0°C
이하의 저온 스트레스(Freezing stress)는 식물체가 0°C
이하의 온도 조건에 노출되었을 때 발생하는 비생물학적
스트레스의 일종이다. 식물이 0°C 이하의 저온에 노출되면
세포 간극의 수분이 얼게 되면서 얼음결정(ice formation)
을 생성하게 되고 이러한 얼음 결정은 삼투 스트레스를 일
으켜 세포 내 탈수 스트레스를 유발 한다(Trischuk et al.,
2006; Sakai et al., 2012). 또한 0°C 이하의 저온 스트레스
는 식물 세포의 원형질막의 유동성을 떨어뜨려 안정성을 저
해시키며, 외부자극에 반응하는 통로를 차단하거나 단백질
파괴 등을 유발한다(Ruelland et al., 2009). 이로 인해 식물
은 생장 및 발달과 작물의 생산성 및 품질에 심각한 피해를
입게 된다(Sanghera et al., 2011; Pu et al., 2019).
유채 두 계통에서 저온 스트레스에 반응하는 전사체 발현 비교 분석
이지은
1,†・ 김광수
1・ 차영록
1・ 안다희
1・ 변종원
2・ 강용구
3Comparative Transcriptome Analysis of the Response of Two Lines of Rapeseed
(Brassica napus L.) to Cold Stress
Ji-Eun Lee
1,†, Kwang-Soo Kim
1, Young-Lok Cha
1, Da-Hee An
1, Jong-Won Byun
2, and Yong-Ku Kang
3ABSTRACT Rapeseed is a typical winter crop, and its freezing stress tolerance is a major feature for winter survival. Therefore,
it is important to comprehend clearly the physical and molecular mechanisms of rapeseed under freezing stress conditions. This
study investigates the physical and transcriptome changes of two rapeseed lines, ‘J8634-B-30’ and ‘EMS26’, under cold
acclimation and freezing temperature treatments. The proline content of ‘J8634-B-30’ at 5 °C increased 8.7-fold compared to that
before treatment, and there was no significant change in that of ‘EMS26’ RNA-sequencing analysis revealed 5,083 differentially
expressed genes (DEGs) of ‘J8634-B-30’ under cold acclimation condition. Among the genes, 2,784 (54.8%) were up-regulated
and 2,299 (45.2%) were down-regulated. The DEGs of ‘EMS26’ under cold acclimation condition were 5,831 genes, and
contained 2,199 up-regulated genes (37.7%) and 3,632 down-regulated genes (62.3%). Among them, only DEGs annotated in the
cold response-related signaling pathways were selected, and their expression in the two rapeseed lines was compared.
Comparative DEGs analysis indicated that cold response related signaling pathways are proline metabolism and ABA (Abscisic
acid) signaling. And ICE (Inducer of CBF expression) - CBF (C-repeat-binding factor) - COR (Cold-regulated) signaling were the
significantly differentially expressed transcripts in the two rapeseed lines. The major induced transcripts of ‘J8634-B-30’ induced
P5CS (Δ‘-pyrroline-5-carboxylate synthetase), which is related to proline biosynthesis, PYL (pyrabactin resistance-like protein,
ABA receptor) and COR413 (cold-regulated 413 plasma membrane 1). In conclusion, these result provide a foundation for
understanding the mechanisms of freezing stress tolerance in rapeseeds. Further functional studies should be performed on the
freezing stress-related genes identified in this study, which can contribute to the transgenic and molecular breeding for freezing
stress tolerance in rapeseed.
Keywords : cold acclimation, freezing stress, rapeseed, transcriptome analysis
Original Research Article
ⓒ 본 학회지의 저작권은 한국작물학회지에 있으며, 이의 무단전재나 복제를 금합니다.
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
1)농촌진흥청 국립식량과학원 바이오에너지작물연구소 농업연구사 (Researcher, Bioenergy Crop Research Institute, National Institute of Crop
Science, RDA, 199 Muanro, Cheonggye, Muan 58545, Republic of Korea)
2)농촌진흥청 국립식량과학원 바이오에너지작물연구소 박사 후 연구원 (Postdoc, Bioenergy Crop Research Institute, National Institute of Crop
Science, RDA, 199 Muanro, Cheonggye, Muan 58545, Republic of Korea)
3)농촌진흥청 국립식량과학원 바이오에너지작물연구소 농업연구관 (Senior Researcher, Bioenergy Crop Research Institute, National Institute
of Crop Science, RDA, 199 Muanro, Cheonggye, Muan 58545, Republic of Korea)
†
Corresponding author: Ji-Eun Lee; (Phone) +82-61-450-0137; (E-mail) [email protected]
많은 식물들은 동해 전 0°C 이상의 일정한 저온에 노출
되게 되면 내동성이 증가하게 되는 데, 이러한 현상은 ‘저온
순화(cold acclimation)’라고 보고되었다(Thomashow, 1999).
저온 순화는 매우 복잡한 과정으로, 수백 개의 유전자 발현
변화를 통해 많은 생화학 및 생리적 변화를 유발한다(Guy,
1990; Xin & Browse, 2000; Shi et al., 2018.). 저온 순화로
인한 변화는 식물의 생장을 억제하여 조직 내 수분 함량을
감소시키고, ABA (Abscisic acid) 호르몬 함량을 증가시키
며, 막 지질 구성요소를 변화시키고, proline, betaine, soluble
sugars와 같은 삼투 스트레스에 저항할 수 있는 물질을 축적
시킨다(Levitt, 1980; Chen et al., 1983; Lynch & Steponkus,
1987; Koster & Lynch, 1992; TaO et al., 1998).
Proline은 식물체 내에서 많은 스트레스에 반응하며 함량
이 증가된다. Proline은 비생물학적 스트레스를 막기 위해
식물체 내에서 생합성되고, 세포질 내 축적되어 molecular
chaperon으로써 단백질 구조를 안정화 시키고, 세포질 pH에
대해 완충 역할을 하며 세포 내 산화환원 반응을 유지시켜,
세포 파괴를 막는다고 알려져 있다(Verbruggen & Hermans,
2008; Szabados & Amould, 2010; Hayat et al., 2012). 또
한 ABA 호르몬은 대표적인 스트레스에 반응하는 식물 호
르몬으로 기공의 개폐 조절 등에 관여하고 생장과 발달에
영향을 미치며, 가뭄, 저온, 고온 등의 스트레스 조건에서
식물이 살아남을 수 있는 저항성을 갖게 한다(Shinozaki &
Yamaguch-Shinozaki, 2000; Schroeder et al., 2001). 또한
이러한 저온 순화에 기여하는 저온 반응 유전자 발현을 조
절하는 기작으로는 ICE (Inducer of CBF expression) - CBF
(C-repeat-binding factor) - COR (Cold-regulated) 신호 경
로가 있다(Shi et al., 2018). 저온에 감응하여 과발현된 ICE
는 CBF와 결합하여 COR 유전자를 발현시키고 저온 순화
와 관련된 COR 유전자의 downstream 유전자들이 발현되
어 내동성을 갖는다고 보고되었다(Xin & Browse, 2000;
Chinnusamy et al., 2007; Ritonga & Chen, 2020).
유채는 전세계적으로 소비되는 카놀라(canola)유를 생산
하는 작물로, 중국, 유럽, 우리나라 등에서는 가을에 재배하
여 이듬해 봄, 여름에 수확하는 동계작물이다(Burbulis et al.,
2011; Pu et al., 2019; Xin et al., 2019). 동계작물인 유채
는 생육기간 중 저온과 0°C 이하의 저온 스트레스에 노출
될 수밖에 없는데, 이와 관련된 여러 연구들이 보고되었다.
기존 연구 결과에서 유채 10 품종을 대상으로 한 0°C 이하
의 저온 노출 기간에 따른 생존율 등의 조사에서 내성을 갖
는 ‘Tosca’ 품종을 선발하였다(Waalen et al., 2011). 또한 7
개 유채 품종을 대상으로 4°C 저온 순화 기간에 따라 내동성
을 확인한 결과, proline 및 soluble sugar 함량이 증가함을
확인하였고 ‘Valesca’ 품종에서 이온 누출도가 낮고 생장 피
해를 덜 입는 다는 결과가 보고되었다(Burbulis et al., 2011).
유채의 이러한 내동성 기작에 관한 연구는 차세대염기서
열분석(Next generation sequencing) 기술이 발달하여 염기
서열 연구 변화가 대량화됨에 따라 염기서열 연구 분야에
서도 수행되어왔다. 최근 연구에서는 중국의 유채 계통인
‘2016TS(G)10’을 대상으로 저온 및 0°C 이하의 저온 스트
레스를 처리한 결과, 저온 순화 조건에서 soluble sugar,
proline, MDA (Malondialdehyde) 함량이 증가하고 세포 내
전분이 축적되는 결과를 확인하였으며, 이를 바탕으로 저
온 순화 조건에서 전사체 분석 연구가 수행되어 관련 기작
에 대해 보고되었다(Pu et al., 2019). 또한 유채 ‘HX17’과
‘HX58’ 품종의 서로 다른 내동성을 확인하고 두 품종을 저
온(4°C)과 0°C 이하의 저온(-4°C) 스트레스에 노출 시켰을
때 전사체의 발현 양상을 비교하는 연구도 수행되었다(Xin
et al., 2019). 국내에서도 국내 육성 5개의 유채 품종의 내
동성을 확인하였으며, microarray 분석을 통해 내성을 갖는
‘내한’ 품종과 중간 내성 ‘영산’, 민감성 ‘탐미’ 품종의 발현
된 유전자군을 비교한 연구가 수행되었다(Jeong et al., 2012).
따라서 본 연구는 국내에서 육성한 유채 2 계통 ‘J8634-
B-30’과 ‘EMS26’을 대상으로 0°C 이하의 저온 스트레스
를 처리하여 이에 따른 생리적 변화를 확인하고 이와 관련
된 저온 순화 시 유전자들의 발현 양상을 전사체 수준에서
확인하여, 국내 유채 계통의 내동성 향상을 위한 저온순화
기작을 이해함으로써 내동성을 갖는 유채 품종 육종에 기
초 자료로 활용하기 위해 수행되었다.
재료 및 방법
실험재료 및 저온 처리
본 시험에 사용된 유채 계통은 국립식량과학원에서 육성
한 ‘J8634-B-30’과 ‘EMS26’이었다. ‘J8634-B-30’ 유채 계
통은 기존 내동성 검정 시험 결과 내동성 계통으로 선발된
계통이었으며, ‘EMS26’ 유채 계통은 유채 ‘탐라’ 계통에
EMS (Ethyl Methane Sulfonate)를 처리하여 인위적인 돌연
변이를 유도하고 자가 수정을 통해 6세대까지 세대를 전개
시킨 계통으로 지방산 조성 중 올레산 함량이 70%가 넘는
우수한 식용유 품질을 갖는 계통이다.
