INTRODUCTION
The 5-methyltryptophan (5MT) is a tryptophan analog and can fit into the allosteric site of anthranilate synthase in the same way as tryptophan or by some other mechanisms. Some papers reported that 5MT resistant mutants accu-mulated tryptophan in rice, tobacco (Wakasa, 1985) and Arabidopsis plant (Li and Last., 1996). The cell responds to the 5MT by halting further synthesis of tryptophan. As the
5MT is unable to replace tryptophan for a protein synthesis and then plant growth is finally ceased. However, the mechanism of an inhibition is not important as long as the mutants that oversynthesize the tyrptophan will be resistant to growth inhibition. The resistance mechanism of amino acid analog is reported to be the altered feedback inhibition (Widholm 1977). The core feedback related enzyme was inhibited by the end-product amino acid. Anthranilate synthase enzyme, which converts chorismate and an amino donor (usually glutamine) to anthranilate, appears to be a key regulated enzyme. In a previous report, we isolated and characterized Oryza sativa Anthranilate Synthase Alpha 2 (OASA2, accession no. CD670874) mutant gene, which is
─ ─ 123 ──
Characterization of Differentially Expressed Genes Related to
5-Methyltryptophan Resistance in Rice (Oryza sativa L.)
Jin-Baek Kim, Dong Sub Kim*, Cheol Seong Jang1, Geung-Joo Lee, Si-Yong Kang, Hye Jung Lee, Jae Young Song and Yong Weon Seo1 Radiation Research Center for Bio-technology , Advanced Radiation Research Institute, Korea Atomic Energy Research Institute, Sinjeong 1266, Jeongeup, Jeonbuk 580-185, Korea
1Division of Biotechnology and Genetic Engineering, College of Life & Environmental Sciences,
Korea University, Anam, Seongbuk, Seoul 136-701, Korea
Abstract -- In order to identify differentially expressed genes between wild-type and
5-Methy-ltryptophan (5MT) resistant rice mutant under growth inhibition by 5MT, we constructed cDNA library from leaves and roots of 5MT resistant rice mutant. The 5MT-resistant M4mutants
deriv-ed from the mutant cell lines resistant to 5MT were obtainderiv-ed by in vitro mutagenesis using gam-ma-rays, and identified differentially expressed genes to them using differential hybridization method. Twenty-six clones were represented as they might have differentially expressed genes from cDNA library of 5MT resistant mutants in random selected 223 clones by single pass sequ-encing and BLAST algorithm program. Five genes encoding heat shock protein 82 (OsHSP1), putative chorismate mutase/ prephenate dehydratase (OsCM1), enolase (2-phosphoglycerate de-hydratase) (2-phospho-glycerate hydrolyase) (OsEno1), polyubiquitin (OsPU1), and putative hyd-roxyproline-rich glycoprotein (OsHRP1), were analyzed by Northern blotting to identify an expre-ssion level at various abiotic stress conditions. Temporal patterns of these genes in response to two tryptophan analogs (5MT and αα-MT) and abiotic stresses (ABA and NaCl) are similar between 5MT resistant mutants and the wild-type control plants. However, transcript levels of these genes were higher in 5MT resistant mutants than in control plants.
Key words : Differential hybridization, Mutant, Northern blot, Rice, 5-Methyltryptophan
Equally contributed first authors: Jin-Baek Kim and Dong Sub Kim * Corresponding author: Dong Sub Kim, Tel. +82-63-570-3311,
one of the key regulated enzyme subunits in the tryptophan biosynthesis pathway, under two tryptophan analogs (5MT and αMT) inhibition and other abiotic stresses (ABA, NaCl, and cold) (Kim et al. 2005b). The OASA2 gene in the 5MT resistant mutant lines was highly expressed in a low 5MT concentration and at an early stage of the 5MT and α MT treatments. However, the mRNA accumulation of the putative OASA2 gene in the mutant plants was gradually decreased by abiotic stresses such as NaCl and a cold-treatment. These results indicate that the 5MT resistance in the mutant lines was due to the altered anthranilate synthase forms. Moreover, in another study, we also suggested that amino acid analog creates stress in vivo much like other environmental stresses, which strongly affects the biomass production (Kim et al. 2005a). Many of these adverse environmental stress conditions such as drought, excess light and extreme temperatures, have been claimed to produce at least part of their deleterious effect through an increased accumulation of ROS, resulting in protein and DNA damage and lipid peroxidation. 5MT resistance me-chanism except for alterations of enzymes in vivo has not been reported, yet.
