Ambient’, ‘High Temp.’)보다 높았으나, 기공전도 도는 반대로 CO2 농도 상승구들이 대기 CO2 농도 처 리구들보다 낮은 경향이 있었다. 그러나 요인분석에 있어서는 광합성속도와 Ci만 CO2 농도 요인에서 유의 차가 인정되었을 뿐, 기공전도도는 CO2 농도 및 기온
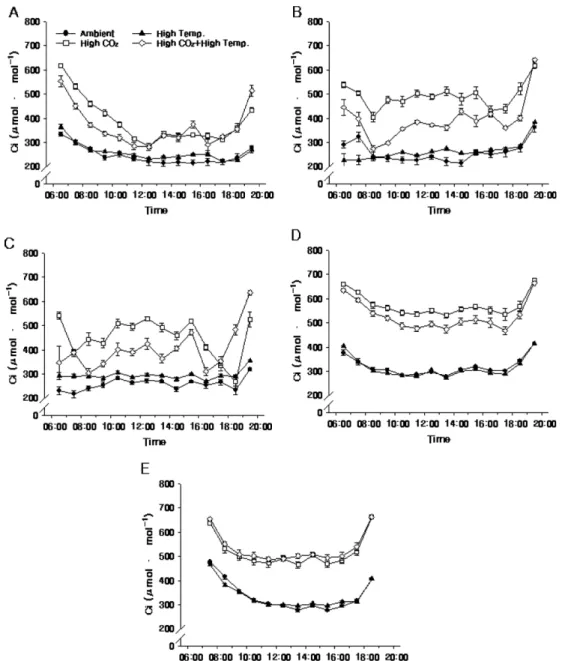
Fig. 4. Diurnal change of photosynthetic rate of ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature on 4 June (A), 8 July (B), 12 August (C), 14 September (D), and 7 October (E) in 2010. Vertical bars indicate standard errors using five-leaf replications.
요인에 영향을 받지 않는 것으로 나타났다.
본 시험에서 CO2 농도 상승구들의 기공전도도가 대 기 CO2 농도 처리구들보다 낮았음에도 불구하고 광합 성속도가 더 높을 수 있었던 것(Table 2)은 CO2 농 도 상승구들은 우선적으로 세포 내 CO2를 소비하면서 광합성을 해도 세포 내에 잔존해 있는 CO2 농도가 대기 CO2 농도 처리구들보다 높아(Table 2), 기공이 닫혀 CO2를 유입하지 않아도 세포 내 CO2를 광합성
에 사용할 수 있었기 때문으로 추정되었다(Byun et al., 2006; Kang et al., 2007). 이는 마치 사막지역 의 식물이 한낮에 기공을 닫은 채 저녁에 흡수한 CO2를 이용하여 광합성을 하는 것(Byun et al., 2006;
Park et al., 2002)과 같아 보였다. 한편 ‘High Temp.’
처리구의 광합성속도가 가장 낮았던 것(Table 2)은 일 평균기온이 30oC를 넘는 날이 많으면서 Ci가 CO2 농 도 상승구들보다 적어(Table 1 and 2), 30oC 이상의
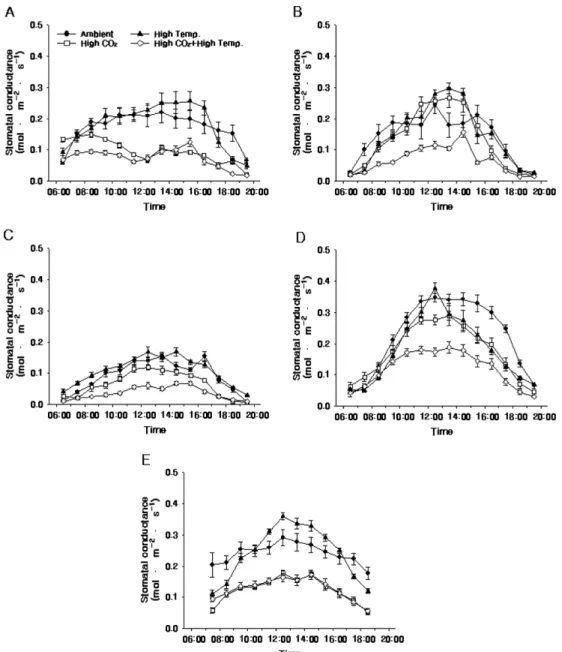
Fig. 5. Diurnal change of stomatal conductance of ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature on 4 June (A), 8 July (B), 12 August (C), 14 September (D), and 7 October (E) in 2010. Vertical bars indicate standard errors using five-leaf replications.