유채 두 계통을 대상으로 각 계통 당 72공 트레이에 12
공(1립씩 파종, 1 개체)을 한 세트로 5세트(전체 60 개체)를
준비하여 스트레스를 처리하였다. 이 중 2 세트(24 개체)에
서 처리 전후 proline 분석을 위해 지상부 3 개체 시료(총
15 개체)를 채취하여 각 시점 당 3 반복으로 사용하였으며,
RNA 분석을 위하여 처리 전, 후 2 개체 시료(총 4 개체)를
반복으로 사용하였다. 남은 3세트는 12 개체를 기준으로 스
트레스 처리 후 생존율 확인을 위한 3반복으로 사용하였다.
0°C 이하의 저온 스트레스 처리 조건은 Jeong et al. (2012)
을 참조하여 다음과 같이 변형하여 처리하였다(Fig. S1).
0°C 이하의 저온 스트레스 처리는 15°C 조건에서 생육한 본
엽 3∼4기의 유채를 5°C에서 24시간 동안 저온 순화(Cold
acclimation) 시킨 후, 2시간에 1°C씩 온도를 내려 -9°C에
서 24시간 동안 0°C 이하의 저온 스트레스 처리(Freezing
stress)하였다. 이후 2시간에 1°C씩 15°C까지 온도를 올린
후 저온 처리를 마무리하였다. Proline 등을 조사하기 위해
처리전(15°C), 저온순화 후(5°C), 온도 감온 2시기(0°C와
-5°C), 0°C 이하의 저온 스트레스 처리 후(-9°C) 유채 지상
부를 채취하였다. RNA seq 분석을 위한 시료는 처리전
(15°C)과 저온순화 후(5°C) 유채 지상부를 채취하였다.
생존율은 0°C 이하의 저온 스트레스 처리 종료 일주일
및 한 달 후 살아남은 유채 개체를 조사하여 확인하였다.
생존율은 12 개체(1세트)를 1 반복으로 다음과 같이 3반복
으로 조사하였다: [(일주일 혹은 한 달 후 살아남은 개체)/
처리 전 개체(12 개체)] × 100.
유채 Proline 분석
유채 proline 함량은 ninhydrin 방법을 변형하여 측정하였
다(McClinchey et al., 2008). 지상부 시료 0.3 g에 증류수
를 넣고 100°C에서 30분간 가열 후, 상등액 200 μl과 acid
ninhydrin (ninhydrin 0.25 g + acetic acid 15 ml + 증류수
10 ml) 1 ml, 증류수 400 ml를 넣고 20분간 100°C에서 가
열하였다. 가열된 분석액을 냉각시킨 후 toluene 3 ml를 첨
가하여 암조건에서 2시간 반응시켰다. 이후 520 nm 흡광도
를 측정하였으며, L-proline으로 표준곡선을 작성한 후 proline
함량은 3 반복으로 계산하였다. Proline 함량 간의 평균에
대한 통계분석은 ANOVA 분석 후, Duncan 다중 검정을
실시하였다(R version 3.6.1).
Total RNA 분리 및 RNA-Seq 분석
Total RNA는 채취한 유채 지상부 0.1 g의 시료를 Mini
BEST Plant RNA extraction kit (TaKaRa Bio Inc, Japan)를
이용하여 분리하였고, Bioanalyzer (Agilent Technologies,
USA)를 사용하여 농도를 측정하다. 분리된 total RNA 2 μg
은 Illumina TruSeq RNA Sample Preparation Kit를 사용하
여 300 bp insert size로 RNA-Seq library를 제작하였고,
Illumina HiSeq X platform (Illumina, SanDiego, CA)을 이
용하여 101 bp paired-end reads를 생성하였다(Macrogen
Co, Seoul, Korea). 낮은 품질과 duplicated된 reads, adapter
sequence들은 Trimmomatic (ver. 0.38, Bolger et al., 2014)
를 이용하여 제거하였다.
유전자 발현 및 DEG 분석
정제된 RNA-Seq read들은 HISAT2 (Kim et al., 2015) 프
로그램을 이용하여 유채 표준유전체 서열(v4.1, http://www.
genoscope.cns.fr/brassicanapus/, Chalhoub et al., 2014)에
맵핑하였다. 이후 HTSeq-count (Anders et al., 2015)를 사
용하여 단백질 코딩 서열에 맵핑된 RNA read들을 계산하
였고, 이를 FPKM (Fragment Per Kilobase of transcript per
Million mapped reads) 계산에 사용하였다.
유채 두 계통의 처리전과 저온순화 후 전사체 사이의 DEG
(Differentially-expressed genes)는 각 시료당 2반복으로 비
교하고자 하는 시료의 count 값을 기반으로 Bioconductor
package DESeq (Anders & Huber, 2010)를 이용하여 DESeq
Normalization 값을 산출하였다. 이를 이용하여 산출된 유
전자들 중 P-value 값을 Benjamini-Hochberg의 방법으로
조절한 FDR (False Discovery Rate) 값을 이용하여 보정된
adjusted P-value 값이 0.05 이하 값을 만족하는 유전자들을
대상으로 Log2 fold change를 사용한 up/down regulation
분석을 실시하였다. 이중 Log2 fold chage 값이 절대값 1
이상이면, 발현량이 2배 이상 차이를 보인다는 것으로 판단
하여 이를 기준으로 DEG로 선별하였다.
유전자 기능 분석
GO (Gene ontology) enrichment 분석은 BLAST2GO 프
로그램(v5.2)을 사용하여 Fisher’s exact test 방법(adjusted
P-value < 0.05)으로 DEG 유전자들을 분석하였으며, p-
value < 0.01, FDR (False discovery rate) < 0.01, 선택된
유전자의 fold change가 전체 유전자 대비 2배 이상의 조건
으로 필터링하였다. 이후 선택된 유전자를 생물학적 기능
(Biological processes), 분자 기능(Molecular function), 그리
고 세포 요소(Cellular component)로 분류하였다. 기능이 확
인된 DEG들을중 Proline metabolism, ABA pathway, 저온
관련 유전자 등을 탐색 하여 선발한 후, MeV (v.4.9.0) 프
로그램을 사용하여 heatmap을 그렸다.
RT-PCR 분석
특이적인 발현이 확인된 DEG들의 유전자 발현을 확인하
기 위하여, 처리전(15°C), 저온순화 후(5°C), 온도 감온시기
(-5°C), 0°C 이하의 저온 스트레스 처리 후(-9°C) 유채 지
상부를 채취하였다. 채취한 유채 지상부는 앞선 방법과 동
일하게 RNA를 분리하였으며, 동량의 RNA를 사용하여 1
ststrand cDNA synthesis kit (TaKaRa Bio Inc, Japan)를 이용
하여 cDNA를 합성하였다. 유전자 발현 확인 위해 선별된
DEG의 primer 목록은 Table S1과 같으며, Bactin은 internal
control로 사용하였다(Lee et al., 2013).
결과 및 고찰
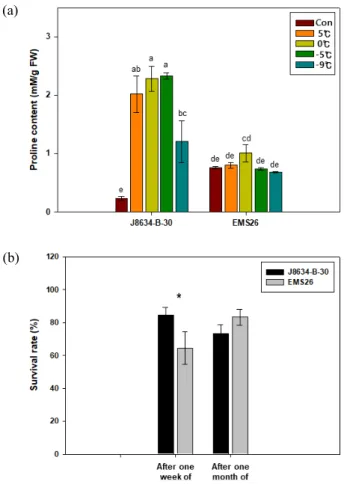
0°C 이하의 저온 스트레스 처리에 따른 유채 proline 함
량 및 생존율 변화
0°C 이하의 저온 스트레스 처리 후 두 유채 계통에서는 proline
함량에서 다른 변화를 보였다(Fig. 1a). 처리 전 proline의
함량은 ‘EMS26’에서 0.76 mM/g FW로 ‘J8634-B-30’ 0.23
mM/g FW보다 약 3.3배 높았다. 그러나 저온 순화를 한
‘J8634-B-30’ 계통에서 proline 함량이 2.02 mM/g FW로 처
리 전보다 8.7배 높아졌으나, ‘EMS26’ 계통에서는 0.80 mM/g
FW으로 차이를 보이지 않았으며 처리 전반적으로 통계적
유의성을 보이지 않았다. 저온 순화 후 0°C에서 ‘J8634-B-30’
계통의 proline 함량은 2.28 mM/g FW, -5°C에서 2.33 mM/g
FW로 높게 유지되었다. 이후 -9°C에서 24시간 처리 후
‘J8634-B-30’ 계통의 proline 함량은 1.21 mM/g FW으로
약간 감소하였다. 다만, 식물체 내에서 식물 0°C 이하의 저
온 스트레스 피해의 지표로 많이 사용되고 있는 엽록소 함
량, 이온 누출도의 경우 두 유채 계통간의 차이는 발견되지
않았다(Data not shown). 이러한 연구 결과는 내한성 유채
품종을 선발하기 위한 기존 연구에서도 proline 함량 증가
만이 0°C 이하의 저온 스트레스 처리 후 높은 생존율과 연
관성이 있다는 보고와 유사한 결과였다(Kim, 2015).
0°C 이하의 저온 스트레스 처리 종료 일주일 후 유채 두
계통의 생존율을 조사한 결과, ‘J8634-B-30’ 계통에서 84.6%
로 ‘EMS26’ 계통의 생존율 64.4% 보다 높아 초기 0°C 이
하의 저온 스트레스 피해에 내성을 보였다(Fig. 1b). 이후 처
리 한 달 뒤 유채 두 계통의 생존율을 조사한 결과, ‘J8634-
B-30’에서 73.4%, ‘EMS26’ 계통에서 83.3%로 차이를 보
이지 않았다. 이는 ‘EMS26’ 계통 중 본엽이 모두 고사하였
다고 판단한 개체에서 새 잎이 생장하여 생존 개체 수가 변
화하였기 때문으로 판단된다. 이러한 결과는 ‘EMS26’ 개체
가 0°C 이하의 저온 스트레스에 대해 장기적인 내동성을 보
여 최종적으로 유채 두 계통에서의 0°C 이하의 저온 스트레
스에 대한 내성이 비슷한 수준으로 나타난 것으로 판단된다.
저온 처리 전후 유채 2 계통 전사체 분석
유채의 초기 내동성과 관련된 기작을 확인하가 위해, 0°C
이하의 저온 스트레스 처리로 proline 함량이 급격한 변화
를 보인 5°C 처리 유채 시료와 처리 전(15°C) 유채 시료의
전사체를 각각 유채 2 계통에서 비교 분석하였다(Table 1).
전체 정제된 read들의 개수는 50,111,336∼60,133,544개였으
며, 전체 read 길이는 7.3∼8.8 Gb였다. Read의 품질을 나
타내는 Q30 이상의 read들은 전체 read들의 98.0∼98.1%
로 나타났으며, 전체 read들의 86.0∼90.5%가 유채 표준유
전체에 맵핑되었다. 발현된 전사체 비율은 전체 표준유전
체의 전사체에 대해 시료 read가 적어도 1개 이상 맵핑된
전사체(유전자)의 비율로써 표준유전체 대비 61.0∼65.3%
가 시료 전사체에서 맵핑되었다.