The discovery of novel genes, determination of their expression patterns in response to abiotic stress, and an improved understanding of their roles in a stress adaptation will provide the basis of effective engineering strategies leading to a greater stress tolerance (Cushman and Bohnert 2000). The progress of genomic studies is now anticipated through the use of techniques such as a high-throughout analysis of expressed sequence tags (ESTs), large-scale parallel analysis of a gene expression, targeted or random mutagenesis, and a gain-of-function or mutant complemen-tation (Zhang et al. 2001). Large-scale single-pass sequ-encing of randomly selected cDNA clones, referred to EST, will provide a cost-effective and rapid alternative route toward the isolation of amino acid analog resistance-related genes and the discovery of broad additional data (Stanton 2001). The differential hybridization method has been used successfully to identify differentially expressed genes from many experimental systems. It has advantages in that is relatively less time-consuming and can result in the iden-tification of rare cDNA, which may be missed by a conven-tional cDNA library screening. Therefore, this method is one of the most powerful techniques in the detection of genes preferentially expressed in different cells and tissues
(Yu and Goh 2000; Takemoto et al. 2001; Ueda et al. 2002). We have been modified the method to improve its sensitivity. In general, traditional approaches to the analysis of mRNA abundance, such as a Northern blotting, tend to be limited by the number of transcripts that can be simul-taneously analyzed. More recent innovations, such as a hybridization to arrayed cDNA libraries or oligonucleotide chips permit a simultaneous analysis of the abundance of thousands of transcripts (Brown and Botstein 1999). In plants, the use of arrays of partially sequenced cDNAs has been successfully applied to the analysis of a gene expre-ssion in light- and dark-grown seedlings of Arabidopsis (Desprez et al. 1998).
In this study, a modified differential screening method was used in order to detect differentially expressed genes between the control and 5MT resistant mutants. This me-thod involves the preparation of several membranes con-taining an equal amount of target DNA from cDNA library and hybridization with different probes which are differen-tially expressed dig-labeled mRNA, and provides a high sensitivity for quantification change and allows for a simultaneous detection of differentially expressed genes with few false positives. Transcript levels of five differen-tially expressed genes, OsPU1, OsHRP1, OsHSP1, OsEno1, and OsCM1, were also investigated under two tryptophan analogs (5MT and αMT) and two abiotic stresses (ABA and NaCl) by a RNA gel blot analysis.
MATERIALS AND METHODS
Plant materialsThe mutant cell lines resistant to growth inhibition by 5MT were selected from the callus irradiated with gamma ray of 50 Gy through an embryo culture. Regenerated plants from 5MT resistant calli were selfied and the M3progeny were obtained. Complementary DNA were constructed from leaves and roots of the 5MT resistant M3plants.
RNA isolation and cDNA construction
M3seeds were germinated and grown in a nutrient solu-tion with 0.5 mM 5MT. After 4 weeks, leaves and roots from the 5MT resistant plants were harvested and imme-diately frozen in liquid nitrogen and stored in -70�C until used.
Total RNA from the bulked leaf and root tissues were extracted using Trizol reagent Poly (A)++
mRNA and sepa-rated from total RNA using PolyATract mRNA isolation system. A cDNA library was constructed from 5µg of Poly (A)++
mRNA using ZAP-cDNA Gigapack III cloning kit (Stratagene) and was cloned into pBluescript II SK (++) (Stratagene). The packaged cDNA (primary library) was amplified by Host bacteria (SOLR strain).
Differential hybridization
Synthesis of a labelled probe and a differential hybridiza-tion were performed with a digoxigenin (DIG) labelling and detection system of alkali phosphatase and chemilumine-scent substrate CSPD (Boehringer Mannheim).
Plasmid DNA of 200 ng was denatured with an alkali solution (0.4 M NaOH and 10 mM EDTA). The denatured DNA was blotted on a positive nylon membrane using a vacuum dot-blot apparatus. Dig labelled probes were syn-thesized from mRNAs of 5MT non-treated Donganbyeo (control), 5MT treated resistant plants, and 5MT treated sensitive plants. The membrane blotted with target DNA was treated with 10 ml of standard buffer (750 mM NaCl, 75 mM sodium citrate; pH 7.0, 0.1% sodium lauroylsar-cosine, 0.02% SDS, and 1% blocking reagent) at 68�C for 2 hours and hybridized with 10 ml of labeled DNA (5~25
ng/ml) in standard buffer at 68�C for 12~16 hours.
