Improvement of Cell Viability Using a Rho-associated Protein Kinase (ROCK) Inhibitor in Human Dental Papilla derived Single-induced Pluripotent Stem Cells
Yoo-Jin Shim1, Young-Hoon Kang2, Hyeon-Ji Kim1, Mi-Jeong Kim1, Hyeon-Jeong Lee3, Young-Bum Son3, Sung-Ho Lee4 and Byeong-Gyun Jeon1,5*
1Department of Biology Education, Gyeongsang National University, Jinju 52828, Korea
2Department of Oral and Maxillofacial Surgery, Gyeongsang National University, Jinju 52828, Korea
3OBS/Theriogenology and Biotechnology, Gyeongsang National University, Jinju 52828, Korea
4Division of Life Science, Gyeongsang National University, Jinju 52828, Korea
5Institute of Education, Gyeongsang National University, Jinju 52828, Korea
Received June 3, 2019 /Revised July 10, 2019 /Accepted July 11, 2019
The aim of the present study was to improve the cell viability of human dental papilla derived sin- gle-induced pluripotent stem cells (iPSCs) using a Rho-associated protein kinase (ROCK) inhibitor, Y-27632. The iPSCs were produced using an episomal plasmid-based reprogramming method. After cell separation using trypsin, the iPSCs were treated with 0, 0.5, 1, 2.5, 5, 7.5, or 10 μM Y-27632 for 5 d. Cell viability increased significantly following the 5 μM Y-27632 treatment (p<0.05). When the iPSCs were exposed to medium containing 10 μM Y-27632 for 0, 1, 2, 3, 4, and 5 d, the cell viability rate increased significantly in accordance with the cell viability rate (p<0.05). To evaluate the effect of the Y-27632 treatment on stemness characteristics, the expression of stem cell-specific transcripts and telomerase activity were investigated in the iPSCs treated with 10 μM Y-27632 for 5 d. The ex- pression levels of stem cell-specific transcripts, such as OCT-4, NONOG, and SOX-2, and telomerase activity were not significantly different in the iPSCs treated with 10 μM Y-27632 as compared with those of untreated control iPSCs (p>0.05). Taken together, the results demonstrated that cell viability can be improved by treatment with the ROCK inhibitor Y-27632, without losing iPSC stemness characteristics.
Key words : Anoikis, human, iPSCs, stem cells, Y-27632
*Corresponding author
*Tel : +82-55-772-2236, Fax : +82-55-772-2239
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2019 Vol. 29. No. 8. 895~903 DOI : https://doi.org/10.5352/JLS.2019.29.8.895
서 론
줄기세포(stem cells)는 우리 몸을 구성하는 모든 종류의 세 포 형태로 분화할 수 있는 능력을 가진 세포를 말하며, 이러한 줄기세포는 특정한 환경과 조건에서 다양한 특정 세포로 분화 할 수 있다. 특히, 우리 몸의 어떤 부분이 손상이나 질병이 생겼 을 때, 그 부분을 약물 등으로 치료하기보다는 손상된 부분을 새로운 세포로 대체하는 재생의학(regenerative medicine) 분 야에서 줄기세포는 큰 주목을 받고 있다. 이러한 줄기세포는 그 기원에 따라 배아줄기세포(embryonic stem cells, ESCs)와 성체줄기세포(adult stem cells)가 있다. 그러나 배아줄기세포 는 무한 증식이 가능하고 우리 몸의 모든 부분으로 분화할 수 있으나, 윤리적인 문제와 면역 거부 반응이 나타나고 있으
며, 반면 성체줄기세포는 배아줄기세포에 비해 분화 능력과 증식 능력이 제한된다는 문제점을 가지고 있다[8, 17]. 그러나, 최근에는 완전히 분화된 체세포로부터 역분화 과정을 거처 배아줄기세포와 유사한 줄기세포학적인 특징을 가지고 있는 유도만능줄기세포(induced pluripotent stem cells, iPSCs)를 만들어 윤리적인 문제를 해결하면서, 환자로부터 직접 줄기세 포를 만들 수 있어 면역 거부 문제를 해결할 수 있다. 이에 iPSCs를 이용한 세포 치료와 함께 질병의 원인 규명과 신약 개발 등에서도 널리 응용될 수 있어, iPSCs는 치과 치료 등의 재생의학 분야에서 크게 주목을 받고 있다[6, 16, 30].