고온에 의한 호흡소비가 다른 처리구들보다 많았기 때 문으로 생각되었다(Lakso et al., 1999; Oh et al., 2004; Sagong et al., 2013).
3.3. 광합성능력의 일변화
2010년 처리구들간 광합성능력 일변화의 차이(Fig.
4, 5, and 6)는 4년 동안의 평균치의 차이(Table 2)
와 비슷하였다. 특히, Ci는 광량에 영향을 받았는데, 광량이 높아 광합성속도가 높아진 시기에 낮아졌다가 광량이 100µmol·m−2·s−1 이하가 되는 해질녘(19-20시, 10월 7일: 18-19시)에 다시 높아졌고, 광량이 1,800 µmol·m−2·s−1까지 올라갔던 6월 4일 12-14시경 CO2
농도 상승구들의 Ci는 280-320µmol·mol−1까지 내려갔 다(Fig. 2A and 6).
Fig. 6. Diurnal change of intercellular CO2 concentration (Ci) of ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature on 4 June (A), 8 July (B), 12 August (C), 14 September (D), and 7 October (E) in 2010. Vertical bars indicate standard errors using five-leaf replications.
처리구들간의 광합성속도 차이는 6월 4일부터 CO2
농도 상승구들이 자연상태의 CO2 농도 처리구들보다 높아지기 시작하여(Fig. 4A), 7월 8일에 처리구들간 광합성속도 차이는 최대가 되었는데, 7월 8일경 CO2
농도 상승구들의 하루 평균 광합성속도는 11.2- 13.0µmol·m−2·s−1로 자연상태의 CO2 농도 처리구들의 8.6-9.4µmol·m−2·s−1 대비 19-51% 정도 높았고, 처리 구들간의 광합성속도는 ‘High CO2’, ‘High CO2+High Temp.’, ‘Ambient’, ‘High Temp.’ 처리구 순으로 낮 아졌다(Fig. 4B).
비가 온 8월 12일에 CO2 농도 상승구들의 하루 평 균 광합성속도는 6.2-7.6µmol·m−2·s−1 정도로, 대기 CO2 농도 처리구들의 5.3-6.2µmol·m−2·s−1 대비 0- 43% 정도 높았고, 7월 8일 대비 55-67% 수준이었다 (Fig. 4C). 9월 14일 처리구들의 하루 평균 광합성속 도는 7월 8일의 83-104% 수준으로, ‘High CO2’ 처리
구만이 7월 8일 대비 83% 수준으로 떨어졌을 뿐
‘Ambient’, ‘High Temp.’, ‘High CO2+High Temp.’ 처 리구들은 7월 8일과 비교해 거의 동일하였다(Fig. 4D).
일조시간이 2시간 줄어든 10월 7일(Fig. 2A)에 CO2 농도 상승구들의 하루 평균 광합성속도는 11.1- 12.2µmol·m−2·s−1 정도로 대기 CO2 농도 처리구들의 10.6-10.9µmol·m−2·s−1 대비 1-15% 정도 높았는데, 그 차이는 2010년 조사기간 중에서 가장 낮았다. 또 한, 10월 7일 ‘Ambient’ 처리구의 광합성속도 하루 총합은 7월 8일 대비 비슷하였으나, ‘High Temp.’
처리구는 오히려 증가하였고, ‘High CO2+High Temp.’ 처리구는 15%, ‘High CO2’ 처리구는 20%
정도 감소되었다(Fig. 4B and 4E). 즉, 2010년에 CO2 농도 상승구들은 생육후기에 광합성속도가 감소되 는 잎의 노화증상이 나타났다(Chae et al., 2006;
Kim and Lee, 2001b).
Fig. 7. Seasonal change of photosynthetic rate (A), stomatal conductance (B), and Ci (C) of ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature in 2012. Checking time per day was between 10:00 and 14:00. Vertical bars indicate standard errors of six-leaf replication.
3.4. 광합성능력의 시기별 변화
비가 와서 평균 광량이 500µmol·m−2·s−1을 넘지 못 했던 흐린 날보다는 광량이 1,000µmol·m−2·s−1 이상이 었던 맑은 날에 처리구들간 광합성능력의 차이가 뚜렷하 게 나타났다(Fig. 3 and 7). 광량이 1,000µmol·m−2·s−1 이상이었던 맑은 날 처리구들의 광합성속도 변화 양상 을 살펴보면, 전반적으로 시간이 지날수록 광합성속도 가 감소되는 경향을 나타내었는데, ‘Ambient’, ‘High CO2’, ‘High CO2+High Temp.’ 처리구들의 10월 11 일 광합성속도는 8월 16일에 비해 감소되었던 반면에
‘High Temp’ 처리구는 오히려 높아졌다(Fig. 3A and 7A). 기공전도도는 처리구들 모두 8월 16일 이후로 감소되는 경향이었다(Fig. 7B). Ci는 대기 CO2 농도 처리구들의 경우 6월초부터 10월말까지 Ci는 258- 361µmol·mol−1 범위에 있었으나, CO2농도 상승구들 은 6월 21일까지 353-407µmol·mol−1 범위에 있다가 6월 21일 이후로 411-585µmol·mol−1 범위로 높아졌다 (Fig. 3A and 7C).
