INTRODUCTION
Phosphorous (P) is the most limiting plant nutrient, as greater part of the soil phosphorous, approximately 95~99% is present in the form of insoluble phosphates (Vassileva et al. 2001). Large proportion of phosphate in P-deficient soil and available P in natural soil are easily converted into in-soluble complexes such as iron and aluminum hydrous oxi-des, crystalline and amorphous aluminum silicates and cal-cium carbonates (Altomare et al. 1999). The release of fix-ed and poorly soluble form of P is an important aspect for increasing soil fertility. Being a part of rich rhizosphere
mi-crobial diversity, P-solubilizing bacteria play an important role in dissolution of insoluble phosphate from soil and allowing sustainable use of phosphate fertilizer. Various mechanisms like lowering pH by acid production, ion che-lation and exchange reactions in the growth environment have been reported to play a key role in P-solubilization by bacteria. Gram negative bacterial isolates are considered as good phosphate solubilizers, probably because of the involve-ment of direct oxidation of glucose into gluconic acid (Tripu-ra et al. 2007). Low molecular weight organic acids, main-ly gluconic acid (GA) and ketogluconic acids (KGA) are produced in the periplasm of Gram negative bacteria by direct oxidation pathway of glucose (DOPG) enzymes i.e., glucose dehydrogenase (GDH) and gluconate dehydrogenase (GADH) (Deubel et al. 2000; Goldstein 2000). Tripura et
─ ─ 39 ──
Mineral Phosphate Solubilization by Wild Type and Radiation
Induced Mutants of Pantoea dispersa and Pantoea terrae
Senthilkumar Murugesan*, Young-Keun Lee and Jung Hun Kim Radiation Application Research Division, Korea Atomic Energy Research Institute,
Jeongeup 580-185, Korea
Abstract -- Three mineral phosphate solubilizing (MPS) bacteria where isolated from rhizosphere soil samples of common bean and weed plants. 16S rDNA analysis indicated that the isolate P2 and P3 are closely related to Pantoea dispersa while isolate P4 is closely related to Pantoea terrae. Isolates P2 and P3 recorded 381.60μμg ml--1and 356.27μμg ml--1of tricalcium phosphate (TCP) solubilization respectively on 3 days incubation. Isolate P4 recorded the TCP solubilization of 215.85μμg ml--1and the pH was dropped to 4.44 on 24 h incubation. Further incubation of P4 sharp-ly decreased the available phosphorous to 28.94μμg ml--1and pH level was raised to 6.32. Gamma radiation induced mutagenesis was carried out at LD99dose of the wild type strains. The total of 14 mutant clones with enhanced MPS activity and 4 clones with decreased activity were selected based on solubilization index (SI) and phosphate solubilization assay. Mutant P2-M1 recorded the highest P-solubilizing potential among any other wild or mutant clones by releasing 504.21μμg ml--1 of phosphorous i.e. 35% higher than its wild type by the end of day 5. A comparative evaluation of TCP solubilization by wild type isolates of Pantoea and their mutants, led to select three MPS mutant clones such as P2-M1, P3-M2 and P3-M4 with a potential to release ¤¤471.67μμg ml--1of phosphorous from TCP. These over expressing mutant clones are considered as suitable candidates for biofertilization.
Key words : Gamma irradiation, Mineral phosphate solubilization, Pantoea
* Corresponding authors: Senthilkumar Murugesan, Tel. +82-63-570-3304, Fax. +82-63-570-3309, E-mail. [email protected]
negative isolates and also developed mutants with enhanced MPS activity compared with wild type Serratia marcescens GPS-5. The present work was aimed to compare the MPS activity of Pantoea strains and to develop the mutants with enhanced MPS activity through radiation mutagenesis.