그러나, iPSCs는 ESCs와 유사한 다재다능한 줄기세포학적 인 특징을 가지고 있지만, 지금까지 많은 체외 세포배양기술 의 발달에도 불구하고 배아줄기세포처럼 유도만능줄기세포 를 배양하는 데 어려움이 있다. 일반적으로 iPSCs 세포 사이에 는 많은 밀착결합(tight junction)으로 단단히 결합하고 있으면 서 간극결합(gap junction)으로 세포 사이에 여러 성장에 필요 한 신호와 소통을 통하여 동반 성장을 하고, 여러 부착 분자를 이용하여 배양접시의 바닥에 부착하면서 자라게 되어 보통 iPSCs는 서로 결합하여 세포군(colony)을 형성하게 되면 잘 자라게 된다[3]. 그러나 iPSCs가 적절한 세포군을 형성하지 못
하게 되면, 세포의 분화가 일어나거나 세포가 사멸 단계에 들 어가게 된다[24]. 그래서 일반적으로 ESCs 배양처럼, iPSCs 배 양에도 세포의 미분화를 유지하기 위해 섬유아세포 성장인자 (basic fibroblast growth factor, bFGF) 같은 물질을 배양액에 첨가하면서, 단일 세포보다는 세포군의 상태로 계속 배양을 하여 줄기세포의 특성을 유지하면서 배양접시에 잘 부착하여 지속적인 성장 효과가 나타나도록 한다[2, 23, 29]. 그러나, iPSCs를 증식하기 위한 계대배양을 하면서 세포의 부착분자 를 제거하는 효소를 사용하지 않고, 세포군의 상태로 분리하 는 것을 쉽지 않을 뿐만 아니라, 흔히 계대배양 시 세포군에서 단일 세포로 분리되어 iPSCs의 소실이 지속적으로 일어나게 된다. 이전의 연구에서도 단일세포로 분리된 미부착 iPSCs의 생존율은 매우 낮게 나타난다고 보고하고 있어, 세포간 접착 에 의한 iPSCs의 세포군의 형성과 배양접시에 부착 능력은 iPSCs의 성공적인 체외배양에 매우 중요한 요인이라 보고하 고 있다[2, 23].
거의 모든 동물세포는 부착의존성을 가지고 세포외 기질에 부착하여 성장하며, 정상적인 세포가 적절히 배양접시에 부착 하지 못하거나 부적절한 세포 부착이 일어나면 anoikis이라고 불리는 세포사멸이 유발되며, 일반적인 체세포와 다르게 대부 분의 악성의 암세포는 anoikis 세포사멸에 상당히 저항적이다 [5]. 세포가 배양접시나 세포외 기질(extracellular matrix, ECM) 에 부착하는 가장 기본적인 부착 분자는 인테그린(intergrins) 단백질이다. 또한, 세포 내에는 인테그린과 연결된 액틴(actin) 과 그 액틴의 운동단백질인 미오신(myosin)이 세포의 수축을 유도하게 되면 세포 부착이 잘 일어나지 않는다, 그러나 분화 된 세포는 이 미오신 운동단백질의 활성이 줄어들면서 인테그 린을 통한 ECM에 쉽게 결합하고 부착 상태에 있게 된다[31].