Hybri-dized membrane was rinsed twice with a washing solution I (0.3 M NaCl 0.15 M sodium citrate; pH 7.0, and 0.1% SDS) for 5 minutes per each wash and three times with a washing solution II (15 mM NaCl, 7.5 mM sodium citrate; pH 7.0, and 0.1% SDS) for 15 minutes, 10 minutes, and 10 min-utes. After the membrane was washed with a maleic acid buffer (150 mM NaCl, 100 mM maleic acid; pH 7.5) for 1 minute, it was blocked with 1% blocking reagent for 45 minutes, and incubated with anti-DIG-AP (Boehrin-ger Mannheim) diluted 1 : 10,000 (v/v) for 30 minutes. Unbo-und antibodies were removed by washing buffer III [maleic acid buffer plus 0.3% (v/v) Tween 20] for three times. The membrane was equilibrated with a detection buffer (100 mM Tris, 100 mM NaCl, 50 mM MgCl2; pH 9.5) for 2 minutes and incubated with CSPD solution diluted 1 : 100 in a detection buffer for 5 minutes. The membrane was covered with plastic wrap and placed under three layers of X-ray films in an exposure cassette followed by an
incu-bation at 37�C for 30 minutes.
Sequencing of the differential displayed clones Insert DNAs from differentially displayed clones were amplified using T3 promoter primer (Promega) with the BigDye Terminator Cycle Sequencing Ready Reaction Kit (Perkin Elmer) for sequencing. Sequenced DNA bases were compared with the BLAST algorithm of the National Cen-ter for Biotechnology Information (NCBI).
Northern blot analysis
5MT resistant mutants and control plants were grown for 4 weeks at 25/18�C (day/night) in a greenhouse. For the two tryptophan analogs (5MT and α-Methyltryptophan (α-MT)) and abiotic stresses (abscisic acid (ABA) and NaCl) treat-ment, 4-week-old plants were transferred to a growth cham-ber at 25/18�C (day/night) for 24 h on a 16/8-h (day/night) photoperiod. The treated concentration of 5MT, α-MT, ABA, and NaCl were 500µM, 200µM, 10µM, and 200 mM, respectively. Plants were harvested at 3, 6, 12, 24, and 48 h after an initiation of the treatments.
Total RNA was isolated from individual tissues using Trizol acording to commercial protocols (GibcoBRL, Gai-thersburg, Md.). Total RNAs (20µg) were fractionated on a 1% formaldehyde agarose gel and transferred to a nylon membrane (MSI). cDNAs to be used as probes were labell-ed using the Random Primlabell-ed DNA Labeling kit (Boehrin-ger Mannheim). The blots were prehybridized for 1 h and then hybridized for 18 h at 65�C in a hybridization solution [1 mM EDTA, 7% SDS, 0.25 M disodium phosphate (pH 7.2), and 5% dextran sulfate]. The membrane was washed with a washing buffer followed by an incubation with Avidx-AP conjugation, and then incubated with a CDP-star solution (Tropis). The membrane was covered with X-ray film, and exposed for 20 minutes at room temperature.
RESULTS AND DISCUSSION
Characteristics of the constructed cDNA library Rice cDNA library was constructed from high-amino acid accumulating M3mutant plants, which were induced from gamma-irradiation mutagenized embryo cultures. Therecombinant efficiency analyzed by white/blue selection was more than a 90% for a primary library. Titer analysis of the primary library indicated that greater than 5×106 plaque forming unit (pfu) were produced by packaging of the vector/cDNA ligation mixture. Phage amplification of the primary library yielded approximately 120 ml of phage -containing SM buffer with a titer of about 3.55×1010 pfu/ ml. The inserted DNA sizes were distributed from 0.35~
2.5 kbp with an average value of 1.14 kbp. Since the 5′ un-translated region (UTRs) of many plant genes are relati-vely short (Joshi 1987), this cDNA library was used to ob-tain sequences from 5′end of the transcripts. Therefore, the cDNA clone derived from this library could provide a high-er chance of sequencing coding regions, which is vhigh-ery help-ful for an identification of the ESTs.
Generating ESTs from 5MT resistant mutant cDNA library will contribute to a gaining a better understanding for amino acid biosynthesis and amino acid analog resistant mechanism. In addition, the ESTs provide necessary mole-cular clones that can be used to prove directly into the phy-siology of plant cells by 5MT at the molecular level.
Characterization of the differentially expressed gene
In order to isolate differentially expressed genes between 5MT resistant and sensitive plants, 223 randomly selected clones were analyzed with a differential hybridization met-hod. About 200 ng of DNA was blotted onto three positive nylon membranes using a vacuum dot-blot apparatus. Dif-ferential screening hybridization must be performed at lim-iting (rather than excess) probe concentrations so as to be able to detect differences in the levels of a mRNA between different populations.