2012년 과대지와 정단신초 잎의 시기별 광합성속도 비교에 있어(Fig. 8), 과대지 잎의 광합성속도가 정단 신초 잎보다 높은 경향이 있었고, 특히, 흐린 날보다 는 맑은 날에 차이가 뚜렷하게 나타났다(Fig. 3A and 8). 이는 과대지 잎이 정단신초의 잎보다 sink(과실)와 의 거리가 가까웠기 때문으로 생각되었다(Byun, et al., 2006; Fujii and Kennedy, 1985).
Fujii and Kennedy(1985)는 사과 잎의 광합성속도 는 시간이 지날수록 잎의 노화가 진행되어 감소된다고
하였다. 본 시험 역시 시간이 지날수록 시험구들의 광 합성속도와 기공전도도가 낮아지는 경향을 보였다(Fig.
7). 그러나 대기 CO2 농도 처리구들의 생육 후기(10월 11일) 광합성속도는 생육초기(6월 21일) 광합성속도 대 비 18-23% 정도 감소되었던 반면에 CO2 농도 상승 구들은 23-34% 정도 감소될 정도로 CO2 농도 상승 구들에서 생육 후기 광합성 감소 정도가 대기 CO2
농도 처리구들보다 더 컸고, 생육 후기에 기온이 높았 던 ‘High Temp’ 처리구의 10월 광합성속도는 다른 처리구들과 달리 8월보다 증가하거나 혹은 생육 후기 광합성속도 감소 정도가 적었다(Fig. 7A). 이는 CO2
농도가 높은 조건에서 생육이 장기화되면 생육 후기의 광합성이 억제된다는 보고(Kim and Lee, 2001b) 및 광합성능력은 엽령과 함께 감소되지만 착과지나 적심 된 가지의 잎, 질소 추비 또는 고온조건이면 광합성능 력 감소시기가 늦어질 수 있다는 보고(Lakso and Lenz, 1986; Kim et al., 2006)와 일치하는 결과를 보였다.
3.5. 잎의 특성
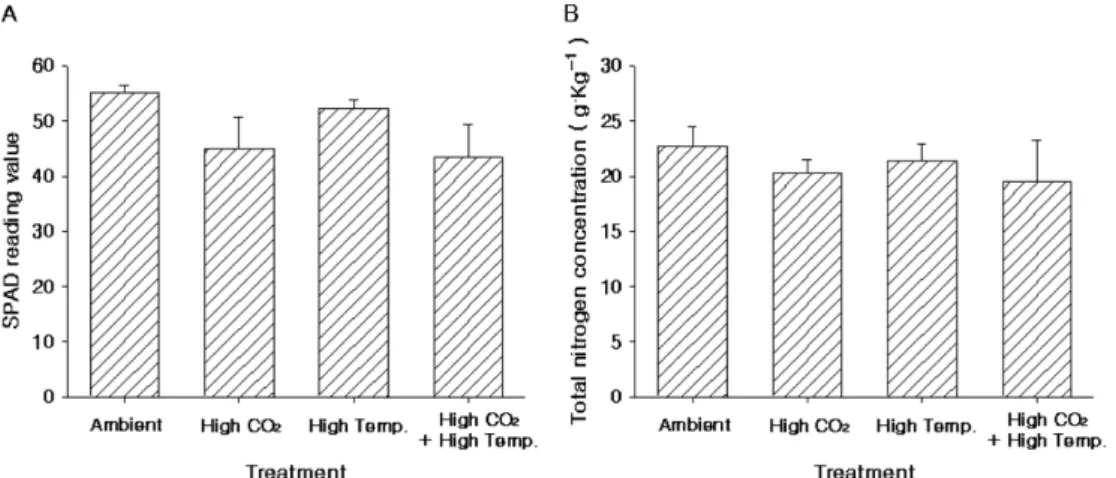
10월의 잎 특성에 있어, 4년 동안 잎의 평균 SPAD값 은 CO2 농도 상승구들이 43-45 (SPAD reading value) 정도로 자연상태의 CO2 농도 처리구들의 52-55(SPAD reading value)보다 낮았다(Fig. 9A). 4년 동안 잎의 평균 질소함량은 19.5-22.7g·kg−1로 CO2 농도 상승구 들이 낮은 경향이 있었으나 유의 차는 없었다(Fig.