iPSCs는 분화된 세포에서 줄기세포로 reprogramming 과정에 서 운동단백질인 미오신의 활성을 조절하는 단백질 역시 활성 이 증가하여 미오신의 활성이 증가하게 되고 단일 세포로 된 iPSCs는 세포외 기질과 잘 결합하지 않아 anoikis 세포사멸이 흔히 일어난다고 보고하고 있다[7, 15, 27]. 또한, 많은 임상 실험을 위해 다량의 iPSCs가 필요하고, 종종 세포동결 후 융해 과정에서도 세포의 미부착에 의한 세포 손실이 종종 일어난 다. 몇몇 선행 연구에서 세포의 부착과 관련된 운동단백질 미 오신은 Rho 단백질과 이 단백질 관련 인산화효소(Rho asso- ciate kinase, ROCK) 신호전달 경로에 의해 활성화되고 있으 며, 이러한 ROCK 단백질의 활성을 억제할 수 있는 물질로는 Y-27632이 대표적으로 알려져 있다[18, 20]. 이에 몇몇의 연구 에서는 Y-27632를 iPSCs의 배양 효율을 증진하기 위해 혹은 세포동결 후 생존성을 높이기 위해 사용되었다[14, 28].
이에 본 연구는 좀 더 효율적으로 iPSCs를 구축하기 위해 Capetian 등(2017)이 제안한 episomal plasmids 방법으로 치 유두 조직(dental papilla)으로부터 분리한 중간엽성 성체줄기 세포를 사용하여 iPSCs를 생산하였고, 이를 단백질 분해 효소
를 처리하여 단일 유도만능줄기세포로 분리한 다음 Y-27632 ROCK 억제제를 사용하여 iPSCs의 생존성을 MTT 방법을 통 해 조사하였다. 또한, Y-27632를 장기간 노출된 iPSCs가 줄기 세포의 특성을 유지하는지 검증하기 위해 줄기세포 특이 전사 체인 OCT-4, NANOG 및 SOX-2의 발현율을 RT-PCR 방법으 로 조사하였고, 더욱 줄기세포의 무한증식과 관련되는 telo- merase 활성도를 RQ-TRAP 방법으로, 이 telomerase의 활성 과 관련된 TERT (Telomerase reverse transcriptase)과 TERC (Telomerase RNA component)의 발현 수준을 RT-PCR 방법 으로 Y-27632를 처리하지 않은 대조군 iPSCs와 비교 조사하 여, Y-27632의 처리가 줄기세포의 특성에 미치는 효과를 조사 하여, 치아조직 유래 유도만능세포의 임상적 적용을 위한 기 초 연구를 진행하였다.
재료 및 방법
세포의 배양 및 처리
이 실험에서 유도만능줄기세포의 역분화를 유도하기 위한 사용된 세포는 사람의 사랑니의 치유두(dental papilla) 조직 기원 중간엽 줄기세포(mesenchymal stem cells, MSCs)를 사 용하였다. 이 세포의 줄기세포학적인 특징은 이전의 연구에서 증명되었다[10]. MSCs의 기본배양액은 3% FBS와 항생제를 첨가한 Advanced Dulbecco's Modified Eagle 배양액(A- DMEM, Gibco, USA) 이었고, 37℃의 5% CO2 항온배양기에서 배양하면서, 세포가 70~80% 이상 성장하면 계대배양을 실시 하여 5 계대배양 세포주를 이 실험에서 사용하였다. 이 MSCs 를 사용하여 iPSCs의 확립 방법은 전의 연구에서 증명된 Epi5™ Episomal iPSC Reprogramming Kit (Thermo Fischer Scie- ntific, USA)을 응용한 방법을 사용하였다[4]. 간단히, Oct3/4, Sox2, Klf4, L-Myc 및 Lin28의 reprogramming factor를 가진 oriP/EBNA-1 (Epstein-Barr Nuclear Antigen-1) episomal plasmids는 MSCs가 70~90% 정도 성장하였을 때, Neon™ Transfection System (Thermo Fischer Scientific, USA)을 사용 하여 세포 내로 형질 도입을 유도하였다. 이 세포는 Geltrex™ (Thermo Fischer Scientific, USA)로 코팅된 세포배양 접시에 서 배양되었고, 다음 날 100 ng/ml의 bFGF가 첨가된 N2B27 배양액(Thermo Fischer Scientific, USA)으로 교체하였고, 매 일 배양액을 교체하면서 14일 동안 배양하였다. 14일 후, TeSR
™-E8™ 배양액 (STEMCELL Technologies, USA)으로 교체하 였으며, 세포군(colony)을 형성한 iPSCs를 채취하였다. 이렇게 채취된 iPSCs는 역시 Geltrex™로 코팅된 6-well 세포배양 플라 스크 혹은 9.6 cm2 세포배양 접시에서 TeSR™-E8™ 배양액에서 체외배양되었다. 매일 일정 시간에 배양액을 교체하였으며,
~90%의 세포 성장 시 계대배양을 실시하였다(Fig. 1). iPSCs의 일반적인 계대배양은 비효소적인 방법으로 0.5 mM EDTA를 세포배양 접시에 첨가한 다음 대략 5분 동안 배양하여 세포
Fig. 1. iPSCs colony produced by Episomal plasmid.