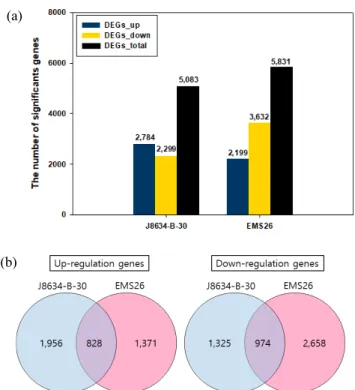
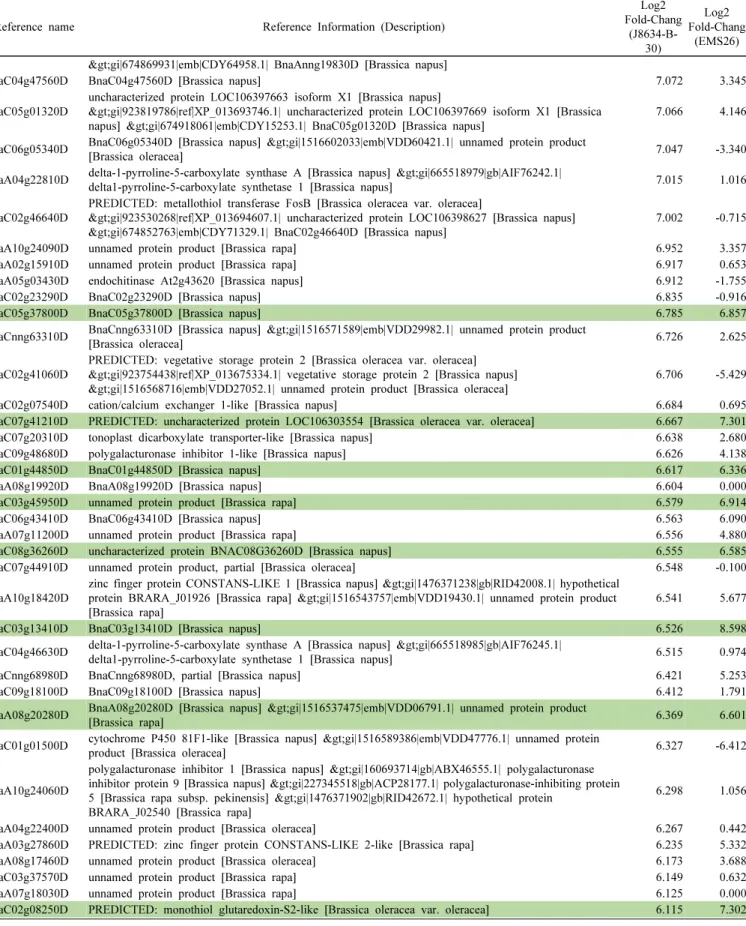
저온 처리 전과 후 전사체의 DEG는 ‘J8634-B-30’과 ‘EMS
26’에서 각각 5,083개, 5,831개였다(Fig. 2a). ‘J8634-B-30’
계통의 DEG는 발현이 유도된 유전자의 수가 2,784개로 발
(a)
(b)
Fig. 1. Changes in proline content (a) and survival rate (b)
of two rapeseed lines according to freezing treatment.
Con (Control) indicates 15°C. Bars represent standard
error. Different lowercase letters indicate significant
difference at P < 0.05 according to Duncan’s multiple
range test. The asterisk indicates significant difference
at P < 0.05 according to T-test.
현이 억제된 유전자 2,299개 보다 많았으나, ‘EMS26’ 계통
의 DEG는 발현이 유도된 유전자 수가 2,199개로, 발현이
억제된 유전자 수 3,632개 보다 적었다. 두 계통에서 공통
적으로 발현이 유도된 DEG는 828개였으며, 발현이 억제된
DEG는 974개였다(Fig. 2b).
Table 1. The alignment statistics data of the reference gene for B. napus samples.
Sample name
Trimmed reads
Read Bases
GC (%)
%≥Q30
Mapped reads
(%)
Expressed genes
(%)
J8634_Con_1
50,111,336
7,343,661,008
46.41
98.08
89.21
65.27
J8634_Con_2
56,626,338
8,269,505,242
46.65
98.08
85.96
64.47
J8634_Treat_1
60,133,544
8,803,705,393
47.68
98.07
90.03
63.85
J8634_Treat_2
52,787,030
7,734,751,081
47.51
98.06
89.81
63.41
EMS26_Con_1
55,220,650
8,101,701,706
46.46
98.08
90.00
64.09
EMS26_Con_2
52,359,812
7,627,197,929
46.60
98.10
90.47
64.88
EMS26_Treat_1
50,938,796
7,444,117,983
46.42
98.09
90.07
61.01
EMS26_Treat_2
54,792,402
8,009,480,373
47.09
98.03
90.47
61.24
*
‘J8634’; ‘J8634-B-30’, Con; Rapeseed RNA sample before cold treatment, Treat; Rapeseeds were kept at 5°C for 24h,
**Trimmed reads; removing a read with an average base quality below 20 and dropping a read less than 50 base long
***%≥Q30; Q-score of trimmed data ≥ 30, Mapped reads; the number of reads mapped to the reference genome, Expressed
genes; the number of transcript with a count of 1 or more, (Expressed transcripts / Total transcript) × 100
(a)
(b)
Fig. 2. Distribution of cold regulated differentially expressed
genes (DEGs) in rapeseed shoots. (a); the number of
DEGs in two rapeseed lines, (b); Venn diagrams
re-present the number of overlap sets of up- or down-
regulated DEGs between J8634-B-30 and EMS26.
(a)
(b)
Fig. 3. GO term enrichments analysis of DEGs induced by
cold treatment in ‘J8634-B-30’ and ‘EMS26’. (a); Up-
regulated DEGs in ‘J8634-B-30’, (b); Up–regulated
DEGs in ‘EMS26. Detailed data are shown in Table
S1 and Table S3.
DEG를 바탕으로 GO 분석을 실시하고 각 주요 GO 3 그
룹별로 분류하여 각 그룹의 상위 5개 GO term을 분석한
결과, ‘J8634-B-30’ 계통에서는 oxidation-dependent protein
catabolic process, lignin 생합성 과정 조절, ribosome
bio-genesis 관련 GO 발현이 높았으며, proline 관련 GO term도
3종류 발현이 높은 것을 확인 할 수 있었다(Fig. 3a, Table S2).
반면, (+)-abscisic acid D-glucopyranosyl ester transmembrane
transport, (+)-abscisic acid D-glucopyranosyl ester transmembrane
transporter activity 등 ABA transport와 관련된 GO term의
발현이 억제되는 것을 확인 할 수 있었다(Table S3).
‘EMS26’ 계통에서는 스트레스에 반응하는 translation 조
절, S-glycoside catabolic 과정 등에서 발현이 높은 양상을
보였으며, proline 관련 GO term은 1 종류에서 발현이 높은
것을 확인 할 수 있었다(Fig. 3b, Table S4). 반면, shikimate
metabolic process, cellular response to sulfur starvation,
shikimate 3-dehydrogenase (NADP+) activity 관련 GO term
의 발현이 억제되는 것을 확인 할 수 있었다(Table S5).
저온 처리로 인한 유채 2 계통 DEG의 기능적 분석(Functional
analysis)
유채 2 계통의 전사체 발현 비교를 위해, 2 계통의 각각의
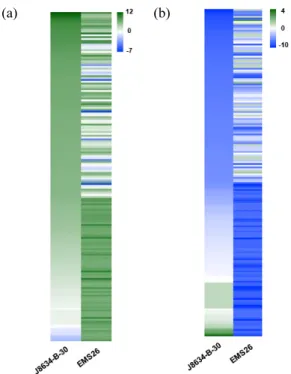
상위 100개 DEG를 선발하고 발현 패턴을 비교하였다(Fig.
4). 발현이 강하게 유도된 각각의 상위 100개의 DEG 중 21
개의 DEG가 공통적으로 상위 100개의 DEG로 선별되어 총
179개의 DEG의 발현을 두 계통에서 비교하였으며(Fig. 4a,
Table S6), 발현이 강하게 억제된 각 계통의 상위 100개
DEG 중 11개의 DEG가 공통적으로 확인되어 총 189개의
DEG의 발현을 두 계통에서 비교하였다(Fig. 4b, Table S7).
발현이 강하게 유도된 각각의 상위 100개의 DEG를 비교
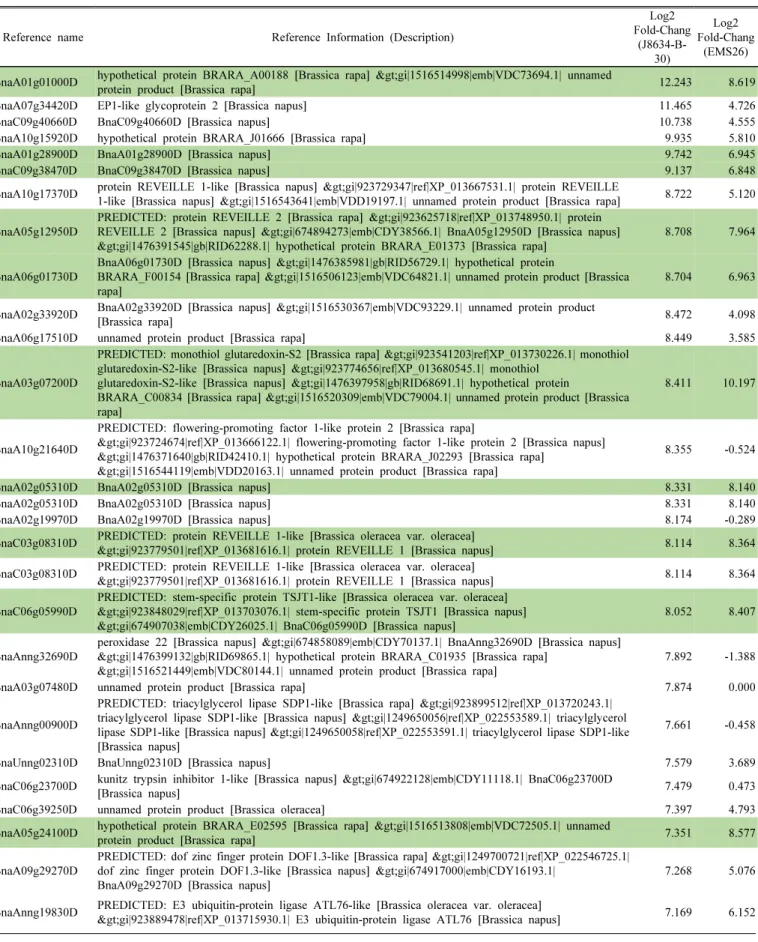
한 결과, BnaA10g21640D, BnaAnng32690D, BnaC06g23700D
같은 유전자는 ‘EMS26’ 대비 ‘J8634-B-30’ 계통에서 강하게
발현이 유도되었다(Fig. 4a). BnaA10g21640D (Flowering-
promoting factor 1-like protein 2)는 flowering- promoting
factor 관련 유전자로 Litchi chinensis에서는 저온과 ROS
(Reactive Oxygen Species) 반응과 관련된 유전자로 분류
되어 저온 처리 후 발현이 유도된다고 보고된 바 있다(Lu
et al., 2017). BnaAnng32690D와 BnaC06g23700D는 각각
peroxidase, 식물체 병원균 방어 기제에 관여하는 kunitz trypsin
inhibitor 1-like 관련 유전자로, Pu (2019) 등은 유채 0°C
이하의 저온 스트레스 반응 시 식물-병원균 상호작용 관련
유전자군의 발현이 증가했다고 보고하여 이와 관련이 있을
것으로 판단된다. 0°C 이하의 저온 스트레스 내성에 관련된
proline 합성 관련 BnaA04g22810D, BmaC04g46630D 유
전자 역시 ‘J8634-B-30’ 계통에서는 log2 fold chage 값이
각각 7.0, 6.5로 발현이 강하게 유도되었다. 또한 cytochrome
P450 관련 BnaC01g01500D 유전자는 ‘J8634-B-30’ 계통의
DEG 에서는 발현이 유도되었지만, ‘EMS26’ 계통에서는 발
현이 크게 감소하여 상반된 발현 패턴을 보였다. Cytochrome
P450 단백질은 생물적 및 비 생물적 스트레스의 방어기작
에 관여한다고 보고되었으며, 특히 수수의 전사체 분석에
서는 저온 스트레스하에 발현이 유도됐다고 보고된 바 있
다(Narusaka et al., 2004; Chopra et al., 2015).