9B). 2012년 시기별 SPAD값은 6월말(6월 20일)부터
Fig. 8. Seasonal change of photosynthetic rate on bourse shoot leaf and terminal shoot leaf of ‘Fuji’/M.9 apple tree under the
‘ambient’ treatment (A) ‘High CO2’treatment (B) in 2012. Checking time per day was between 10:00 and 14:00. Vertical bars indicate standard errors of three-leaf replication.
8월 중(8월 15일)까지 CO2 농도 및 기온 상승구들이
‘Ambient’ 처리구보다 높게 유지되다가, 8월 중 이후 부터 CO2 농도 상승구들이 ‘Ambient’ 처리구보다 낮 아졌는데 특히, ‘High CO2+High Temp.’ 처리구가 급격하게 낮아졌다. ‘High Temp.’ 처리구는 7월 18 일부터 ‘Ambient’ 처리구보다 높게 유지되었다(Fig.
10). 이는 대기 중 CO2 농도를 상승시킨 곳에서 식물 이 장기간 생육할 경우 생육 후반에 잎의 엽록소 함 량이 줄거나 혹은 노화가 촉진되어 빨리 시든다는 보 고(Chae et al., 2006)와 동일하였다. 또한, 2012년
‘High CO2+High Temp.’ 처리구의 SPAD값이 8월 중 이후로 급격하게 감소되었던 것(Fig. 10)은 2012 년에 ‘High CO2+High Temp.’ 처리구에서는 sink(과 실)가 없어 잎의 노화가 촉진되었기 때문(Byun et al., 2006)으로 추정되었는데, 이러한 결과(Fig. 10)는 재식 당년인 2009년 10월에 CO2 농도 상승구들만 조 기낙엽 현상이 발생했던 것(자료 미제시)과 비슷한 현 상이었다.
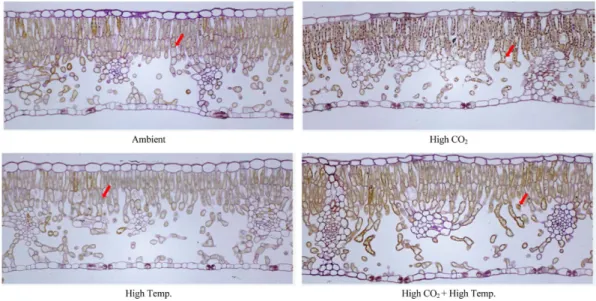
2010년 7월초에 채취한 과대지엽의 조직에서는 CO2 농도 상승구들이 대기 CO2 농도 처리구들보다 전 분(검은색 점)이 많았는데, 특히 ‘High Temp.’ 처리 구와 비교하였을 때 전분의 축적은 온도보다는 CO2
농도에 더 영향을 받는 것으로 관찰되었다(Fig. 11).
이는 대기 중의 CO2 농도가 높아지면 잎의 전분이 많아진다는 보고(Lee et al., 2009)와 동일하였다. 그 러나 11월초에는 ‘Ambient’ 처리구보다 ‘High Temp.’ 처리구의 전분함량이, ‘High CO2’ 처리구보다
‘High CO2+High Temp.’ 처리구의 전분함량이 더 높 았다(Fig. 12).
본 시험에서 기온상승구들의 11월초 잎의 전분 함 량이 대기온도 처리구들보다 다소 높았던 것(Fig. 12) 은 사과나무는 10월경에 잎의 노화가 시작되면서 광합 성능력이 감소되나(Fujii and Kennedy, 1985), 가을철 에 사과나무 주변 온도를 높여주면 잎의 노화를 지연 시킬 수 있다는 보고(Lakso and Lenz, 1986) 및 2012년 ‘High Temp.’ 처리구의 6월말(6월 21일) 광 합성속도 대비 10월말 광합성속도 감소 정도는 24.5%로 다른 처리구들의 광합성속도 감소 정도(32.6- 42.8%)보다 낮았던 결과(Fig. 7A)와 2012년 ‘High Fig. 9. SPAD reading value (A) and total nitrogen contents (B) of leaf in October of ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature, from 2009-2012. Vertical bars indicate standard errors of four-year replications.
Fig. 10. Seasonal change of SPAD reading values of ‘Fuji’/
M.9 apple tree under elevated CO2 concentration and air temperature in 2012. Vertical bars indicate standard errors of ten-leaf replication.