덩어리를 분리하였고, 이를 1:4의 비율로 분할하여 Geltrex™로 코팅된 새 배양접시에서 계대배양을 하였다. 계대배양 후 일 부 iPSCs는 동결보존되었고, 일부의 iPSCs 중 ~90% 이상 성장 한 20 계대배양 iPSCs는 트립신을 첨가하여 2~3분 동안 배양 하여 단일 세포로 분리를 한 다음 이 실험에 사용되었다.
iPSC에서 Y-27632 적정 처리 농도 분석
단일 세포로 분리된 iPSCs의 생존율을 향상하기 위해 적정 한 Y-27632의 농도를 측정하였다. 우선, 분리된 iPSCs는 6- well 세포배양 플라스크의 각 well에 분주한 다음 0.5 uM, 1 uM, 2.5 uM, 5 uM, 7.5 uM 및 10 uM의 Y-27632를 첨가한 TeSR™-E8™ 배양액에서 5일 동안 각 농도의 배양액이 첨가된 배양액을 매일 교체하면서 세포 배양을 실시하여, 각 농도에 노출된 세포의 생존율은 MTT 분석 방법으로 측정하였다. 5일 후 각 well의 배양액을 제거한 후 D-PBS로 세척하였고, 3 mg/
ml의 MTT 용액을 각 well에 첨가 하여 4시간 동안 추가적으 로 배양하여, 포마잔이 형성된 살아있는 iPSCs 세포군을 현미 경을 확인하였고, 이를 다시 D-PBS로 세척한 다음, DMSO를 200 ul 첨가하여 30분 동안 실온에 두어 포마잔을 녹인 다음, 이를 회수하여 50 ul씩 96-well 플레이트에 옮겼다. 이를 ELISA-리더기를 이용하여 570 nm의 파장으로 그 흡광도를 측정한 다음, 각 농도에 대한 세포의 생존율을 상대적인 값으 로 환산하여 계산하였다.
iPSC에서 Y-27632 적정 처리 시간 분석
단일세포로 분리된 iPSCs은 역시 6-well 세포배양 플라스크 의 각 well에 분주한 다음, 각 well의 iPSCs는 10 uM의 Y- 27632가 첨가된 TeSR™-E8™ 배양액에서 0일, 1일, 2일, 3일, 4일 및 5일 동안 노출하였고, 각 요일에 대한 세포의 생존율을 역 시 위에 서술한 MTT 방법으로 분석하였다.