발현이 많이 억제된 각각의 상위 100개의 DEG의 경우,
두 계통에서 공통적으로 강하게 억제되는 BnaA03g57560D,
BnaC03g38520D, BnaC07g05770D 등의 유전자들은 아직
기능이 밝혀지지 않은 것으로 나타났다(Fig. 4b). ‘EMS26’
계통의 DEG 중 ‘J8634-B-30’ 계통 대비 발현이 억제된 유
전자는 BnaC02g41180D, BnaC01g12610D, BnaC02g31910D
등이었다. BnaC02g41180D는 heavy metal-associated
iso-prenylated plant protein을 코딩하는 유전자로, 벼에서 저온
(a)
(b)
Fig. 4. Heatmap analysis of Top 100 up- and down-regulated
DEGs in two rapeseed lines. The heatmap represent
up (a) or down (b) regulated top 100 DEGs under cold
treatment in two rapeseed lines. Twenty-one DEGs were
the common up-regulated DEGs between ‘J8634-B-30’
and ‘EMS26’. Eleven DEGs were the common down-
regulated DEGs between ‘J8634-B-30’ and ‘EMS26’.
The expression levels of each gene (log2 fold change)
are indicated by different colors. Green and blue means
up- and down- regulation, respectively.
과 가뭄 스트레스 조건에서 품종에 따라 발현이 유도되어
환경 스트레스 반응에 역할을 할 것이라 보고된 바 있다(de
Abreu-Neto et al., 2013). BnaC01g12610D와 BnaC02g31910D
는 각각 gibberellin 2-beta-dioxygenase과 senescence-specific
cystein protease SAG12 유전자로 식물체 생장과 관련된
유전자였다.
저온 스트레스에 반응하는 Proline 물질대사 관련 유전자
발현 비교
식물 내 proline 축적은 생합성과 이화작용(Cataboilsm)
사이의 균형에 의해 좌우된다(Sharma et al., 2011). 식물에
서 proline은 glutamate pathway와 ornithine pathway에 의
해 생합성된다고 알려져 있다(Fig. 5a). 먼저 glutamate는 Δ’-
pyrroline-5-carboxylate synthetase (P5CS)에 의해 Δ’-pyrroline-
5-carboxylate (P5C)로 변환되고, 이후 Δ’-pyrroline-5-carboxylate
reductase (P5CR)에 의해 proline으로 생합성된다(Hu et al.,
1992; Savouré et al., 1995). Proline의 이화작용은 미토콘
드리아 안에서 일어나며, proline dehydrogenase (PDH)에
의해 P5C로 전환 된 후, P5C dehydrogenase (P5CDH)에 의
해 다시 glutamate로 변환된다(Deuschle et al., 2001; Ribarits
et al., 2007).
0°C 이하의 저온 스트레스 처리에 의한 유채 2 계통의
DEG 중 proline 생합성과 이화작용에 관여하는 유전자들의
발현 양상은 계통 간 차이를 보였다(Fig. 5b). P5CS 유전자
중 P5CSA 유전자인 BnaA04g22810D와 BnaC04g46630D
유전자들은 ’EMS26’ 계통에서는 거의 발현이 되지 않았지
만, ‘J8634-B-30’ 계통에서는 log2 fold change 값이 각각
7.0, 6.5로 나타나 강하게 발현이 유도되었다. P5CSB 유전
자인 BnaA09g35230D는 두 계통에서 유사한 발현 패턴을
보였다. 또한, ornithine를 P5C로 변환시키는 OAT 유전자
(Verbruggen & Christian, 2008; Hayat et al., 2012)에서는
두 유채 계통 모두 특이적인 발현 변화를 보이지 않았다.
Proline을 다른 세포로 운반시키는 transporter인 proline
trans-porter (ProT)와 amino acid permease 6 (AAP
6)의 발현은
두 유채 계통에서 유사하게 발현이 억제되어 세포 내 축적
을 유도한다는 것을 확인하였다. 식물체내 proline의 생합
성은 주로 P5CS 유전자에 의해 강력하게 조절된다고 보고
되고 있어(Szabados & Amould, 2010; Sharma et al., 2011;
Pu et al., 2019), 본 결과 또한 P5CSA 유전자가 강하게 발
현이 유도되어 ‘J8634-B-30’ 계통에서 proline이 축적된 것
으로 판단된다.
유채의 proline 이화작용과 관련된 유전자 중 PDH를 코
딩하는 BnaAnng37880D와 BnaC04g31100D 유전자들은
‘EMS26’ 계통에서는 발현 변화가 없었으나, ‘J8634-B-30’
계통에서는 log2 fold change 값이 각각 -7.6, -4.3으로 강력
하게 발현이 억제됨을 확인하였다(Fig. 5b). 이러한 결과들
을 종합해 볼 때, proline의 생합성을 유도하는 P5CS 유전
자가 강하게 유도되고 proline을 분해시키는 PDH 유전자
가 억제됨으로써, ‘J8634-B-30’ 계통에서 저온 처리에 의한
proline 함량이 증가한 것으로 판단된다.
(a)
(b)
Fig. 5. Proline metabolism in plants. (a) Proline metabolism step. ‘J’; ‘J8634-B-30, ‘E’; ‘EMS26’. The arrows indicate up- or
down- regulated DEGs. (Figure adapted from Hayat et al., 2012). (b) Heatmap analysis of genes involved in proline
metabolism in two rapeseed lines. P5CS (Δ’-pyrroline-5-carboxylate synthetase), P5CR (Δ’-pyrroline-5-carboxylate
reductase), PDH (Proline dehydrogenase), P5CDH (P5C dehydrogenase), OAT (Orinithine-delta-aminotransferase), ProT
(Proline transporter), and AAP
6(Amino acid permease 6). The expression levels of each gene (log2 fold change) are
(a)
(b)
(c)
(d)
Fig. 6. Heatmap analysis of the genes in ABA signal pathway (a) CE-DREB-COR signal pathway (b), cold shock domain related
protein (c) and vernalization pathway (d). PYR (Pyrabactin resistance); PYL (Pyrabactin resistance-like protein); ABA
receptor, ABI (Abscisic acid- insensitive); bZIP transcription factor, HAB (Hypersensitive to ABA); Protein phosphatase
2C, AHG (ABA-hypersensitive germination); Protein phosphatase 2C, SRK2 (SNF1-related kinase 2); Serine/threonine-
protein kinase. ICE (Inducer of CBF expression); bHLH transcription factor, DREB (Dehydration responsive element-
binding); CBF (C-repeat-binding factor), COR (Cold-regulated), VRN (Vernalization); B3 domain-containing transcripton
factor and Zinc finger protein, VIN (Vernalization insensitive); PHD finger protein, VIL (VIN3-like protein), FLC
(Flowering locus C); MADS-box protein. The expression levels of each gene (log2 fold change) are indicated by different
colors. Green and blue means up- and down- regulation, respectively.
저온 스트레스에 관여하는 생리반응 경로 관련 유전자 발
현 비교
식물의 저온 피해에 반응하는 대표적인 호르몬인 ABA 신
호 체계가 유채 두 계통의 저온 반응에 미치는 영향을 확인하
기 위해 관련 유전자를 선별하여 발현 양상을 확인하였다
(Fig. 6a). ABA 수용체로 알려진 PYR (Pyrabactin resistance)
와 PYL (Pyrabactin resistance-like protein)는 ABA가 존재할
때, SRK2 (SNF1-related kinase 2)를 억제하고 있는 PP2C
(Protein phosphatase 2C)와 결합하여 SRK2를 활성화 시킨다
(Ng et al., 2014; Sah et al., 2016). 이후 SRK2는 transcription
factor 같은 downstream 유전자들을 활성화 시켜, 이온 이
동, 유전자 발현, 대사 작용을 유도하고 이로 인해 식물체
는 비생물학적 스트레스에 내성을 갖게 된다(Hrabak et al.,
2003; Saruhashi et al., 2015). ABA 수용체인 PYL5 유전자
를 코딩하는 BnaAnng40650D는 ‘J8634-B-30’ 계통에서 log2
fold change 값이 4.2로 강하게 발현되는 것을 확인하였으
며, PYL9 유전자인 BnaA07g38130D와 BnaC06g17940D
또한 ‘J8634-B-30’ 계통에서만 발현이 강하게 유도됨을 확
인하였다(Fig. 6a). 벼에서 확인된 OsPYL9 유전자는 가뭄
과 저온에 반응하는 ABA receptor로 이 유전자를 과발현시
킨 벼에서는 저온 스트레스에 내성이 향상된다는 결과가 보
고되었다(Tian et al., 2015). PYL7인 BnaC03g31730D 유전
자는 ‘J8634-B-30’와 ‘EMS26’ 계통에서 log2 fold change
값이 각각 3.4, 5.0으로 나타나 두 계통에서 공통적으로 강
하게 발현이 유도되었으며, 본 결과와 유사하게 기존 유채
에서 저온과 0°C 이하의 저온 스트레스 처리에 의해 PYL7
유전자의 발현이 유도된다고 보고된 바 있다(Xin et al.,
2019).