MATERIALS AND METHODS
1. Isolation and identification of P-solubilizing bacteria
Soil samples were collected from bean rhizosphere and dry fields in Ipyeong-ri, Jeollanam-do province, South Korea. Samples were serially diluted and plated on Pikovskaya (PVK) agar with 0.5% tricalcium phosphate (Pikovskaya 1948). Mineral phosphate solubilizing colonies with clear halo zone were purified and stored at -70�C as 25% gly-cerol stocks. The clones were identified based on 16S rDNA sequence. Genomic DNA was extracted by using Qiagen DNeasy Plant Mini Kit (Qiagen, Valencia, CA). 16S rDNA was amplified by using the primers 27F (5 ′-AGAGTTTGA-TCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTA-CGACTT-3′) (Lane 1991). The sequences were determined by fluorescent dye terminators method using ABI prismTM BigdyeTM terminator cycle sequencing ready reaction kit V.3.1. Products were run on ABI 3730XL capillary DNA sequencer (ABI prism 310 Genetic analyzer, Tokyo, Japan). Sequences were subjected to BLASTn analysis with the NCBI data base and phylogenetic trees were generated with NJPLOT programme of CLUSTAL X (Thompson et al. 1997).
2. Phosphate solubilization assay
Solubilized phosphorous by selected bacterial isolates was quantitatively estimated in PVK broth containing 0.5% tricalcium phosphate. One hundred ml of PVK broth in 500 ml flask was inoculated with 1ml of overnight cultured bac-terial isolates (~107~108CFU ml-1) and incubated at 28± 2�C, 200 rpm for 5 days. Uninoculated medium incubated along with other treatments served as control. Each 10 ml of culture supernatants was taken at different time intervals by centrifugation at 5,000 rpm, 4�C for 15 min and used to determine the available phosphorous as well as pH.
Phos-ascorbic acid method (Watanabe and Olsen 1965) and ex-pressed as μg ml-1over the control. One ml of culture broth taken at different time intervals was used to determine the bacterial population by standard plant count method.
3. Radiation sensitivity
Cell pellets were collected by centrifuging 20 ml of log phase bacterial isolates cultured in LB, broth washed twice with sterile distilled water and suspended (~107~109CFU ml-1). 500μl of cell suspensions were transferred to 1.5 ml microcentrifuge tubes and irradiated at 0.25, 0.5, 0.75, 1, 3, 5 kGy respectively. Samples were irradiated in cobalt-60 irradiator at Advanced Radiation Technology Institute, Ko-rea Atomic Energy Research Institute, South KoKo-rea. Bacte-rial population in irradiated samples was determined by se-rial dilution and plate count method. The D10-value was de-termined by plotting radiation dose (kGy) on X-axis and survival ratio on Y-axis. Negative reciprocal of slope indi-cated the D10-value.
4. Radiation mutagenesis and selection
To induce the mutagenesis, bacterial cell suspensions were prepared as described above and exposed to gamma irradiation at the dose of LD99. Irradiated samples were seri-ally diluted and plated on PVK agar and incubated at 28± 2�C for 4 days. Master plates were prepared from the mutant clones and stored under refrigerated conditions. MPS acti-vity of mutant clones was determined by spot inoculating them on PVK agar as well as National Botanical Research Institute’s phosphate growth medium containing bromophe-nol blue (NBRIP-BPB) agar media. P-solubilizing potentials of clones were initially determined by their P-solubilization index and further quantitatively determined by the ascorbic acid method (Watanabe and Olsen 1965) and expressed as μg ml-1over the control.