Twenty-six clones were represented as differentially exp-ressed genes between 5MT resistant and sensitive plants under 5MT treatment. Nine out of the 26 clones were
high-ly expressed in 5MT resistant plants with 5MT treatment than without 5MT (Fig. 1). Putative identification of twenty -six clones was conducted by a single pass sequencing and BLAST algorithm program. Five of the 26 clones showed a higher expression of more than 2 folds in the 5MT resistant plants treated with 5MT than those non-treated. Table 2 summarizes the details of each of the 5 isolated clones,
Table 1. Information of cDNA libraries from 5MT resistant
mu-tant rice
Average insert size 1.14 kbp (0.35~2.5 kbp) Successful sequencing reads (ESTs) 502 Anomalous sequence (excluded) 29 ESTs with homology matches 413 ESTs with e-value ¤1×10-4 452
Unmatched (unknown) ESTs 89 ESTs of mammalian (host cell) origin 39
Fig. 1. Differential hybridization of the cDNA library from leaves
and roots of 5MT resistant rice mutants. A positive nylon membrane containing randomly selected clones was hy-bridized with digoxigenin-labeled cDNAs from A: 5MT resistant plants without 5MT treatment, B: 5MT resistant plants with 5MT treatment, and C: 5MT sensitive plants with 5MT treatment. A B C A B C D E F G H A B C D E F G H A B C D E F G H 1 2 3 4 5 6 7 8 9 10 11 12 1 2 3 4 5 6 7 8 9 10 11 12 1 2 3 4 5 6 7 8 9 10 11 12
which were assigned putative identities with scores bet-ween 57.8 and 411 using blast algorithms. The results of the identification of differentially expressed genes showed that a few genes were correlated with tolerance to environ-ment stress conditions.
Among the clones characterized, clone RK02 code for a protein that shows a high degree of homology with a heat shock protein 82 of rice, acts as protective agent, and some are localized in the nucleus or chloroplast HSPs which may include an enzyme associated with a mechanism of a thermal tolerance, such as a membrane lipid saturation for protein stability (Van Breusegem et al. 1994). They also include widely different environmental stresses or condi-tions, including water deficit, ABA treatment, wounding, and salinity.
Clone RF09 showed 411 scores and 99% identities to nucleotide sequence of polyubiquitin (CAA53665) of Oryza sativa. Ubiquitin is encoded by small multigenic families containing two types of genes, polyubiquitin and ubiquitin extension / fusion genes. Both types of genes are translated as a poly protein precursor, and then proteolytically pro-cessed to the ubiquitin monomers for an ubiquitin function (Callis and Vierstra 1989). The best-characterized function of ubiquitin is its conjugation to target proteins as a recog-nition signal for a protein degradation (Hochstrasser 1996). Ubiquitin is also involved in other cellular processes, such as a ribosome biosynthesis, chromatin structure and cell cycle control. Differential expression of ubiquitin genes has been documented is response to injury, hormone treatment, heavy metal treatment, pathogen injection, and a heat-shocked treatment (Christensen et al. 1992; Liu et al. 1995).
One clone was identified as hydroxyproline-rich glyco-protein (RJ05) involved in a structural stabilization of the cell wall. Characterization of this class of genes suggested
that they might function as a repression of 5MT absorption or protection cell wall from a protein degradation by 5MT.
In the category ‘energy metabolism’, with small overall increases, the nature of the transcripts, compared to the unstressed state, changes under stress conditions. Differ-ential expressions of enolase (2-phosphoglycerate dehy-dratase) (2-phosphoglycerate hydrolyase) (RM02) in res-ponse to 5MT, might be due to the rice cells increased Gly-colysis, which contribute to ATP production, as a compen-sation for energy levels reduced by 5MT. Enolase is also involved in a phosphoenolpyruvate (PEP) formation from 2- phosphoglycerate.
Clone RA12 was identified as putative chorismate mu-tase/prephenate dehydratase (AAC73018) of Arabidopsis thaliana with 307 bits score and 50% of identities, an en-zyme which converts chorismate into prephenate in the biosynthesis of tyrosine and phenylalanine.
Several clones were highly expressed in 5MT non-treated resistant plants more than in 5MT treated resistant plants, these results were considered to be that 5MT non-treated resistant plants correlatively had a good growth compared with the resistant plants with the inhibition of 5MT.