Temp.’ 처리구의 생육 후기 SPAD값이 ‘Ambient’ 처 리구보다 높게 유지되었던 결과(Fig. 10)를 미루어 보 아, 2010년 기온상승구들의 잎의 노화가 정상기온 처 리구들보다 늦게 진행되었기 때문으로 생각되었다.
3.6. 과실품질 및 에틸렌 발생량
시험구들의 과중, 산 함량 및 전분지수는 해에 따라 달리 나타났으나 가용성 고형물 함량과 착색은 CO2
농도가 상승할 경우 높아지는 것으로 나타났다. 2012 년 과실의 에틸렌 발생량과 호흡속도는 ‘High CO2’ 처리구가 가장 높았다(Table 3).
본 시험에서 ‘High CO2’ 처리구의 가용성 고형물 함량과 착색이 다른 처리구들보다 높았던 것(Table 3) 은 ‘High CO2’ 처리구의 광합성속도가 다른 처리구 들보다 높았기 때문(Table 1; Fig. 4 and 7A)으로 생각되었다(Han et al., 2012). 또한, 기온상승구들의 Fig. 11. Starch contents of mesophyll tissue of bourse shoot’s leaf in ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature on 7 July 2010. Arrows indicate starch of mesophyll tissue.
Fig. 12. Starch contents of mesophyll tissue of bourse shoot’s leaf in ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature on 3 November 2010. Arrows indicate starch of mesophyll tissue.
착색 정도가 대기온도 처리구들보다 낮았던 것(Table 3)은 잎으로부터 전류 되는 광합성산물에 의해 생성되 는 과피의 안토시아닌은 만개 후 146일부터 190일(4 월 20일을 만개기로 정할 경우 9월 중순에서 10월말) 까지의 평균기온이 14-15oC일 때 가장 많이 생성된다 는 보고(Lee, 1999) 및 4년 동안 수원의 9-10월(과실 성숙기)의 평균기온이 17.4oC 정도(기온상승구들:
21.4oC)였던 결과(Fig. 1)를 미루어 보아, 기온상승구 들의 과실 성숙기 평균기온이 21.0oC를 넘어 안토시 아닌의 생성이 억제되었기 때문으로 생각되었다(Saure, 1990).
한편 2011년 대기 CO2 농도 처리구들에 꽃이 거 의 없었고(Table 3), 이러한 현상은 2013년에 동일한 처리구들에서 다시 발생하였다(자료 미제시). 이는 재 배포트의 크기가 광합성활동에 영향을 미친다는 보고 (Kim and Lee, 2001b) 및 CO2 농도 상승구들의 광 합성속도와 잎 내 축적된 전분이 대기 CO2 농도 처 리구들보다 높았던 결과(Table 1; Fig. 11 and 12)를
미루어 보아, 본 시험의 포트가 나무에 비해 작은데다 가 대기 CO2 농도 처리구들의 수체 내 축적된 동화 산물이 CO2농도 상승구들보다 적었기 때문으로 추정 되었다. 또한, 2012년 기온상승구들에서 낙화가 심했 던 것(Table 1)은 개화 후 1주일 동안 기온상승구들 의 최고기온이 30oC 이상 올라가는 날이 많아(Fig.
1D) 정상기온 처리구들에 비해 화분 발아 및 화분관 신장에 문제가 발생했을 가능성이 있을 것으로 생각되 었다(Lee, 2008).