RESULTS
1. Isolation and identification of phosphorous solubilizing rhizobacteria
Plating of serially diluted soil samples on PVK agar pro-duced several P-solubilizing colonies with varying diameter
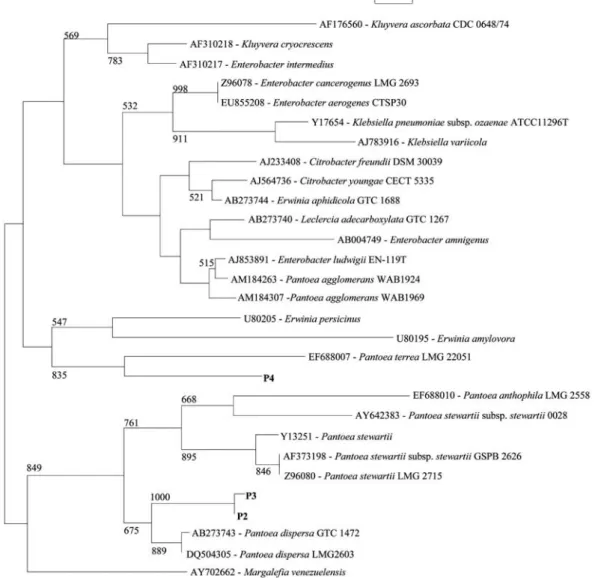
Fig. 1. 16S rDNA sequence based phylogenetic tree showing the relationship of phosphate solubilizing isolates with other related genera. Bootstrap analysis was done with 1,000 cycles. Only bootstrap values ¤50% are given at the nodes.
Table 1. Phosphorous solubilizing activity of Pantoea wild strains
Bacterial isolate Incubation duration(days) P-solubilization(μg ml-1) pH Population(cfu ml-1)
1 251.03±7.05c 4.25 12.83±2.86×106
3 381.60±4.48g 4.14 6.633±1.3×103
Pantoea dispersa P2 5 364.09±4.32efg 4.16
-7 362.02±7.91defg 4.16 -9 367.67±14.22fg 4.18 -1 238.60±14.84bc 4.23 11.00±1.01×106 3 356.27±5.94defg 4.17 5.83±1.19×106 Pantoea dispersa P3 5 335.01±6.82d 4.20 -7 347.83±5.62def 4.21 -9 339.14±10.87de 4.20 -1 215.85±5.89b 4.44 6.7±2.57×109 3 28.94±2.50a 6.32 24.3±3.68×106 Pantoea terrae P4 5 8.49±0.67a 6.46 15.2±1.36×106 7 10.43±1.24a 6.26 1.69±0.13×106 9 9.28±0.42a 6.13 4.26±2.17×105
zones greater than 10 mm were selected and purified. P2 and P3 were isolated from field soil, while P4 was isolated from bean rhizosphere.
16S rDNA analysis indicated that Pantoea dispersa was the closest related species to P2 and P3 with 99.0% and 97.4% sequence similarity, respectively. Isolate P4 showed 98.8% sequence similarity with Pantoea terrae. Bacterial isolates generally differed in their mineral phosphate solu-bilizing abilities. Isolate P2 recorded larger zone of clearance i.e. 14 mm in diameter compared to that of other isolate P3 and P4 that produced 12 and 10 mm clearing zone respec-tively. Growth kinetics, pH reduction and P-solubilization of these isolates were studied in PVK broth for 9 days. Iso-lates P2 and P3 recorded 251.03 and 238.60μg ml-1of TCP solubilization respectively on 24 h incubation (Table 1). Fur-ther incubation improved the TCP solubilization and record-ed maximum value by P2 (381.60μg ml-1) and P3 (356.27
μg ml-1) on 3 days incubation. However, population levels of P2 and P3 on 5thday were reduced to zero due to the low
pH of the environment. Further incubation upto 9 days did not influence the available phosphorous. Isolate P4 recorded the TCP solubilization of 215.85μg ml-1on 24 h incubation and the pH was dropped to 4.44. However, the available P-content was sharply reduced to 28.94μg ml-1 on the 3rdday
of incubation and pH value was raised to 6.32. In contrast to P2 and P3, P4 population was not reduced to zero on the 5thday of incubation and recorded 107CFU ml-1.