Northern blot analysis
The expression of five genes among the isolated genes in randomly selected ESTs and differential hybridization anal-yses was examined by a Northern blot analysis with RNA prepared from control plants and 5MT resistant M4lines with time periods of 3, 6, 12, 24, and 48h under the stresses of two tryptophan analogs (5MT and α-MT) and abiotic str-esses (ABA and NaCl). The analyzed genes were designat-ed as OsPU1, OsHRP1, OsHSP1, OsEno1, and OsCM1, encoding polyubiquitin, hydroxyproline rich protein, heat shock protein 82, enolase (2-phosphoglycerate dehydratase)
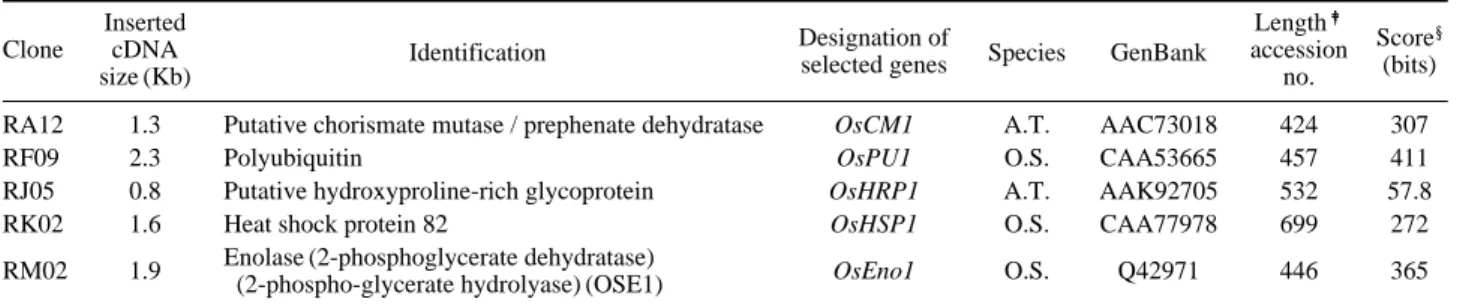
Table 2. Characterization of 5 selected clones differentially expressed in 5MT resistant mutant
Inserted
Designation of Length
�
Score§
Clone cDNA Identification selected genes Species GenBank accession (bits)
size (Kb) no.
RA12 1.3 Putative chorismate mutase / prephenate dehydratase OsCM1 A.T. AAC73018 424 307
RF09 2.3 Polyubiquitin OsPU1 O.S. CAA53665 457 411
RJ05 0.8 Putative hydroxyproline-rich glycoprotein OsHRP1 A.T. AAK92705 532 57.8
RK02 1.6 Heat shock protein 82 OsHSP1 O.S. CAA77978 699 272
RM02 1.9 Enolase (2-phosphoglycerate dehydratase) OsEno1 O.S. Q42971 446 365 (2-phospho-glycerate hydrolyase) (OSE1)
(2-phosphoglycerate hydrolyase) (OSE1), and chorismate mutase, respectively (Table 2). These gene products, either directly or indirectly, involved in functional adaptation me-chanisms under various environmental stress conditions such as a water deficit, ABA treatment, wounding, salinity and a heat-shocked treatment. Chorismate mutase is direct-ly involved in an aromatic amino acid biosynthesis, phenyl-alanine and tyrosine, by catalyzing a conversion from cho-rismate into prephenate.
5MT treatment
The temporal expression patterns of each OsPU1, Os-HRP1, OsHSP1, OsEno1, and OsCM1 gene to the 5MT treatment were analyzed (Fig. 2). The temporal expression patterns of three genes, OsPU1, OsEno1, and OsCM1, in mutants were observed at an early stage of 5MT treatment and compared to the control plants. However, the control plants revealed a rapid expression pattern of OsHSP1 gene (Fig. 2). The expression of OsHRP1 in the control plants
Fig. 2. Northern blot analysis of transcription of the OsPU1, OsHRP1, OsHSP1, OsEno1, and OsCM1 after 500µM 5MT treatment for the indicated time periods. Total RNAs (20µg per lane) from 4-week-old leaves in control (the original variety, Donganbyeo) and 5MT resistant M4mutants were fractionated on a 1% denaturing agarose gel.
Fig. 3. Northern blot analysis of transcriptions of the OsPU1, OsHRP1, OsHSP1, OsEno1, and OsCM1 after 200µM αMT treatment for the indicated time periods. Total RNAs (20µg per lane) from 4-week-old leaves in control (the original variety, Donganbyeo) and 5MT resistant M4mutants were fractionated on a 1% denaturing agarose gel.
Control Mutant 5MT 5MT Non 3 6 12 24 48 h Non 3 6 12 24 48 h OsPU1 OsHRP1 OsHSP1 OsEno1 OsCM1 rRNA OsPU1 OsHRP1 OsHSP1 OsEno1 OsCM1 rRNA Control Mutant αMT αMT Non 3 6 12 24 48 h Non 3 6 12 24 48 h
increased at 6 h after 5MT treatment, remained stable for 24 h and then decreased for 48 h treatment. However, the tran-script levels of this gene in mutants were continuously increased to a 24 h treatment and decreased at a 48 h treat-ment. The transcription levels of OsPU1, OsEno1, and Os-CM1 in 5MT resistant plants were extremely high com-pared to the control plants.