유채 두 계통의 PP2C (Protein phosphatase 2C) 그룹 중
ABI (Abscisic acid-insensitive) 유전자군과 SRK2 유전자
군에서는 특이적인 발현 변화를 보이지 않았으며, SRK2J
인 BnaC04g35560D 유전자는 ‘J8634-B-30’ 계통에서 발현
이 강하게 억제되는 것을 확인하였다. SRK2J 유전자는 삼투
압 스트레스에 반응한다고 알려져 있으며, SRK2A, SRK2D,
SRK2E, SRK2I 유전자와 함께, SRK2 subclass 중 ABA에
반응하지 않다고 보고된바 있다(Boudsocq et al., 2004; Ng
et al., 2014).
식물체의 저온순화와 관련된 대표적인 신호체계에는 ICE-
CBF-COR 신호 회로가 있다. 저온 처리 초기 ICE는
transcrip-tion factor로 CBF와 함께 COR 유전자 promoter의 cis-element
에 결합하여 COR 유전자의 발현을 유도한다(Chinnusamy et
al., 2007; Ritonga & Chen, 2020). CBF는 DREB1 (dehydration-
responsive element-binding proteins)에 속하는 단백질로
식물체 내에서 건조 및 저온 스트레스하에 발현이 유도된
다고 보고되었다(Sakuma et al., 2002; Borba et al., 2018).
유채 2 계통의 DREB1를 코딩하는 유전자 중 BnaC03g
71900D, BnaA03g13620D, BnaA10g07630D는 ‘EMS26’에
서만 발현이 강하게 유도되었으며, BnaCnng49280D는 두 계
통 모두 발현이 강하게 유도되었다(Fig, 6b). BnaCnng49280D
는 이전 연구에서도 유채의 ‘HX17’, ‘HX58’ 계통에서도
강하게 발현이 유도된다고 보고되었다(Xin et al., 2019).
반면, DREB3 유전자 BnaA10g20950D, BnaC02g03940D,
BnaC09g45000D는 유채 2 계통에서 모두 발현이 억제되었
다. DREB3는 저온 스트레스에 반응한다는 결과보다 과발
현 시켰을 시 고온과 염스트레스에 내성이 보인다는 결과
가 보고된 바 있어(Niu et al., 2020), 이러한 결과가 나타났
을 것으로 판단된다.
저온에 반응하는 COR15A와 COR15B 유전자의 발현은
유채 2 계통 모두 발현에 변화를 보이지 않았지만, COR413
유전자 BnaA08g15470D와 BnaC03g61740D는 ‘J8634-B-30’
계통에서만 발현이 강하게 유도되는 결과를 보였다(Fig.
6b). COR413 유전자는 원형질막 단백질로 알려져 있으며, 건
조 및 저온 스트레스하에 식물체의 원형질막을 안정화 시
켜, 스트레스에 내성을 갖게 한다고 보고되었다(Guo et al.,
2019; Ritonga & Chen, 2020).
또한, 저온과 관련된 cold shock domain, cold responsive
protein kinase 등과 관련된 단백질들을 선별하여 유채 2 계
통에서 저온 처리에 의한 발현 변화를 비교하였다(Fig. 6c).
Cold shock protein 2를 코딩하는 BnaC07g11440D는 ‘EMS
26’에서만 발현이 억제되었으며, cold responsive protein
kinase 1인 BnaC05g12740D는 ‘J8634-B-30’에서만 발현이
유도되는 결과를 확인하였다. 두 유전자는 기능이 확실히
보고되지 않았지만, 단백질 도메인 예측 프로그램으로 저
온에 반응하는 단백질 관련 도메인을 포함하고 있다고 예
측되어, 추후 저온에서 두 유전자의 기능을 더 연구할 필요
가 있을 것이다.
마지막으로 저온에 반응하는 기작 중
춘화처리(Vernali-zation)와 관련된 유전자를 선별하여 두 유채 계통의 DEG
발현을 비교하였다(Fig. 6d). VRN (Vernalization)은 DNA
binding 단백질로 식물체의 개화에 관여하며, 밀과 같은 동
계작물이 저온에 노출될 때 발현이 유도된다고 보고되었다
(Yan et al., 2003; Amasino, 2004). PHD (Histone binding
motif) 도메인을 포함하는 VIN3 (Vernalization insensitive)
과 VIL (VIN3-like protein)은 FLC (Flowering locus C)을
억제하는 단백질로, 식물의 개화를 억제하고 있는 FLC를 억
제함으로써 개화를 유도한다고 보고되었다(Sheldon et al.,
1999; Amasino, 2004). VIN3를 코딩하는 BnaA02g08140D
와 BnaC02g11530D는 두 유채계통에서 공통적으로 발현
이 유도되었으며, FLC 유전자인 BnaA07g13680D에서는
두 계통에서 모두 발현이 감소되는 결과를 확인하였다. 이
는 저온에 노출된 두 유채 계통 모두 춘화처리에 반응하여
봄철 개화가 유도되고 있다는 점을 나타낸다.
저온 스트레스에 반응하는 유전자 발현 확인
저온 스트레스에 특이적으로 반응하는 주요 DEG들의 발
현을 확인하고, 저온 순화 이후 0°C 이하의 저온 스트레스
처리에도 어떠한 발현 변화를 보이는 지 확인하기 위해
RT-PCR을 실시하였다. RT-PCR의 결과는 전사체 분석 결
과와 동일하였으며, 이후 0°C 이하의 저온 스트레스 처리
에서 몇몇 유전자에서 유채 두 계통간의 발현 변화를 보였
다(Fig. 7). DREB1 단백질을 코딩하는 BnaC03g71900D와
BnaA10g07630D, COR413 단백질을 코딩하는 BnaA08g
15470D 유전자의 발현의 경우, 초기 저온순화 과정에서는
‘J8634-B-30’ 계통에서 강하게 발현하였으나, 이후 -9°C 처
리 시 발현이 감소하였다. 반면, ‘EMS26’ 계통에서는 초기
저온순화 과정에서는 발현 변화가 없었으나, -5°C와 -9°C
의 스트레스 조건하에서 강하게 발현되는 변화를 확인하였
다. 이는 초기 저온순화 시 ‘J8634-B-30’ 계통에서 proline
함량이 높게 확인되고, 초기 생존율이 높은 결과와 일치하
며, ‘EMS26’ 계통에서는 후기 생존율이 회복되는 결과와
유사한 것으로 판단된다.
본 연구 결과, 0°C 이하의 저온 스트레스 처리로 인해 유
채 두 계통의 생리적 지표 중 proline 함량 및 초기 내동성
에서 차이를 확인하였다. 이러한 변화와 관련된 기작을 확
인하기 위해 두 유채 계통을 대상으로 전사체 비교 분석을
실시하였으며, 저온스트레스에 반응하는 proline 대사, ABA
호르몬 신호 경로, ICE-CBF-COR 신호 경로, 춘화처리와
관련된 유전자들의 발현을 비교하여 유채 두 계통에서의
발현 차이점을 구명하였다. 이러한 특이적인 발현 양상을
보인 유전자들은 추후 유전자 기능 구명 등을 통해 0°C 이
하의 저온 스트레스 피해에 적응할 수 있는 유채 품종 육성
및 관련 기작 연구에 기여할 수 있을 것으로 판단된다.
적 요
본 연구에서는 유채 두 계통에 0°C 이하의 저온 스트레
스를 처리하고 이에 따른 proline 함량 및 생존율 변화를 확
인하고 이에 따른 저온에서의 유전자 발현 변화를 비교 분
석하기 위해 수행되었으며, 결과는 다음과 같다.
1. 0°C 이하의 저온 스트레스 처리 전 저온 순화 후 유채
‘J8634-B-30’ 계통에서 proline 함량이 2.02 mM/g FW
로 증가하여 처리 전보다 8.7배 증가하였으며, ‘EMS26’
계통에서는 0°C 이하의 저온 스트레스 처리로 인한
proline 함량 변화를 보이지 않았다.
2. 저온 순화 전후 유채 두 계통의 전사체를 분석한 결과,
‘J8634-B-30’ 계통의 DEG는 발현이 유도된 DEG가 2,784
개로, 발현이 억제된 DEG 2,299개보다 많았으며, ‘EMS
26’ 계통에서는 발현이 유도된 DEG가 2,199개로 발현이
억제된 DEG 3,632개로 적었다.
3. 저온 스트레스 처리에 의한 유채 두 계통의 상위 100개
DEG를 분석한 결과, ‘J8634-B-30’ 계통에서는 flowering-
Fig. 7. RT-PCR validation analysis of rapeseed two lines under
cold stress treatment on DEGs. Con (Control) indicates
15°C. Bactin was used as an internal control. Detailed
information is listed in Table S1.
promoting factor (BnaA10g21640D) 유전자가 강하게
발현되었으며, 특히 proline 생합성 관련 유전자의 발현
이 강하게 유도되었다. ‘EMS26’ 계통의 DEG에서는 식
물체 생장과 관련된 유전자가 발현이 억제되었다.
4. ‘J8634-B-30’ 계통에서는 proline 생합성 관련 P5CSA
(BnaA04g22810D, BnaC04g46630D) 유전자의 발현이
유도되었으며, proline 이화작용 관련 PDH (BnaAnng
37880D, BnaC04g31100D) 유전자 발현이 억제되어, 저
온 처리에 의한 proline 함량 이 증가한 것으로 판단된다.
5. 저온 반응 관련 생리반응 경로 유전자 중 ABA 호르몬 수
용체 PYL5 (BnaAnng40650D), PYL9 (BnaA07g38130D,
BnaC06g17940D)는 ‘J8634-B-30’ 계통에서만 발현이 유
도되었다. 또한, ICE-CBF-COR 신호 회로 중 원형질막
안정화에 관여하는 COR413 유전자(BnaA08g15470D,
BnaC03g61740D)에서도 ‘J8634-B-30’ 계통에서만 발현
이 유도되었다.
6. 이러한 저온 스트레스 반응과 관련된 유전자 발현 차이
는 초기 저온 처리 후 두 유채 계통의 스트레스 반응에
관여했을 것으로 판단되며, 특이적인 발현 양상을 보인
유전자에 대해 향후 추가적인 기능을 분석하여 내동성
유채 품종 개발에 활용할 수 있을 것으로 판단된다.
사 사
본 연구는 농촌진흥청 작물시험연구사업(과제번호: PJ01
433202)의 지원에 의해 수행되었다.
인용문헌(REFERENCES)
Amasino, R. 2004. Vernalization, competence, and the epigenetic
memory of winter. Plant Cell. 16 : 2553-2559.
Anders, S. and W. Huber. 2010. Differential expression analysis
for sequence count data. Nature Precedings, 1-1.
Anders, S., P. T. Pyl, and W. Huber. 2015. HTSeq—a Python
framework to work with high-throughput sequencing data.