2. Development of MPS mutants by gamma irradiation
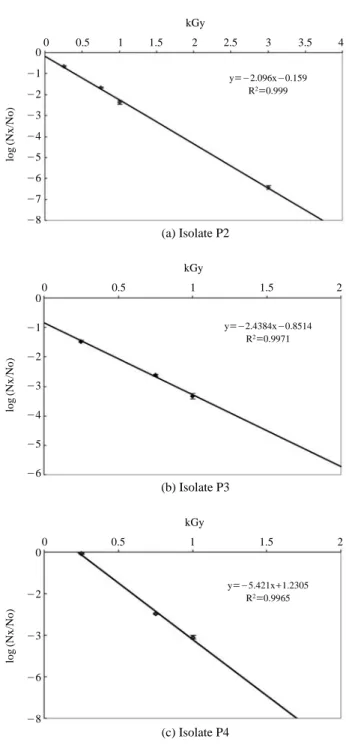
Radiation sensitivity of phosphate solubilizing isolates was studied in order to determine D10and LD99values. D10
value-radiation dose that produce 90% reduction in viable bacterial cells was determined by taking the negative reci-procal of slope (-1/slope) (Fig. 2). P2 and P3 recorded the
higher D10-value (0.48 kGy and 0.41 kGy respectively) than
isolate P4. P4 is more radiation sensitive and recorded the D10-value of 0.18 kGy. LD99value-radiation dose required
to kill 99% of viable bacterial cells was determined by using the formula LD99==-log (0.01)/slope. Isolates P2, P3 and P4
recorded the LD99 values of 0.95 kGy, 0.82 kGy and 0.37
kGy respectively.
The isolates were exposed to gamma ray for their LD99
values and plated on Pikovskaya agar. The mutant clones with altered MPS activity were transferred to master plates and selected based on their solubilization index (SI). Five mutants with enhanced MPS activity were developed from wild type P4. Among them, P4-M5 released 336.84μg ml-1
0 -1 -2 -3 -4 -5 -6 -7 -8 0 -1 -2 -3 -4 -5 -6 0 -2 -3 -6 -8 kGy 0 0.5 1 1.5 2 kGy 0 0.5 1 1.5 2 (a) Isolate P2 (b) Isolate P3 (c) Isolate P4 y==-2.096x-0.159 R2==0.999 y==-2.4384x-0.8514 R2==0.9971 y==-5.421x++1.2305 R2==0.9965 log (Nx/No) log (Nx/No) log (Nx/No)
Fig. 2. Radiation sensitivity of mineral phosphate solubilizing
bac-terial isolate P2 (a), isolate P3 (b) and isolate P4 (c). 500μl of bacterial cell suspensions (~107~108CFU ml-1) were
exposed to gamma rays and reduction on their population was determined. Negative reciprocal of slope indicated the D10-value.
phosphorous i.e. 52.4% higher than that of wild type P4 by the end of day 1. However, further incubation drastically reduced the available P ranging from 95.02 to 8.2μg ml-1 with increased pH of spent medium. Mutant P2-M1 record-ed the highest P-solubilizing potential among any other wild or mutant clones by releasing 504.21μg ml-1P i.e. 35% higher values than its wild type by the end of day 5. Another mutant, P2-M2 recorded the stable increase in MPS activity with 45.5% and 24.97% increased release of P over its wild type by the end of day1 and day 5 respectively. In the case of P3 mutants, P3-M2 released 487.3μg ml-1P with 34.4% increase over wild type strain.