3.7. 수체생장
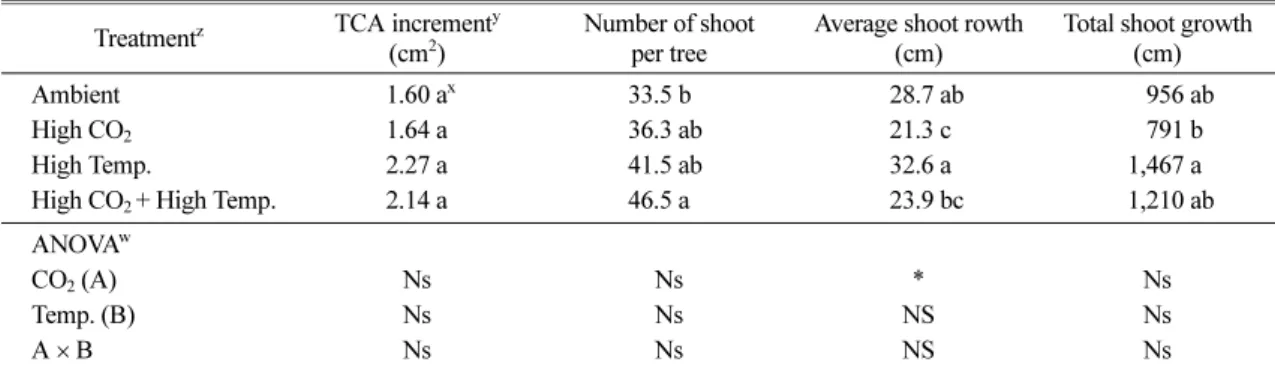
4년 동안 시험구들의 연 평균 TCA 비대량은 1.60- 2.14cm2로 시험구들간에 차이가 없었다. 나무당 총 신 초수는 ‘High CO2+High Temp.’ 처리구가 41.5개로 가장 많았고, 평균 신초장과 총 신초장은 ‘High Temp.’ 처리구가 각각 32.6cm, 1,467cm로 가장 길었 다. 전반적으로 기온상승구들의 수체생장이 대기온도 처리구들보다 높은 경향이었다. 요인분석에 있어서는 Table 3. Number of flower cluster per tree, fruit quality, ethylene production and respiration rate of ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature for 3 years
Treatment
zNo. of flower cluster per
tree
yNo. of fruit per
tree
xFruit weight
x(g)
Soluble solid
x(°Bx)
Titratable acidity
x(%)
Fruit red color
x(Hunter a)
Starch pattern index (0-6)
Ethylene production
w( µL·kg
−1·hr
−1)
Respiration rate
w(CO
2mL·kg
−1·hr
−1) In 2010
vAmbient 27 11 365±15.0
u10.7±0.11 0.38±0.009 -3.1±1.44 - - -
High CO
217 10 295±19.8 12.3±0.36 0.36±0.009 8.2±2.44 - - -
High Temp. 16 14 365±18.7 10.9±0.19 0.35±0.008 -1.5±1.24 - - -
High CO
2+High Temp. 10 7 238±13.8 12.0±0.43 0.35±0.005 -9.7±0.65 - - -
In 2011
vAmbient 1 - - - - - - - -
High CO
218 16 331±17.7
u12.3±0.43 0.37±0.014 0.9±1.91 1.4±0.26 - -
High Temp. 1 - - - - - - - -
High CO
2+High Temp. 17 15 296±20.4 13.9±0.34 0.41±0.016 -3.0±1.63 0.5±0.13 - - In 2012
vAmbient 36 16 342±19.5
u12.4±0.17 0.37±0.014 9.3±1.34 0.4±0.12 0.13±0.034 13.3±0.28 High CO
231 18 386±14.1 14.4±0.08 0.37±0.008 15.0±2.06 1.4±0.15 0.22±0.015 16.4±0.26 High Temp. 14 4 307±26.6 12.7±0.25 0.37±0.248 6.4±2.38 0.7±0.25 0.22±0.059 12.1±0.90
High CO
2+High Temp. 11 - - - - - - - -
zSee the footnote of Table 1.
yNumber of flower cluster per tree observed from the middle of April to the early of May.
xFruit qualities observed on each fruit in the early of November; Starch pattern index values are from 0-6, with 0=immaturity.
wEthylene production observed on four fruit per treatment in the early of November.
vIn 2010, treatment was applied on 4 fruits/cm2 TCA; In 2012, ‘High Temp.’ and ‘High CO2 + High Temp.’ treated trees was flower abscission critically from the end of April to the early of May.
uMean ± standard errors.
평균 신초장만 CO2 농도 요인에서 유의 차가 인정되 었을 뿐, TCA 비대량, 총 신초수 및 총 신초생장량 은 CO2 농도 및 기온 요인에 영향을 받지 않는 것으 로 나타났다(Table 4).
사과나무의 신초생장은 기온이 높을수록 증가되지만 30oC 이상의 고온조건이 장기화되면 감소되는 것으로 알 려져 있다(Johnson and Lakso, 1985; Lakso, 2003;
Sagong et al., 2013). 그러나 본 시험에서는 열대야 및 30oC 이상의 고온이 자주 발생한 기온상승구들의 신초생장이 대기온도 처리구들보다 왕성했다(Table 1 and 4). 이는 포트의 토양수분장력이 -50kPa 이하로 내려가지 않도록 충분히 관수 해주었기 때문이거나 혹 은 개화기 고온(Fig. 1)에 의해 기온상승구들에서 낙 화가 심하게 발생되면서 기온상승구들의 착과량이 대 기온도 처리구들보다 적어져 신초생장이 왕성해졌기 때문(Forshey and Elfving, 1989)으로 생각되었다 (Table 3 and 4). 즉, CO2 농도 상승구들의 평균 신 초생장이 대기 CO2 농도 처리구들보다 짧았던 것 (Table 4)은 CO2 농도 상승구들의 착과수가 대기 CO2 농도 처리구들보다 많았기 때문으로 판단되었다 (Table 3).