RT-PCR 방법에 의한 줄기세포 전사인자의 발현 분석 Y-27632의 처리 후 iPSCs의 줄기세포학적인 특성 변화를 유발할 수 있는지 검증하기 위해 우선 줄기세포 특이 전사인
자의 발현을 RT-PCR 방법으로 분석하였다. 이 실험에 사용된 줄기세포 특이 다능성 전사인자는 OCT-4, NANOG 및 SOX-2 이었다. 단일세포로 분리된 iPSCs를 10 uM의 Y-27632가 첨가 된 TeSR™-E8™ 배양액에서 5일 동안 배양한 실험군과 Y-27632 를 처리하지 않은 대조군을 서로 비교 분석하였다. RT-PCR 방법은 실험군과 대조군의 각 세포에서 RNeasy kit (Qiagen, USA)을 사용하여 total RNA을 추출하였다. 이 total RNA를 Qubit flurometer를 사용하여 RNA의 농도 측정한 다음, total RNA를 Omniscript reverse transcriptase kit (Qiagen, USA) 를 이용하여 cDNA 합성을 유도하였다. 20 ul의 각 cDNA 반 응물은 1 ug의 total RNA, 2 ul의 10 uM random hexamer, 1 ul의 10 U/ul RNase inhibitor, 2 ul의 dNTP 및 4 U의 re- verse transcriptase을 포함하고 있었고, 42℃에서 1시간 동안 반응을 유도하였다. cDNA 합성 후, 각 전사인자의 발현은 Maxime-PCR PreMix Kit (iNtRON Biotechnology, Korea)을 이용하여 PCR 방법으로 측정하였다. 각 PCR 반응물은 2 ul의 cDNA 및 1 ul의 10 uM forward 및 reverse 프라이머를 첨가 하여 94℃에서 1분 동안 변성 단계, 56-60℃에서 30분 동안 프라이머 결합 단계 및 72℃에서 1분 동안 신장 단계로 구성하 여 30회 동안 반응을 유도하였다. PCR 수행 후, 각 반응물은 아가로스 겔로 확인하였고, 이미지 프로그램을 이용하여 항존 유전자 glyceraldehyde 3-phosphate dehydrogenase (GAPDH) 에 대한 상대적인 농도를 대조군과 실험군에서 측정하였다.
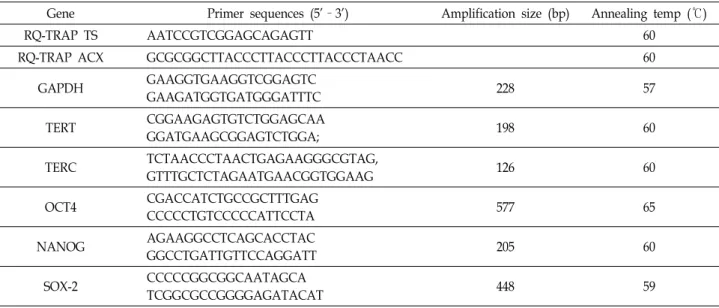
각 유전자에 대한 프라이머 서열은 Table 1에서 나타내었다.
RQ-TRAP을 통한 telomerase 활성도 분석
10 uM의 Y-27632를 5일 동안 처리 후 iPSCs의 telomerase 활성은 RQ-TRAP 방법으로 분석하였다. 우선 대조군과 실험 군의 세포 샘플을 준비한 다음, 0.5% CHAPS lysis buffer를 첨가하여 30분 동안 4℃에서 처리하여 세포의 파쇄를 유도하 였다. 이후 샘플들을 4℃에서 14,000 g의 속도로 30분간 원심 분리를 하였고, 상층액의 70%를 채취한 다음 각 샘플의 telo- merase를 포함한 단백질 농도를 분광광도계로 측정하였다. 단 백질 추출액 1 ug과 2 ul의 Lightcycler FastStart DNA Master SYBR Green 1(Qiagen, UASA), 0.02 ug의 TS 프라이머 및 0.4 ug의 ACX 프라이머를 혼합하여 총 20 ul의 반응물을 만들고, 실시간 PCR 방법(Qiagen, UASA)을 이용하여 telomerase의 활성을 측정하였다. Telomerase 반응 조건은 30℃에서 30분 동안 전처리 과정을 하였고, 증폭 조건은 94℃에서 30초 동안 변성 과정, 60℃에서 90초 동안 결합 과정과 72℃에서 0초 동 안의 과정을 40회 반복하였다. 증폭 후 Quantification Soft- ware (Qiagen, UASA)로 Ct 값을 분석하여 A-549 폐암세포주 와 MRC-5 정상세포주의 telomerase 활성과 비교 분석하였다.