DISCUSSION
Phosphorous found in the soil as apatite, hydroxyapatite,
and oxyapatite as well as found associated with the surface of hydrated oxides of Fe, Al and Mn are poorly soluble and assimilative. Mineral phosphate solubilizing microorgan-isms can significantly affect the phosphorous cycling in both natural and agricultural ecosystem. Recently, phosphate solubilizing potential of Pantoea species was well estab-lished. Pantoea ananatis and Pantoea agglomerans strains isolated from iron rich soil samples, showed their potential on mineral phosphate solubilization (Perez et al. 2007). Sel-vakumar et al. (2008) isolated a cold tolerant Pantoea dis-persa 1A that solubilized 17.9μg of P2O5ml-1day-1at 15�
C in NBRIP broth. In this study, we isolated potential phos-phate solubilizing strains of Pantoea dispersa and Pantoea terrae. All these isolates reduced the pH of PVK broth ap-proximately 3 units from 7 to 4 on 24 h incubation. Produc-tion of low molecular weight organic acid and their ability
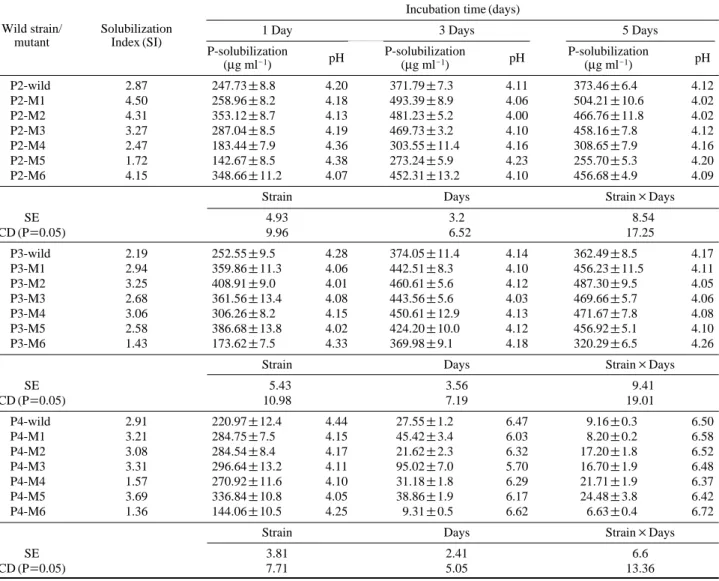
Table 2. Phosphorous solubilizing activity of wild strains and their mutants
Incubation time (days)
1 Day 3 Days 5 Days
Wild strain/ Solubilization
P-solubilization P-solubilization P-solubilization mutant Index (SI)
(μg ml-1) pH (μg ml-1) pH (μg ml-1) pH P2-wild 2.87 247.73±8.8 4.20 371.79±7.3 4.11 373.46±6.4 4.12 P2-M1 4.50 258.96±8.2 4.18 493.39±8.9 4.06 504.21±10.6 4.02 P2-M2 4.31 353.12±8.7 4.13 481.23±5.2 4.00 466.76±11.8 4.02 P2-M3 3.27 287.04±8.5 4.19 469.73±3.2 4.10 458.16±7.8 4.12 P2-M4 2.47 183.44±7.9 4.36 303.55±11.4 4.16 308.65±7.9 4.16 P2-M5 1.72 142.67±8.5 4.38 273.24±5.9 4.23 255.70±5.3 4.20 P2-M6 4.15 348.66±11.2 4.07 452.31±13.2 4.10 456.68±4.9 4.09
Strain Days Strain×Days
SE 4.93 3.2 8.54 CD (P==0.05) 9.96 6.52 17.25 P3-wild 2.19 252.55±9.5 4.28 374.05±11.4 4.14 362.49±8.5 4.17 P3-M1 2.94 359.86±11.3 4.06 442.51±8.3 4.10 456.23±11.5 4.11 P3-M2 3.25 408.91±9.0 4.01 460.61±5.6 4.12 487.30±9.5 4.05 P3-M3 2.68 361.56±13.4 4.08 443.56±5.6 4.03 469.66±5.7 4.06 P3-M4 3.06 306.26±8.2 4.15 450.61±12.9 4.13 471.67±7.8 4.08 P3-M5 2.58 386.68±13.8 4.02 424.20±10.0 4.12 456.92±5.1 4.10 P3-M6 1.43 173.62±7.5 4.33 369.98±9.1 4.18 320.29±6.5 4.26
Strain Days Strain×Days
SE 5.43 3.56 9.41 CD (P==0.05) 10.98 7.19 19.01 P4-wild 2.91 220.97±12.4 4.44 27.55±1.2 6.47 9.16±0.3 6.50 P4-M1 3.21 284.75±7.5 4.15 45.42±3.4 6.03 8.20±0.2 6.58 P4-M2 3.08 284.54±8.4 4.17 21.62±2.3 6.32 17.20±1.8 6.52 P4-M3 3.31 296.64±13.2 4.11 95.02±7.0 5.70 16.70±1.9 6.48 P4-M4 1.57 270.92±11.6 4.10 31.18±1.8 6.29 21.71±1.9 6.37 P4-M5 3.69 336.84±10.8 4.05 38.86±1.9 6.17 24.48±3.8 6.42 P4-M6 1.36 144.06±10.5 4.25 9.31±0.5 6.62 6.63±0.4 6.72
Strain Days Strain×Days
SE 3.81 2.41 6.6