IV. 요약 및 결론
이상의 결과를 종합해보면, CO2농도 상승은 생육 초기에는 광합성을 크게 증가시켰으나 후기에는 광합
성 효율이 감소되었다. 반면에 기온상승은 생육 초기 30oC 이상의 고온과 낮은 상대습도에 의해 광합성이 감소되는 경향이 있었으나(Oh et al., 2004), 후기에 는 광합성을 높게 유지시키는 경향이 있었다(Fig. 4, 7A, and 8). CO2농도 상승구들의 광합성 효율이 생 육 후기에 감소된 것(Fig. 4, 7A, and 8)은 CO2농 도 상승이 잎의 노화를 촉진시켰기 때문(Chae et al., 2006; Kim and Lee, 2001b)으로 생각되었다(Fig. 9 and 10). 기온상승구들의 광합성능력이 생육 후기에 높게 유지될 수 있었던 것(Fig. 4 and 7A)은 기온상 승구들의 생육 후기 기온이 대기온도 처리구들보다 높 아 잎의 노화를 어느 정도 지연시켰기 때문(Fig. 9, 10, and 12)으로 생각되었다(Lakso and Lenz, 1986;
Kim et al., 2006).
한편 30oC 이상의 고온조건이 장기화되면 사과나무 는 체내 수분유지를 위해 기공을 닫아 광합성속도를 감소시키는데(Lakso, 2003), 본 시험에서는 ‘High Temp.’ 처리구가 고온기인 7, 8월에 그러한 경향이 있었다(Fig. 4 and 7A). 그러나 기온과 CO2 농도를 동시에 높인 ‘High CO2+High Temp.’ 시험구는 기 공을 적게 열어도 세포 내 CO2농도가 높아 고온에 서도 광합성속도를 높게 유지할 수 있었다(Table 2;
Fig. 4, 5, and 7). 즉, CO2 농도 및 기온 중 어느 한 요인만 높아진다면 사과나무는 수분스트레스 혹은 노화 촉진에 의해 광합성능력이 감소될 수 있지만 두 요인이 동시에 높아지면 광합성 억제현상이 줄어들 수
Table 4. TCA increment and shoot growth of ‘Fuji’/M.9 apple tree under elevated CO2 concentration and air temperature for 4 years
Treatmentz TCA incrementy (cm2)
Number of shoot per tree
Average shoot rowth (cm)
Total shoot growth (cm)
Ambient 1.60 ax 33.5 b 28.7 ab
,0956 ab
High CO2 1.64 a 36.3 ab 21.3 c
0,791 b
High Temp. 2.27 a 41.5 ab 32.6 a 1,467 a
High CO2 + High Temp. 2.14 a 46.5 a 23.9 bc 1,210 ab ANOVAw
CO2 (A) Ns Ns * Ns
Temp. (B) Ns Ns NS Ns
A × B Ns Ns NS Ns
zSee footnote on Table 1.
yFour-year average value (2009-2012); one replicate was from one-year average. TCA increment was difference between TCA in March and that of December in every year; Total shoot growth was checked on the end of December.
xMeans followed by the same letter are not significantly different using Duncan's multiple range test, at P=0.05.
wCorrelation between: CO2 and all parameters tested; air temperature and all parameters tested; and CO2 and air temperature, were not significant (Ns) at P= 0.05, 0.05(*).
있다고 생각되었는데, 이는 지구온난화에 따른 CO2
농도 상승이 식물의 생육후기 광합성을 억제시키는 요 인으로 작용한다면, 기온의 상승은 CO2농도 상승에 따른 광합성 억제 현상을 완화시키는 요인으로 작용한 다는 보고(Kim and Lee, 2001b)와 일치하였다.
과실품질에서 과중과 가용성 고형물 함량은 CO2 농 도에 영향을 받는 경향이 있었고(Table 3), 착색은 CO2 농도가 높아지면 과실의 에틸렌 발생량이 증가되 어 hunter a값이 높아지지만, 기온이 상승되면 CO2
농도가 높아져도 hunter a값이 감소되는 것으로 나타 났다(Table 3). 즉, CO2 농도 상승은 과실의 성숙을 촉진시켜 수확시기를 앞당길 필요가 있었던 반면에 기 온상승은 착색을 감소시키므로 착색 증진을 위해 수확 시기를 저온이 올 때까지 연장시킬 필요가 있는(Park et al., 2011) 등, 지구온난화에 의해 CO2 농도 및 기 온이 상승할 경우 ‘후지’에서 가장 문제가 되는 것은 수확시기 및 저장성의 변동이라고 생각되었다.