αα-MT treatment
Transcript levels of 5 genes responding to the α-MT (200 µM) treatment were evaluated. OsHRP1 and OsCM1 m-RNA transcript levels revealed similar temporal patterns between the mutants and the control plants, but the expres-sion of OsHRP1 was strongly induced 3 h after an exposure to α-MT and OsCM1 remained high from 6 h to 24 h after the treatment in the mutant plants (Fig. 3). The transcription levels of OsHSP1 mRNA were about the same between the control plants and the mutants, but the mutants showed a rapid expression pattern. In this experiment, the OsEno1 mRNA levels remained constant until 48 h after a treatment in the control plants, while its expressions were gradually increased until 24 h after a treatment and decreased at a 48 h treatment in the mutants.
ABA treatment
It is known that ABA induces many genes and proteins that are also induced by salt and osmotic stresses. In the
present study, the short-term effects of an exogeneous treat-ment of ABA (10µM) on the control and 5MT resistant mutants were examined.
As shown in Fig. 4, the steady-state level of the OsPU1 mRNA transcript had already increased markedly in the mutants after an exposure to ABA. The maximal level was obtained after a 24 h treatment. In the control plants, there was only a minor transient increase in the OsPU1 mRNA transcript level at 6 h, followed by a small and stable incre-ase in the later stages of the ABA treatment. The OsHRP1 transcript levels showed a transient induction with a maxi-mal value at 6 h followed by a decrease to the basal levels between 24 h and 48 h in the mutants. On the other hand, in the control plants, there was only a minor increase in the OsHRP1 mRNA transcript level at 6 h, followed by a stea-dy-state expression by a 48 h treatment. In the OsHSP1 and OsEno1 expression patterns, there was a significant differ-ence between the control plants and the mutants. The re-markable increases of the OsHSP1 and OsEno1 mRNA transcript levels were observed so far as the ABA non-treat-ed mutant is concernnon-treat-ed and continually expressnon-treat-ed during the time periods examined. However, minor increases of these genes were observed by a 24 h treatment in the cont-rol plants.
NaCl treatment
Salt stress is a complex stress imposing a water deficit
Fig. 4. Northern-blot analysis of transcriptions of the OsPU1, OsHRP1, OsHSP1, OsEno1, and OsCM1 after 10µM ABA treatment for the indicated time periods. Total RNAs (20µg per lane) from 4-week-old leaves in control (the original variety, Donganbyeo) and 5MT resistant M4mutants were fractionated on a 1% denaturing agarose gel.
OsPU1 OsHRP1 OsHSP1 OsEno1 OsCM1 rRNA Control Mutant ABA ABA Non 3 6 12 24 48 h Non 3 6 12 24 48 h
because of osmotic effects and it exerts ion-specific toxic effects. Salinity stress is caused a steady accumulation of the OsPU1 transcript in the mutant shoots, and its level reached a peak at 24 h, and then slightly declined. In the control plant, although the temporal pattern of this tran-script was similar to that of the mutants, its trantran-script levels were very weak (Fig. 5).
The OsHSP1 mRNA transcript levels showed a remark-able increase for all the treatment times on the mutants as shown in the ABA treatment, but these transcript levels were slightly increased at 48 h in the control. Exogenous application of NaCl also affected the expression of OsEno1. Its transcript level reached a peak at 48 h and 24 h in the control and mutant, respectively. It is of interest that the salinity treatment did not really affect the OsHRP1 trans-cript level, which is different with the case of the ABA treatment.
The increase of the OsPU1 transcript level indicates its conjugation to target proteins as a recognition signal for a protein degradation. Fraser et al. (1991) reported that poly-ubiquitin genes are transiently induced as an adaptation of a cell to environmental changes so as to ensure a program-ming of a cellular metabolism and a rapid turnover of re-gulatory or enzymatically active proteins which are no lon-ger necessary under the new conditions. In this experiment, the OsPU1 transcription levels of the mutant plants showed a remarkable increase under 5MT, ABA, and NaCl
treat-ments.