to form stable metal complexes were linked with MPS sol-ubilization (Goldstein 1986; Salih et al. 1989; Mattey 1992). Inorganic P-solubilization by Pantoea agglomerans was also linked to a mechanism involving both gluconic acid production and proton extrusion as a consequence of NH4++
uptake (Sulbaran et al. 2009). In contrast to isolate P2 and P3, a decrease in the available phosphorous content and increase in the pH (upto pH 6.46) of growth medium was reported towards the end of incubation by isolate P4. Pre-sence of soluble phosphate may have an inhibitory effect on further phosphate solubilization (Narsian et al. 1995) by showing the negative effect on microbial acid productivity (Gyaneshwar et al. 1999; Krishnaraj et al. 2001). Increase
organic acids as part of their own metabolism. In contrast to this, pH of isolate P4 was not increased in PVK broth where tricalcium phosphate was replaced with K2HPO4and the
population was reduced to zero on 5 days after inoculation (Data not shown). Calcium ions released from TCP may mediate the uptake of gluconic acid by P4 isolate.
We also developed the mutant clones with altered MPS activity through gamma irradiation induced mutagenesis. Significant increase in soluble P level and decrease in the pH of culture supernatant of mutants over wild type strain was observed and the relationship could be established bet-ween amount of P solubilized and drop in pH by all mutants and wild type strains including isolate P4. Achal et al. (2007) reported 35.04% increased P solubilization by UV induced mutant of Aspergillus tubingensis AtM-5 (47.4 mg 100 ml-1) over wild type At-W (35.1 mg 100 ml-1). Tripura et al. (2007) reported that EMS XVIII Sm-35, a mutant developed throu-gh ethyl methane sulfonate mutagenesis, showed 40% in-crease in amount of phosphate released in liquid medium compared with wild type Serratia marcescens GPS-5. In the present study, mutant P2-M1 recorded the highest P-solubilizing potential among any other wild or mutant clones by releasing 504.21μg ml-1P i.e. 35% higher values than its wild type by the end of day 5, therefore it could be used as biofertilizing agents.
ACKNOWLEDGMENT
This study was carried out under the Nuclear R & D Pro-gram of the Ministry of Education, Science and Technology, Republic of Korea.
REFERENCES
Achal V, Savant VV and Reddy MS. 2007. Phosphate solubili-zation by a wild type strain and UV-induced mutants of
Aspergillus tubingensis. Soil Biol. Biochem. 39:695-699.
Altomare C, Norvell WA, Bjorkman T and Harman GE. 1999. Solubilization of phosphates and micronutrients by plant growth promoting and biocontrol fungus Trichoderma
har-zianum Fifai 1295-22. Appl. Environ. Microbiol.
65:2926-2933.