통계 분석
대조군 및 실험군의 모든 실험은 3회 이상 반복하여 실시하
Table 1. Primer sequences, PCR product size and annealing temperature used for RQ-TRAP and RT-PCR
Gene Primer sequences (5'–3') Amplification size (bp) Annealing temp (℃)
RQ-TRAP TS AATCCGTCGGAGCAGAGTT 60
RQ-TRAP ACX GCGCGGCTTACCCTTACCCTTACCCTAACC 60
GAPDH GAAGGTGAAGGTCGGAGTC
GAAGATGGTGATGGGATTTC 228 57
TERT CGGAAGAGTGTCTGGAGCAA
GGATGAAGCGGAGTCTGGA; 198 60
TERC TCTAACCCTAACTGAGAAGGGCGTAG,
GTTTGCTCTAGAATGAACGGTGGAAG 126 60
OCT4 CGACCATCTGCCGCTTTGAG
CCCCCTGTCCCCCATTCCTA 577 65
NANOG AGAAGGCCTCAGCACCTAC
GGCCTGATTGTTCCAGGATT 205 60
SOX-2 CCCCCGGCGGCAATAGCA
TCGGCGCCGGGGAGATACAT 448 59
Fig. 2. Attachment of iPSCc by Y-27632 treatment. Single iPSCs isolated after trypsin treatment (A). Attached (B) and de- tached (C) iPSCs at 1 day after seeding. Attached iPSCs treated with Y-27632 at 2 days (C), 3 days (D) and 5 days (E) after seeding. Fibroblast-like iPSCs were ob- served after Y-27632 treatment (C and D). Scale bar in- dicates 100 μm.
Fig. 3. Cell viability and formation of cell colony in single iPSCs cultured for 0 day (A), 1 day (B), 2 days (C), 3 days (D), 4 days (E) and 5 days (F) in TeSR™-E8™ medium supple- mented with 10 uM Y-27632 by MTT assay. Cell viability was significantly increased by increased exposure day.
Scale bar indicates 100 μm.
여, 얻어진 모든 결과는 평균±표준오차를 구하였다. 각 처리군 에 대한 유의성은 SPSS (21.0)를 이용하여 던컨 다중범위검정 방 법으로 분석하였고, p<0.05의 수준에서 유의성을 검정하였다.
결 과
Y-27632 처리의 적정 시간
iPSCs를 트립신 처리하여 단일 세포로 분리한 다음, 10 uM 의 Y-27632를 5일 동안 처리하여 Y-27632의 노출 일수에 대한 iPSCs의 세포의 생존율을 조사하였다(Fig. 2). 단일 세포로 분 리한 후, Y-27632를 처리한 세포는 Fig. 2의 B에서 나타난 것처 럼 세포가 부착하였으나, Y-27632를 처리하지 않은 iPSCs는
Fig. 2의 C에서 나타난 것처럼 세포가 부착하지 못하여 세포가 부정형으로 변하면서 세포의 사멸 과정이 일어나는 것을 알 수 있었다.
또한, 각 요일 별로 10 uM의 농도로 Y-27632를 5일 동안 처리한 다음, 6-well에 MTT 용액을 처리하고 iPSCs 세포군의 크기 및 농도를 관찰하고, 흡광도를 이용하여 각 요일 별 iPSCs 의 세포 생존율을 조사한 결과는 Fig. 3에 나타난 바와 같다.
Fig. 4. Cell viability and formation of cell colony of iPSCs in E8 medium containing 0.5 (A), 1.0 (B), 2.5 (C), 5 (D), 7.5 (E) and 10.0 (F) uM Y-27632 for 5 days. Cell viability was also significantly increased over 5 uM Y-27632.
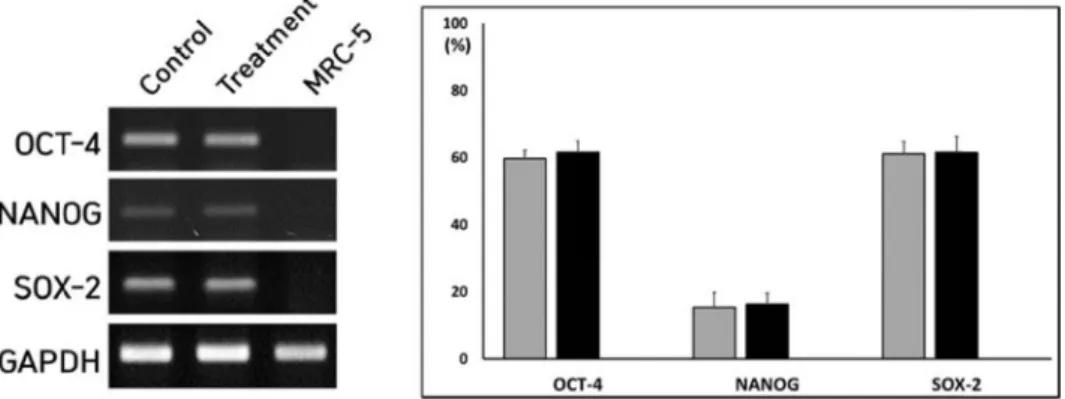
Fig. 5. Expression level of OCT-4, NANOG and SOX-2 stemness transcripts in untreated control (■) and Y-27632-treated iPSCs (■) treated 10 uM Y-27632.