Bioinformatics 31 : 166-169.
Bolger, A. M., M. Lohse, and B. Usadel. 2014. Trimmomatic: a
flexible trimmer for Illumina sequence data. Bioinformatics.
30 : 2114-2120.
Borba, A. R., T. S. Serra, A. Górska, P. Gouveia, A. M. Cordeiro,
I. Reyna-Llorens, J. Kneřová, P. M. Barros, I. A. Abreu, and
M. M. Oliveira. 2018. Synergistic binding of bHLH transcription
factors to the promoter of the maize NADP-ME gene used in
C
4photosynthesis is based on an ancient code found in the
ancestral C
3state. Mol Biol Evol. 35 : 1690-1705.
Boudsocq, M., H. Barbier-Brygoo, and C. Laurière. 2004.
Iden-tification of nine sucrose nonfermenting 1-related protein
kinases 2 activated by hyperosmotic and saline stresses in
Arabidopsis thaliana. J Biol Chem. 279 : 41758-41766.
Burbulis, N., V. Jonytiene, R. Kupriene, and A. Blinstrubiene.
2011. Changes in proline and soluble sugars content during
cold acclimation of winter rapeseed shoots in vitro. J Food
Agric Environ. 9 : 371-374.
Chalhoub, B., F. Denoeud, S. Liu, I. A. Parkin, H. Tang, X. Wang,
J. Chiquet, H. Belcram, C. Tong, and B. Samans. 2014. Early
allopolyploid evolution in the post-Neolithic Brassica napus
oilseed genome. Science 345 : 950-953.
Chen, H. H., P. H. Li, and M. L. Brenner. 1983. Involvement of
abscisic acid in potato cold acclimation. Plant Physiol. 71 :
362-365.
Chinnusamy, V., J. Zhu, and J. K. Zhu. 2007. Cold stress regulation
of gene expression in plants. Trends Plant Sci. 12 : 444-451.
Chopra, R., G. Burow, C. Hayes, Y. Emendack, Z. Xin, and J.
Burke. 2015. Transcriptome profiling and validation of gene
based single nucleotide polymorphisms (SNPs) in sorghum
genotypes with contrasting responses to cold stress. BMC
Genomics. 16 : 1-11.
de Abreu‐Neto, J. B., A. C. Turchetto‐Zolet, L. F. V. de Oliveira,
M. H. Bodanese Zanettini, and M. Margis-Pinheiro. 2013.
Heavy metal‐associated isoprenylated plant protein (HIPP):
characterization of a family of proteins exclusive to plants.
FEBS J. 280 : 1604-1616.
Deuschle, K., D. Funck, H. Hellmann, K. Däschner, S. Binder,
and W. B. Frommer. 2001. A nuclear gene encoding
mito-chondrial Δ1-pyrroline‐5‐carboxylate dehydrogenase and its
potential role in protection from proline toxicity. Plant J. 27 :
345-356.
Guo, X., L. Zhang, G. Dong, Z. Xu, G. Li, N. Liu, A. Wang, and
J. Zhu. 2019. A novel cold-regulated protein isolated from
Saussurea involucrata confers cold and drought tolerance in
transgenic tobacco (Nicotiana tabacum). Plant Sci. 289 : 110246.
Guy, C.L. 1990. Cold acclimation and freezing stress tolerance:
role of protein metabolism. Annu Rev Plant Biol. 41 : 187-
223.
Hayat, S., Q. Hayat, M. N. Alyemeni, A. S. Wani, J. Pichtel, and
A. Ahmad. 2012. Role of proline under changing environments:
a review. Plant Signal Behav. 7 : 1456-1466.
Hrabak, E. M., C. W. Chan, M. Gribskov, J. F. Harper, J. H. Choi,
N. Halford, J. Kudla, S. Luan, H. G. Nimmo, and M. R.
Sussman. 2003. The Arabidopsis CDPK-SnRK superfamily
of protein kinases. Plant Physiol. 132 : 666-680.
Hu, C., A. J. Delauney, and D. Verma. 1992. A bifunctional enzyme
(delta 1-pyrroline-5-carboxylate synthetase) catalyzes the first
two steps in proline biosynthesis in plants. PNAS. 89 : 9354-
9358.
Jeong, Y. J., Y. H. Choy, H. J. Joo, J. H. Hwang, Y. J. Byun, Y. M.
Lee, J. S. Lee, Y. S. Jang, and D. H. Lee. 2012. Identification
and analysis of cold stress-inducible genes in Korean rapeseed
varieties. J Plant Biol. 55 : 498-512.
Kim, B. M. 2015. Studied on physiological properties and seletion
of low-temperature resistant plant of Brassica napus L.
Un-published master’s dissertation. Mokpo National University.
Mokpo.
Kim, D., B. Langmead, and S. L. Salzberg. 2015. HISAT: a fast
spliced aligner with low memory requirements. Nat Methods.
12 : 357-360.
Koster, K. L. and D. V. Lynch. 1992. Solute accumulation and
compartmentation during the cold acclimation of Puma rye.
Plant Physiol. 98 : 108-113.
Lee, K. R., S. I. Sohn, J. H. Jung, S. H. Kim, K. H. Roh, J. B. Kim,
M. C. Suh, and Kim, H. U. 2013. Functional analysis and
tissue-differential expression of four FAD2 genes in
amphidi-ploid Brassica napus derived from Brassica rapa and Brassica
oleracea. Gene. 531 : 253-262.
Levitt, J., 1980. Responses of Plants to Environmental Stresses:
Chilling, Freezing and High Temperature Stresses, 2nd Edn,
Vol. 1. NY: Academic Press. New York. 0160-9327.
Lu, X., J. Li, H. Chen, J. Hu, P. Liu, and B. Zhou. 2017. RNA-seq
analysis of apical meristem reveals integrative regulatory
network of ROS and chilling potentially related to flowering
in Litchi chinensis. Sci Rep. 7 : 1-13.
Lynch, D. V. and P. L. Steponkus. 1987. Plasma membrane lipid
alterations associated with cold acclimation of winter rye
seedlings (Secale cereale L. cv Puma). Plant Physiol. 83 :
761-767.
McClinchey, S. L., and L. S. Kott. 2008. Production of mutants
with high cold tolerance in spring canola (Brassica napus).
Euphytica. 162 : 51-67.
Narusaka, M., M. Seki, T. Umezawa, J. Ishida, M. Nakajima, A.
Enju, and K. Shinozaki. 2004. Crosstalk in the responses to
abiotic and biotic stresses in Arabidopsis : analysis of gene
expression in cytochrome P450 gene superfamily by cDNA
microarray. Plant Mol. Biol. 55 : 327-342.
Ng, L. M., K. Melcher, B. T. Teh, and H. E. Xu. 2014. Abscisic
acid perception and signaling : structural mechanisms and
applications. Acta Pharmacol Sin. 35 : 567-584.
Niu, X., T. Luo, H. Zhao, Y. Su, W. Ji, and H. Li. 2020.
Identifi-cation of wheat DREB genes and functional characterization
of TaDREB3 in response to abiotic stresses. Gene. 144514.
Pu, Y., L. Liu, J. Wu, Y. Zhao, J. Bai, L. Ma, J. Yue, J. Jin, Z. Niu,
and Y. Fang. 2019. Transcriptome profile analysis of winter
rapeseed (Brassica napus L.) in response to freezing stress,
reveal potentially connected events to freezing stress. Int J
Mol Sci. 20 : 2771.
Ribarits, A., A. Abdullaev, A. Tashpulatov, A. Richter, E. Heberle-
Bors, and A. Touraev. 2007. Two tobacco proline
dehydro-genases are differentially regulated and play a role in early
plant development. Planta. 225 : 1313-1324.
Ritonga, F. N. and S. Chen. 2020. Physiological and molecular
mechanism involved in cold stress tolerance in plants. Plants.
9 : 560.
Ruelland, E., M. N. Vaultier, A. Zachowski, and V. Hurry. 2009.
Cold signalling and cold acclimation in plants. Adv Bot Res.
49 : 35-150.
Sah, S. K., K. R. Reddy, and J. Li. 2016. Abscisic acid and abiotic
stress tolerance in crop plants. Front Plant Sci. 7 : 571.
Sakai, A. and W. Larcher. 2012. Frost survival of plants: responses
and adaptation to freezing stress. Vol. 62. Springer Science &
Business Media.
Sakuma, Y., Q. Liu, J. G. Dubouzet, H. Abe, K. Shinozaki, and K.
Yamaguchi-Shinozaki. 2002. DNA-binding specificity of the
ERF/AP2 domain of Arabidopsis DREBs, transcription factors
involved in dehydration-and cold-inducible gene expression.
Biochem Biophys Res Commun. 290 : 998-1009.
Sanghera, G. S., S. H. Wani, W. Hussain, and N. Singh. 2011.
Engineering cold stress tolerance in crop plants. Curr Genomics.
12 : 30.
Saruhashi, M., T. K. Ghosh, K. Arai, Y. Ishizaki, K. Hagiwara, K.
Komatsu, Y. Shiwa, K. Izumikawa, H. Yoshikawa, and T.
Umezawa. 2015. Plant Raf-like kinase integrates abscisic acid
and hyperosmotic stress signaling upstream of SNF1-related
protein kinase2. PNAS. 112 : 6388-6396.
Savouré, A., S. Jaoua, X. J. Hua, W. Ardiles, M. Van Montagu,
and N. Verbruggen. 1995. Isolation, characterization, and
chromosomal location of a gene encoding the Δ 1-pyrroline-
5-carboxylate synthetase in Arabidopsis thaliana. FEBS letters.
372 : 13-19.
Schroeder, J. I., J. M. Kwak, and G. J. Allen. 2001. Guard cell
abscisic acid signalling and engineering drought hardiness in
plants. Nature. 410 : 327-330.
Sharma, S., J. G. Villamor, and P. E. Verslues. 2011. Essential
role of tissue-specific proline synthesis and catabolism in
growth and redox balance at low water potential. Plant Physiol.
157 : 292-304.
Sheldon, C. C., J. E. Burn, P. P. Perez, J. Metzger, J. A. Edwards,
W. J. Peacock, and E. S. Dennis. 1999. The FLF MADS box
gene: a repressor of flowering in Arabidopsis regulated by
vernalization and methylation. Plant Cell. 11 : 445-458.
Shi, Y., Y. Ding, and S. Yang. 2018. Molecular regulation of CBF
signaling in cold acclimation. Trends Plant Sci. 23 : 623-637.
Shinozaki, K. and K. Yamaguchi-Shinozaki. 2000. Molecular
responses to dehydration and low temperature: differences
and cross-talk between two stress signaling pathways. Curr
Opin. Plant Biol. 3 : 217-223.
Szabados, L. and A. Savoure. 2010. Proline : a multifunctional
amino acid. Trends Plant Sci. 15 : 89-97.