Glycolysis has a role in the production of a wide range of metabolites including amino acids, lipids, and related com-pounds (Plaxton 1996). Forsthoefel et al. (1995) have shown that an enolase activity increases more than four-fold in leaves during salt stress in ice plant Mesembryanthemum crystallinum. The reason for this increase was accompanied by a dramatic increase in Pgh1, encoding an enolase tran-scription rate. Pgh1 transcript levels also increased in leav-es in rleav-esponse to a low temperature, drought, and anaerobic stress conditions. Recently, Minhas and Grover (1999) re-ported that the eno mRNA increased in response to a desic-cation stress in both shoot and root tissues of rice. Increase of enolase (OsEno1) transcript levels might be due to the rice cells increased glycolysis rates to compensate for re-duced energy levels brought about by 5MT or other stres-ses. According to Plaxton (1996), the cytosolic network of glycolytic enzymes may provide an essential metabolic flexibility that facilitates in a plant development and accli-mation to an environmental stress. In addition, an increase of enolase promotes the synthesis of phosphoenolpyruvate (PEP) by a catalyzing the conversion 2-phosphoglycerate into PEP. PEP synthesizes shikimic acid, a precursor of chorismate in the shikimate pathway, by combination with erythrose-4-phosphate. Successively, the shikimic acid fin-ally synthesizes aromatic amino acids such as tryptophan, phenylalanine, and tyrosine.
Fig. 5. Northern blot analysis of transcriptions of the OsPU1, OsHRP1, OsHSP1, OsEno1, and OsCM1 after 200 mM NaCl treatment for the
indicated time periods. Total RNAs (20µg per lane) from 4-week-old leaves in control (the original variety, Donganbyeo) and 5MT resistant M4mutants were fractionated on a 1% denaturing agarose gel.
Control Mutant NaCl NaCl Non 3 6 12 24 48 h Non 3 6 12 24 48 h OsPU1 OsHRP1 OsHSP1 OsEno1 OsCM1 rRNA
In the transcript levels of OsHSP1, there was no differen-ce between the control and mutant plants by 5MT. How-ever, this gene was highly expressed under abiotic stresses, ABA and NaCl. A primary function of the hsp family is a molecular chaperone guiding an initial folding of proteins, and a refolding of unfolded proteins (Nelson et al. 1992). Heat shock protein acts to prevent a protein aggregation and the adverse effects that would follow under the con-ditions of a heat shock. A second function of a heat shock proteins involves cellular recovery from heat shock or other protein-denaturing stresses. The presence of a heat shock protein is required to protect the functional domains of denatured proteins. The protection provided by the heat shock protein binding offers an opportunity for refolding (Becker and Craing 1994).
Considerable efforts have been made in recent years to identify novel and potentially useful genes involved in a molecular basis of a systemic acquired resistance. The data presented in this study demonstrates the potential value of the 5MT resistant mutants for an identification of novel cDNAs for defense and stress related proteins.
We usually considered an isolation from other stresses to simplify the interpretation. In nature, however, a plant en-counters stress combinations and presents an integrated response to them. The signal transduction pathways trig-gered by different stresses are common to more than one stress type. One possible reason for this is that, under certain conditions, these two stresses cannot be distinguish-ed from one another. Alternatively each stress might re-quire the same protective action. Knight and Knight (2001) defined ‘cross-talk’, abiotic stress signaling networks, as an instance of two signaling pathways from different stressors that converge. This might take the form of different path-ways achieving the same end or of pathpath-ways integrating and affecting each other’s outcome, including the flux through one pathway affecting another. These might act in an additive or negatively regulatory way, or might compete for a target. Cross-talk can also occur between pathways, in different organs of the plant when a systemic signal such as a hydrogen peroxide moves from a stimulated cell into another tissue to elicit a response (Foyer et al. 1997).
Analyses of differential hybridization and Northern blot from 5MT resistant mutant cDNA library identified expres-sion of many genes related to a defense and stress response. These results demonstrate presence of the cross-talk
bet-ween 5MT and other stresses, and the possibility of the presence of other resistance mechanisms similar to in abiotic stresses besides resistance mechanisms for an amino acid analog reported up to date. Further works are required for identification of a physiological response and resistance mechanism of a plant in response to an amino acid analog related to other stresses.
ACKNOWLEDGEMENTS
This work was supported by a grant (Code 20070501034 005) from BioGreen 21 Program, RDA (Rural Develop-ment Administration), and from the KOSEF (Korean Scien-ce and Engineering Foundation) in the MOST (Ministry of Science and Technology), Republic of Korea.
REFERENCES
Becker J and Craing EA. 1994. Heat-shock protein as mole-cular chaperones. Eur. J. Biochem. 219:11-23.
Brown PO and Botstein D. 1999. Exploring the new world of the genome with DNA microarrays. Nat. Genet. 21:33-37. Callis J and Vierstra RD. 1989. Ubiquitin and ubiquitin genes
in higher plants, Oxford Surv. Plant Mol. Cell Biol. 6:1-30. Christensen AH, Sharrock RA and Quail PH. 1992. Maize polyubiquitin genes: structure, thermal perturbation of exp-ression and transcript splicing, and promoter activity fol-lowing transfer to protoplasts by electroporation. Plant
Mol. Biol. 18:675-689.