Deubel A, Gransee A and Merbach W. 2000. Transformation
Fig. 3. Mineral phosphate solubilizing ability of wild type strains
and their mutants on PVK agar plates. Wild and mutant clones were spot inoculated on PVK agar plates and observ-ed 4 days after incubation.
of organic rhizo-depositions by rhizosphere bacteria and its influence on the availability of tertiary calcium phosphate.
J. Plant Nutr. Soil Sci. 163:387-392.
Goldstein AH. 1986. Bacterial solubilization of mineral phos-phates: Historic perspective and future prospects. Am. J.
Altern. Agric. 1:57-65.
Goldstein AH. 2000. Bioprocessing of rock phosphate ore: essen-tial technical considerations for the development of a suc-cessful commercial technology. pp. 1-21. Alfred Univer-sity, New York.
Gyaneshwar P, Nareshkumar G and Parekh LJ. 1998. Effect of buffering on phosphate solubilizing ability of microorga-nisms. World J. Microbiol. Biotechnol. 5:72-74.
Krishnaraj PU and Goldstein AH. 2001. Cloning of a Serratia
marcescens DNA fragment that induces quinoprotein
glu-cose dehydrogenase mediated gluconic acid production in
Escherichia coli in the presence of a stationary phase Ser-ratia marcescens. FEMS Microbiol. Lett. 205:215-220.
Lane DJ. 1991. 16S/23S rRNA sequencing. pp. 115-175. In: Nucleic Acid Techniques in Bacterial Systematics (Stacke-brandt E and Goodfellow M eds.) Chichester: Wiley. Mattey M. 1992. The production of organic acids. Rev.
Biotech-nol. 12:87-113.
Perez E, Sulbaran M, Ball MM and Yarzabal LA. 2007. Isola-tion and characterizaIsola-tion of mineral phosphate solubilizing bacteria naturally colonizing a limonitic crust in the south-eastern Venezuelan region. Soil Biol. Biochem. 39:2905-2914.
Pikovskaya RI. 1948. Mobilization of phosphorus in soil con-nection with the vital activity of some microbial species.
Microbiologiya. 17:362-370.
Salih HM, Yahya AI, Abdul-Rahem AM and Munam BH. 1989.
Availability of phosphorous in calcareous soil treated with rock phosphate or super phosphate as affected by phosphate dissolving fungi. Plant Soil. 120:181-185.
Selvakumar G, Kundu S, Joshi P, Nazim S, Gupta AD, Mishra PK and Gupta HS. 2008. Characterization of a cold tolerant plant growth promoting bacterium Pantoea dispersa 1A isolated from a sub-alpine soil in the north western Indian Himalayas. World J. Microbiol. Biotechnol. 24:955-960. Sulbaran M, Perez E, Ball MM, Bahsas A and Yarzabal LA.
2008. Characterization of the mineral phosphate solubiliz-ing activity of Pantoea agglomerans MMB051 isolated from an iron rich soil in southeastern Venezuela (Bolivar State). Curr. Microbiol. 58:378-383.
Thompson JD, Gibson TJ, PleWniak F, Jeanmougin F and Hig-gins DG. 1997. The clustalx windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 24:4876-4882.
Tripura C, Sashidhar B and Podile AR. 2007. Ethyl methane sulfonate mutagenesis-enhanced mineral phosphate solubi-lization by groundnut associated Serratia marcescens GPS-5. Curr. Microbiol. 54:79-84.
Vassileva M, Vassilev N, Fenice M and Federici F. 2001. Im-mobilized cell technology applied in solubilization of insol-uble inorganic (rock) phosphate and P plant acquisition.
Bioresource Technol. 79:263-271.
Watanabe FS and Olsen SR. 1965. Test of an ascorbic acid method for determining phosphorus in water and NaHCO3
extracts from soil. Soil Science Society of America
Proceed-ings 29:677-678.
Manuscript Received: March 2, 2009 Revision Accepted: March 20, 2009