Tao, D. L., G. Öquist, and G. Wingsle. 1998. Active oxygen
scavengers during cold acclimation of Scots pine seedlings in
relation to freezing tolerance. Cryobiology. 37 : 38-45.
Thomashow, M. F. 1999. Plant cold acclimation: freezing tolerance
genes and regulatory mechanisms. Annu Rev Plant Biol. 50 :
571-599.
Tian, X., Z. Wang, X. Li, T. Lv, H. Liu, L. Wang, H. Niu, and Q.
Bu. 2015. Characterization and functional analysis of pyrabactin
resistance-like abscisic acid receptor family in rice. Rice. 8 :
1-13.
Trischuk, R. G., B. S. Schiling, M. Wisniewski, and L. V. Gusta.
2006. Freezing stress: systems biology to study cold tolerance,
Physiology and Molecular Biology of Stress Tolerance in
Plants. Springer, Dordrecht. 131-155.
Verbruggen, N. and C. Hermans. 2008. Proline accumulation in
plants: a review. Amino acids. 35 : 753-759.
Waalen, W. M., K. K. Tanino, J. E. Olsen, R. Eltun, O. A. Rognli,
and L. V. Gusta. 2011. Freezing tolerance of winter canola
cultivars is best revealed by a prolonged freeze test. Crop Sci.
51 : 1988-1996.
Xin, H., N. Xianchao, X. Pan, L. Wei, Y. Min, K. Yu, Q. Lunwen,
and H. Wei. 2019. Comparative transcriptome analyses
revealed conserved and novel responses to cold and freezing
stress in Brassica napus L. G3. 2723-2737.
Xin, Z. and J. Browse. 2000. Cold comfort farm : the acclimation
of plants to freezing temperatures. Plant Cell Environ. 23 :
893-902.
Yan, L., A. Loukoianov, G. Tranquilli, M. Helguera, T. Fahima,
and J. Dubcovsky. 2003. Positional cloning of the wheat
vernalization gene VRN1. PNAS. 100 : 6263-6268.
Fig. S1. The experimental diagram of freezing stress treatment of two rapeseed line. The arrows indicate the timing of sampling
for the experiment.
Table S1. RT-PCR primers designed for DEGs induced and depressed by cold treatment in rapeseed two lines.
Reference name Tentative annotation Primers
Top 100 UP DEGs
BnaA10g21640D Flowering-promoting factor 1-like protein 2 F : ATGTCAGGCGTTTGGGTGTTC
R : TCAGTGGAAATCACGGACG
BnaAnng32690D Peroxidase 22 F : CCCAGATTGAGAGAGCTTGC
R : TTGCTGCTGTATTGCTCCAC
BnaC06g23700D Kuniz trypsin injibitor 1-like F : TCGCTCTTGCCTTAACCCTA
R : CAAACGCCTAACACCGATTT
BnaC01g01500D Cytochrome P450 81F1-like F : AACTGACGGCCAAATACGTC
R : CAAGTAATCCGCCCTGTGAT
Top 100 Down DEGs
BnaC02g41180D Heavy metal-associated isoprenylated plant protein 9 F : TCCGCCCTTCATACTCTACG
R : CAAGCGTTAGGGTTCTCGTC
BnaC01g12610D Gibberellin 2-beta-dioxygenase 8 F : CAGACAGCAATGTCCTCGAA
R : ACTGTCCGTGTGTGGCATTA
BnaC02g31910D Senescence-specific cysteine protease SAG12 F : TCATGCAAAAGAAGCACGAC
R : CCAACAACATCCACAACTGC
Proline metabolism
BnaA04g22810D P5CSA, Delta-1-pyrroline -5-carboxylate synthase A F : ACCAAGGGCAAGTGCAATAC
R : TCCCATTGTCTCCATTCACA
BnaC04g46630D P5CSA, Delta-1-pyrroline -5-carboxylate synthase A F : GCGCATAGTTTCTGATGCAA
R : CCACAACTTGTCCGTTTCCT
BnaA09g35230D P5CSB, Delta-1-pyrroline -5-carboxylate synthase B F : GCTCAAATGCTGGTGACTGA
R : ATGTGCATCTTGGTGGAACA
BnaAnng37880D PDH, Proline dehydrogenase mitochondrial F : CAGCTCACGAACGCATTAAA
R : AACCAGCCCTTTTCAATCCT
BnaC04g31100D PDH, Proline dehydrogenase 2 F : TCCCGGAGATTCTTTCCTTT
R : CGTCTTCACCTGCACAGAAA
ABA signal pathway
BnaAnng40650D PYL5, Abscisic acid receptor PYL5 isoform X1 F : ATGAGATCAGCGGTGCAGATCC
R : TTATTGCCGGTTGGTACTTC
BnaA07g38130D PYL9, Abscisic acid receptor PYL9 F : GGTTTGGTCTCTGGTGAGGA
R : TCTTTGGTGTTCCCTTCTGG
BnaC03g31730D PYL7, Abscisic acid receptor PYL7-like F : CCCTGTTCATCTCGTTTGGT
R : ATGTCCTGAGCAGCCAATCT
ICE-DREB-COR signal pathway and vernalization pathway
BnaC03g71900D DREB1, Dehydration-responsive element-binding 1B-like F : GAGCTGTCCGAAGAAACCTG
R : GGCATCTCCGTCAAAGTCAT
BnaA10g07630D DREB1, Dehydration-responsive element-binding protein 1D F : GAGCTGTCCGAAGAAACGAG
R : GGTTTCACTGCCTCAACCAT
BnaA08g15470D COR413, Cold-regulated 413 plasma membrane protein 4 F : GTTGGGGCAGTGATTTGTCT
R : CCAAACTGGGTAAACGAGGA
BnaA02g08140D VIN3 F : AGTGAAGGTGGCGAAAGAGA
R : AGAGCTTGAGGCAGATTCCA
BnaA07g13680D FLC F : CCGTTCTGTCTCCTCTCCTG
R : ATTCTCTGCTGCACCCATCT
Internal control
Bactin
Ta bl e S2 . In for mat ion o f GO en rich me nt of DEGs in du ced by cold treat m en t in ‘J8634 -B-30 ’. GO Cate gor y GO ID GO Nam e Fol d-ch an ge (%T est / % Ref) FDR P-V alu e -L O G (P-Val ue) Nr Test Nr Re ference Non Anno t Te st Non Anno t Referen ce Tes tS eq Seq uen ces Re fSet Se que nces Te st (%) Ref (% ) BIO LOGIC A L_ PROCES S GO:0070407 oxi dat ion - depen de nt protei n catabolic proces s 57.99 8.90E-04 1.75E-05 4. 76E+00 4 2 1909 55463 GSBRN A2T 00054450001;G SBRNA 2T 00147732001 ;GSBRN A2 T00133 726001;G SBRNA 2T 00005991001 G SBRNA 2T 00133727001 ;GSBRN A2 T00120490001 0.21 0. 00 BIO LOGIC A L_ PROCES S GO:1901141 re gul at io n of lignin bi os ynt het ic proc ess 57.99 0.058303054 0.003259034 2. 49E+00 2 1 1911 55464 GSBRN A2T 00069390001;G SBRNA 2T 00072620001 G SBRNA 2T 00152142001 0.10 0. 00 BIO LOGIC A L_ PROCES S GO:0090069 re gul at io n of riboso m e bio gen esi s 57.99 0.058303054 0.003259034 2. 49E+00 2 1 1911 55464 GSBRN A2T 00104673001;G SBRNA 2T 00140412001 G SBRNA 2T 00047238001 0.10 0. 00 BIO LOGIC A L_ PROCES S GO:0071428 rR NA - con tai ni ng ribon ucleoprotei n co m plex ex port fr om nuc le us 57.99 0.058303054 0.003259034 2. 49E+00 2 1 1911 55464 GSBRN A2T 00035507001;G SBRNA 2T 00090475001 G SBRNA 2T 00094092001 0.10 0. 00 BIO LOGIC A L_ PROCES S GO:0000055 riboso m al lar ge sub uni t exp or t fr om nuc le us 57.99 0.058303054 0.003259034 2. 49E+00 2 1 1911 55464 GSBRN A2T 00035507001;G SBRNA 2T 00090475001 G SBRNA 2T 00094092001 0.10 0. 00 BIO LOGIC A L_ PROCES S GO:0055129 L-proli ne bi os ynt het ic proc ess 30.36 8.92E-06 5.62E-08 7. 25E+00 7 9 1404 55060 GSBRN A2T 00072614001;G SBRNA 2T 00012577001 ;GSBRN A2 T00116 809001;G SBRNA 2T 00069393001 ;GSBRN A2 T00124740001;GSBRN A2 T00123351001 ;GSBRN A2 T00155189001 G SBRNA 2T 00048188001 ;GSBRN A2 T00054698001;GSBRN A2T 00126 340001 ;GSBRN A2 T00135564001;GSBRN A2T 00128580001;G SBRNA 2 T00031643001;GSBRN A2T 00058037001;G SBRNA 2T 00061209001 ;GS BRN A2T 00034359001 0.50 0. 02 BIO LOGIC A L_ PROCES S GO:0006561 prol in e bio sy nt hetic proces s 12.49 1.35E-04 1.18E-06 5. 93E+00 8 25 1403 55044 GSBRN A2T 00072614001;G SBRNA 2T 00012577001 ;GSBRN A2 T00082 899001;G SBRNA 2T 00116809001 ;GSBRN A2 T00069393001;GSBRN A2 T00124740001 ;GSBRN A2 T00123351001;GSBRN A2T 00155189001 G SBRNA 2T 00054698001 ;GSBRN A2 T00126340001;GSBRN A2T 00114 224001 ;GSBRN A2 T00008252001;GSBRN A2T 00105795001;G SBRNA 2 T00031643001;GSBRN A2T 00001624001;G SBRNA 2T 00059225001 ;GS BRN A2T 00058037001;G SBRNA 2T 00034359001 ;GSBRN A2 T00025136 001 ;GSBRN A2 T00112701001;GSBRN A2T 00017092001;G SBRNA 2T 00 008196001 ;GSBRN A2 T00009793001;GSBRN A2T 00048188001;G SBR N A2T 00135564001;G SBRNA 2T 00061209001 ;GSBRN A2 T0014296400 1;GSBRN A2 T00037219001;GSBRN A2T 00037220001;G SBRNA 2T 0000 9027001 ;GSBRN A2 T00128580001;GSBRN A2T 00146239001;G SBRNA 2T 00030525001 0.57 0. 05 BIO LOGIC A L_ PROCES S GO:0006560 prol in e m eta bo lic proces s 2.88 0.074645786 0.002403154 2. 