Cushman JC and Bohnert HJ. 2000. Genomic approaches to plant stress tolerance. Curr. Opin. Plant Biol. 3:117-124. Desprez T, Amselem J, Caboche M and Hofte H. 1998.
Differ-ential gene expression in Arabidopsis monitored using cDNA arrays. Plant J. 14:643-652.
Forsthoefel NR, Cushman MAF and Cushman JC. 1995. Posttranscriptional and posttranslational control of enolase expression in the facultative Crassulacean acid metabolism plant Mesembryanthemum crystallinum L. Plant Physiol.
108:1185-1195.
Foyer CH, Lopez-Delgado H, Dat JH and Scott IM. 1997. Hydrogen peroxide and glutathione-associated mechani-sms of acclimatory stress tolerance and signaling. Physiol.
Plant 100:241-254.
Fraser J, Luu HA, Neculcea J, Thomas DY and Storms RK. 1991. Ubiquitin gene expression: response to
environmen-tal changes. Curr. Genet. 20:17-23.
Hochstrasser M. 1996. Ubiquitin-dependent protein degrada-tion. Annu. Rev. Genet. 30:405-439.
Joshi CD. 1987. An inspection of the domain between puta-tive TATA box and translation start site in 79 plants.
Nu-cleic Acids Res. 15:6643-6653.
Kim DS, Lee IS, Jang CS, Kang SY, Park IS, Song HS and Seo YW. 2005a High amino acid accumulating 5-methyl-tryptophan resistant rice mutants may include an increased antioxidative response system. Physiol. Plant 123: 302-313. Kim DS, Lee IS, Jang CS, Kang SY, Song HS and Seo YW. 2005b Altered anthranilate synthase in 5-methyltryptophan resistant rice mutants. Plant Cell Rep. 24:357-365. Knight H and Knight MR. 2001. Abiotic stress signaling
path-ways: specificity and cross-talk. Trends Plant Sci. 6:262-267.
Li J and Last RL. 1996. The Arabidopsis thaliana trp5 mutant has a feedback-resistant anthranilate synthase and elevated soluble tryptophan. Plant Physiol. 110:51-59.
Liu L, Maillet DS, Frappier JR, Walden DB and Atkinson BG. 1995. Characterization, chromosomal mapping, and expre-ssion of different polyubiquitin genes in tissues from con-trol and heat-shocked maize seedlings. Biochem Cell Biol.
73:19-30.
Minhas D and Grover A. 1999. Transcript levels of genes en-coding various glycolytic and fermentation enzymes chan-ge in response to abiotic stresses. Plant Sci. 146:41-51. Nelson RJ, Ziegelhoffer T, Nicolet C, Werner-Washburne M
and Craing EA. 1992. The translation a machinery and 70 kd heat shock protein cooperate in protein synthesis. Cell
71:97-105.
Plaxton WC. 1996. The organization and regulation of plant
glycolysis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47: 185-214.
Stanton LW. 2001. Methods to profile gene expression. Trends
Cardiovasc. Med. 11:49-54.
Takemoto D, Doke N and Kawakita K. 2001 Characterization of elicitor-inducible tobacco genes isolated by differential hybridization. J. Gen. Plant Pathol. 67:89-96.
Ueda A, Shi W, Nakamura T and Takabe T. 2002. Analysis of salt-inducible genes in barley roots by differential display.
J. Plant Res. 115:119-130.
Van Breusegem F, Dekeyser R, Garcia AB, Claes B, Gielen J, Van Montagu and Caplan AB. 1994. heat-inducible rice
hsp82 and hsp70 are not always co-regulated. Planta 193:
57-66.
Wakasa K. 1985. Mutation selection in vitro in rice and tobac-co. In: Reddy GM, Coe EH Jr. (eds) Gene structure and function in higher plants Oxford & IBH, New Delhi, India, pp. 247-251.
Widholm JM. 1971. Control of tryptophan biosynthesis in plant tissue cultures: lack of repression of anthranilate and tryptophan synthetases by tryptophan. Physiol. Plant 25: 75-79.
Yu H and Goh CJ. 2000. Differential gene expression during floral transition in an orchid hybrid Dendrobium Madame Thong-In. Plant Cell Rep. 19:926-931.
Zhang L, Ma X-L, Zhang Q, Ma C-L, Wang P-P, Sun Y-F, Zhao Y-X and Zhang H. 2001. Expressed sequence tags from a NaCl-treated Suaeda salsa cDNA library. Gene 267 :193-200.
Manuscript Received: October 15, 2007 Revision Accepted: November 7, 2007