신초생장은 기온 상승구들이 대기온도 처리구들보다 높은 것으로 나타났는데(Table 4), 이는 토양수분장력 이 충분하여 수분스트레스가 발생하지 않았기 때문이 거나 혹은 기온 상승구들의 착과량이 대기온도 처리구 들보다 적어져 신초생장이 왕성해졌기 때문(Table 3 and 4)으로 생각되었다(Forshey and Elfving, 1989).
따라서, 향후 CO2 농도 및 기온상승에 따른 사과나 무의 노화 혹은 수분스트레스 발생 여부를 밝히기 위 해서는 광합성반응과 더불어 수분퍼텐셜 및 수체 내 호르몬을 같이 조사해야 하며, 사과나무의 수체생장 및 과실품질을 조사하기 위해서는 처리구별로 착과가 안정된 뒤에 관련처리를 한다면 더 명확한 기대결과를 도출할 수 있는 것으로 생각되었다.
적 요
본 시험은 기후변화의 영향과 관련하여 기온 및 CO2 농도 상승이 ‘후지’/M.9 사과나무의 광합성 및 과실품질에 미치는 영향을 알아보고자 지난 4년(2009- 2012)동안 시험이 이루어졌다. 처리구들은 ‘Ambient’
(대기온도+대기 CO2 농도), ‘High CO2’(대기온도+상 승 CO2 농도), ‘High Temp.’(상승온도+대기 CO2 농 도), ‘High CO2+High Temp.’(상승온도+상승 CO2
농도)이었다. 상승온도 처리구들은 대기온도보다 4oC 상 승시켰고, 상승 CO2 농도 처리구들은 700µmol·mol−1로
유지하였다. 4년 동안 매년 처리기간은 4월말부터 11 월초까지였다. CO2 상승은 기공전도도와 잎의 엽록체 함량(SPAD 계량기 값)을 감소시켰으나, 광합성속도, 세포 내 CO2농도(Ci) 및 잎의 전분함량은 증가시켰 다. 수체생장에 있어, 기온 상승은 나무당 총 신초수 와 총 신초생장량을 증가시켰으나, CO2상승은 평균 신초장을 감소시켰다. 과실품질에 있어, CO2 상승은 착색, 가용성 고형물 함량, 및 에틸렌 발생량을 증진 시켰다. 결론적으로, CO2농도가 상승되면 생육초기에 사과나무의 광합성속도가 증가되었으나 생육후기에는 CO2 상승에 따른 광합성속도 증진 효과가 감소되었다.
반면에 기온 상승은 생육초기 광합성속도를 감소시켰 으나 생육후기에 광합성속도를 증진시키는 경향이 있 었다. CO2와 기온의 동시 상승은 각 요인에 의한 광 합성 감소 정도가 줄어드는 경향이 있었다.
REFERENCES
Byun, J. Y., S. S. Lee, K. S. Choi, and S. M., Kang, 2006: Plant Physiology (2nd Ed.). Byun, J. Y. (Eds.) Hyangmoonsha Press. Seoul, 52-64, 145-153, and 439-440. (In Korean) Chae, J. C., S. J. Park, B. H. Kang, and S. H. Kim, 2006:
Principles of Crop Cultivation (3rd ed.). Chae, J. C.
(Ed.). Hyangmoonsha Press, Seoul, 190-195. (In Korean) Chung, U., K. J. Lee, and B. W. Lee, 2007: Preliminary
report of observed urban-rural gradient of carbon dioxide concentration across Seoul, Suwon, and Icheon in South Korea. Korea Journal of Agricultural and Forest Meteorology 9(4), 268-276. (In Korean with English abstract)
Chung, M. S., S. H. Han, D. H. Kim, J. C. Lee, and P. G.
Kim, 2012: Effects of elevated atmospheric CO2 and nitrogen fertilization on growth and carbon uptake of yellow poplar seedlings. Korea Journal of Agricultural and Forest Meteorology 14(3), 108-118. (In Korean with English abstract)
Forshey, C. G and D. A. Elfving, 1989: The relationship between vegetative growth and fruiting in apples. Horticultural Review 11, 229-287.
Fujii, J. A. and R. A. Kennedy, 1985: Seasonal changes in the photosynthetic rate in apple tree: a comparison between fruiting and nonfruiting tree. Plant Physiology 78(3), 519-524.
Han, J. H., J. G. Cho, I. C. Son, S. H. Kim, I. B. Lee, I. M.
Choi, and D. Kim, 2012: Effects of elevated carbon dioxide and temperature on photosynthesis and fruit characteristics of ‘Niitaka’ pear (Pyrus pyrifolia Nikai).
Horticulture, Environment, and Biotechnology 53(5), 357-361.