62E+00 11 149 1400 54920 GSBRN A2T 00072614001;G SBRNA 2T 00012577001 ;GSBRN A2 T00082 899001;G SBRNA 2T 00116809001 ;GSBRN A2 T00069393001;GSBRN A2 T00153666001 ;GSBRN A2 T00124740001;GSBRN A2T 00123351001;G S BRN A2 T00155189001;GSBRN A2T 00032666001;G SBRNA 2T 00112018 001 G SBRNA 2T 00012960001 ;GSBRN A2 T00141139001;GSBRN A2T 00095 662001 ;GSBRN A2 T00117861001;GSBRN A2T 00008252001;G SBRNA 2 T00130859001;GSBRN A2T 00060796001;G SBRNA 2T 00112041001 ;GS BRN A2T 00127717001;G SBRNA 2T 00088172001 ;GSBRN A2 T00154466 001 ;GSBRN A2 T00090776001;GSBRN A2T 00048994001;G SBRNA 2T 00 140751001 ;GSBRN A2 T00031643001;GSBRN A2T 00059225001;G SBR N A2T 00058037001;G SBRNA 2T 00034359001 ;GSBRN A2 T0011432800 1;GSBRN A2 T00050558001;GSBRN A2T 00061209001;G SBRNA 2T 0001 3826001 ;GSBRN A2 T00135480001;GSBRN A2T 00153644001;G SBRNA 2T 00074513001;G SBRNA 2T 00122620001 ;GSBRN A2 T00009027001;G SBRN A2T 00128580001;G SBRNA 2T 00044870001 ;GSBRN A2 T0005374 1001 ;GSBRN A2 T00027811001;GSBRN A2T 00143588001;G SBRNA 2T 0 0126340001 ;GSBRN A2 T00114224001;GSBRN A2T 00022314001;G SBR N A2T 00095093001;G SBRNA 2T 00039314001 ;GSBRN A2 T0013585100 1;GSBRN A2 T00086894001;GSBRN A2T 00062619001;G SBRNA 2T 0015 2027001 ;GSBRN A2 T00036879001;GSBRN A2T 00044877001;G SBRNA 2T 00140522001;G SBRNA 2T 00049468001 ;GSBRN A2 T00000987001;G SBRN A2T 00063557001;G SBRNA 2T 00126668001 ;GSBRN A2 T0012707 8001 ;GSBRN A2 T00008196001;GSBRN A2T 00009793001;G SBRNA 2T 0 0095657001 ;GSBRN A2 T00011769001;GSBRN A2T 00047491001;G SBR N A2T 00135564001;G SBRNA 2T 00077768001 ;GSBRN A2 T0002332500 1;GSBRN A2 T00102439001;GSBRN A2T 00057197001;G SBRNA 2T 0004 5277001 ;GSBRN A2 T00083847001;GSBRN A2T 00025165001;G SBRNA 2T 00094219001;G SBRNA 2T 00143147001 ;GSBRN A2 T00104042001;G SBRN A2T 00037219001;G SBRNA 2T 00028951001 ;GSBRN A2 T0014623 9001 ;GSBRN A2 T00086237001;GSBRN A2T 00030525001;G SBRNA 2T 0 0136128001 ;GSBRN A2 T00068628001;GSBRN A2T 00143695001;G SBR N A2T 00121061001;G SBRNA 2T 00038640001 ;GSBRN A2 T0014957400 1;GSBRN A2 T00129814001;GSBRN A2T 00086399001;G SBRNA 2T 0014 0557001 ;GSBRN A2 T00099831001;GSBRN A2T 00121350001;G SBRNA 2T 00078912001;G SBRNA 2T 00051624001 ;GSBRN A2 T00095675001;G SBRN A2T 00099078001;G SBRNA 2T 00012185001 ;GSBRN A2 T0000162 4001 ;GSBRN A2 T00025136001;GSBRN A2T 00153113001;G SBRNA 2T 0 0112701001 ;GSBRN A2 T00084114001;GSBRN A2T 00078411001;G SBR N A2T 00017092001;G SBRNA 2T 00048188001 ;GSBRN A2 T0006763000 1;GSBRN A2 T00105056001;GSBRN A2T 00036370001;G SBRNA 2T 0014 1763001 ;GSBRN A2 T00154299001;GSBRN A2T 00037220001;G SBRNA 2T 00136055001;G SBRNA 2T 00099384001 ;GSBRN A2 T00110274001;G SBRN A2T 00088173001;G SBRNA 2T 00054195001 ;GSBRN A2 T0012300 1001 ;GSBRN A2 T00051069001;GSBRN A2T 00017538001;G SBRNA 2T 0 0054698001 ;GSBRN A2 T00007601001;GSBRN A2T 00063737001;G SBR N A2T 00130857001;G SBRNA 2T 00056355001 ;GSBRN A2 T0001384600 1;GSBRN A2 T00053473001;GSBRN A2T 00108512001;G SBRNA 2T 0012 1349001 ;GSBRN A2 T00093897001;GSBRN A2T 00075292001;G SBRNA 2T 00028950001;G SBRNA 2T 00105795001 ;GSBRN A2 T00154468001;G SBRN A2T 00081792001;G SBRNA 2T 00130526001 ;GSBRN A2 T0003528 3001 ;GSBRN A2 T00015956001;GSBRN A2T 00065940001;G SBRNA 2T 0 0014571001 ;GSBRN A2 T00140204001;GSBRN A2T 00130797001;G SBR 0.78 0. 27
Ta bl e S2 . In for mat ion o f GO en rich me nt of DEGs in du ced by cold treat m en t in ‘J8634 -B-30 ’ (C on tin ued). GO Cate gor y GO ID GO Nam e Fol d-ch an ge (%T est / % Ref) FDR P-V alu e -L O G (P-Val ue) Nr Test Nr Re ference Non Anno t Te st Non Anno t Referen ce Tes tS eq Seq uen ces Re fSet Se que nces Te st (%) Ref (% ) N A2T 00044882001;G SBRNA 2T 00116060001 ;GSBRN A2 T0002542200 1;GSBRN A2 T00079160001;GSBRN A2T 00076263001;G SBRNA 2T 0001 1329001 ;GSBRN A2 T00142964001;GSBRN A2T 00088529001;G SBRNA 2T 00007872001;G SBRNA 2T 00068154001 ;GSBRN A2 T00099079001;G SBRN A2T 00130118001;G SBRNA 2T 00035269001 ;GSBRN A2 T0013616 6001 ;GSBRN A2 T00107098001;GSBRN A2T 00019541001;G SBRNA 2T 0 0081082001 ;GSBRN A2 T00100028001;GSBRN A2T 00000932001 CELL UL A R_C OM PONEN T GO:0097361 CI A co mp lex 57.99 0.058303054 0.003259034 2. 49E+00 2 1 1911 55464 GSBRN A2T 00127215001;G SBRNA 2T 00049582001 G SBRNA 2T 00073786001 0. 10 0. 00 CELL UL A R_C OM PONEN T GO:0061927 TOC-TIC super comp le x I 43.49 0.009894981 3.52E-04 3. 45E+00 3 2 1910 55463 GSBRN A2T 00107247001;G SBRNA 2T 00144123001 ;GSBRN A2 T00144 124001 G SBRNA 2T 00096595001 ;GSBRN A2 T00140436001 0.16 0. 00 CELL UL A R_C OM PONEN T GO:0071541 euk aryo tic tra ns latio n ini tiatio n fa ct or 3 comp lex , eIF 3m 43.49 0.009894981 3.52E-04 3. 45E+00 3 2 1910 55463 GSBRN A2T 00029637001;G SBRNA 2T 00076282001 ;GSBRN A2 T00153 634001 G SBRNA 2T 00057231001 ;GSBRN A2 T00140463001 0.16 0. 00 CELL UL A R_C OM PONEN T GO:0031354 int rin si c comp one nt of plast id outer m embra ne 28.99 0.016599521 6.86E-04 3. 16E+00 3 3 1910 55462 GSBRN A2T 00107247001;G SBRNA 2T 00144123001 ;GSBRN A2 T00144 124001 G SBRNA 2T 00041825001 ;GSBRN A2 T00110778001;GSBRN A2T 00140 436001 0.16 0. 01 CELL UL A R_C OM PONEN T GO:0031355 in tegral co mpo ne nt of plast id outer m embra ne 28.99 0.016599521 6.86E-04 3. 16E+00 3 3 1910 55462 GSBRN A2T 00107247001;G SBRNA 2T 00144123001 ;GSBRN A2 T00144 124001 G SBRNA 2T 00041825001 ;GSBRN A2 T00110778001;GSBRN A2T 00140 436001 0.16 0. 01 MOLECU LA R_ FUNCT ION GO:0003922 G M P synt ha se (g lut amin e-hyd ro ly z ing ) ac tiv ity 57.99 0.058303054 0.003259034 2. 49E+00 2 1 1911 55464 GSBRN A2T 00000003001;G SBRNA 2T 00081839001 G SBRNA 2T 00113476001 0.10 0. 00 MOLECU LA R_ FUNCT ION GO:0009927 his tid in e ph os pho tra nsf er kin as e ac tiv ity 57.99 0.058303054 0.003259034 2. 49E+00 2 1 1911 55464 GSBRN A2T 00063796001;G SBRNA 2T 00013120001 G SBRNA 2T 00006623001 0.10 0. 00 MOLECU LA R_ FUNCT ION GO:0070626 (S)- 2- (5-am in o- 1- (5-pho sph o- D-ribo sy l)i m idazole- 4-carbo xa mi do )su cc ina te A MP-ly ase (fuma ra te-f orming ) acti vit y 43.49 0.009894981 3.52E-04 3. 45E+00 3 2 1910 55463 GSBRN A2T 00017154001;G SBRNA 2T 00126945001 ;GSBRN A2 T00112 573001 G SBRNA 2T 00146820001 ;GSBRN A2 T00017349001 0.16 0. 00 MOLECU LA R_ FUNCT ION GO:0004018 N6 -(1,2-dicarbox yet hy l)AMP A MP-ly ase (fuma ra te-f orming ) acti vit y 38.66 0.001847934 3.98E-05 4. 40E+00 4 3 1909 55462 GSBRN A2T 00131331001;G SBRNA 2T 00126945001 ;GSBRN A2 T00017 154001;G SBRNA 2T 00112573001 G SBRNA 2T 00146820001 ;GSBRN A2 T00030224001;GSBRN A2T 00017 349001 0.21 0. 01 MOLECU LA R_ FUNCT ION GO:0008839 4-hyd roxy -te trahyd r odi picoli nate redu ctase 28.99 0.016599521 6.86E-04 3. 16E+00 3 3 1910 55462 GSBRN A2T 00045154001;G SBRNA 2T 00022021001 ;GSBRN A2 T00101 443001 G SBRNA 2T 00052621001 ;GSBRN A2 T00139025001;GSBRN A2T 00066 748001 0.16 0. 01 MOLECU LA R_ FUNCT ION GO:0004070 as pa rtate carbam oy ltran sferas e acti vit y 28.99 0.09524379 0.006373748 2. 20E+00 2 2 1911 55463 GSBRN A2T 00029544001;G SBRNA 2T 00043712001 G SBRNA 2T 00095869001 ;GSBRN A2 T00075936001 0.10 0. 00