Y-27632를 첨가하지 않은 경우, 아주 작은 세포군이 관찰되었 다. 요일 별 각 세포의 생존율은 Y-27632를 5일 동안 처리한 세포에서 세포의 생존율을 100%로 기준하여, Y-27632를 0 처 리하지 않은 경우 생존율은 1±0.15%이었으나, Y-27632를 1일, 2일, 3일 및 4일 동안 처리하였을 때 세포의 생존율은 각각 5±1.45%, 45±3.98%, 65±5.11% 및 89±6.41%를 나타내어 노출 일수가 증가할수록 세포의 생존율이 유의적으로(p<0.05)증가
하는 것을 알 수 있었다.
Y-27632 처리의 적정 농도
단일세포로 분리된 iPSCs에 Y-27632의 농도를 0.5 uM, 1 uM, 2.5 uM, 5 uM, 7.5 uM 및 10 uM의 농도로 Y-27632를 TeSR™-E8™ 배양액에 첨가한 다음 5일 동안 각각 배양하였고, 그 세포의 생존율을 위에서 서술한 MTT 방법으로 조사하였다 (Fig. 4). 10 uM의 Y-27632를 첨가하였을 때, 세포의 생존율을 100%로 계산하여 각각의 농도에서 상대적인 세포의 생존율을 측정하였다. 0.5 uM, 1 uM, 2.5 uM, 5 uM 및 7.5 uM에서 세포 의 생존율은 각각 15±2.18%, 40±6.64%, 80±5.45%, 98±3.48%
및 101±6.52%를 나타내어 Y-27632를 5 uM 이상으로 첨가하 였을 때 세포의 생존율과 세포군의 크기가 유의적으로 (p<0.05) 증가하는 것을 관찰할 수 있었다.
줄기세포 특이 전사인자의 발현 분석
이상의 결과에서 Y-27632 ROCK 억제제의 처리는 iPSCs의 생존율이 향상되는 것을 관찰하였다. 그러나 Fig. 2D 및 Fig.
2E에서 나타난 바와 같이, Y-27632 처리 후, 세포의 형태적인 변화가 관찰되었다. 이러한 형태적인 변화는 분화된 세포에서 흔히 나타나는 현상이어서, Y-27632 처리한 iPSCs에서 줄기세 포의 세포학적인 특징을 나타내는지 검증하였다. 우선 단일세 포로 분리된 iPSCs에 10 uM의 Y-27632를 5일 동안 노출시킨 후, OCT-4, NANOG 및 SOX-2 줄기세포 특이 전사인자의 발 현을 RT-PCR 방법으로 분석하였다(Fig. 5). 먼저, OCT-4, NANOG 및 SOX-2의 줄기세포 전사인자의 발현은 정상 체세 포인 MRC-5에서 발현이 되지 않는 것을 확인하였다. OCT-4, NANOG 및 SOX-2의 발현량을 항존유전자인 GAPDH의 발 현량과 상대적으로 비교하였다. Y-27632를 처리하지 않은 대 조군 iPSCs에서 OCT-4, NANOG 및 SOX-2의 발현량은 각각 60±2.4, 15±4.5 및 61±3.7%이었다. 또한, Y-27632를 처리한 실 험군의 iPSCs에서 OCT-4, NANOG 및 SOX-2의 발현량은 각 각 62±3.3, 16±3.4 및 62±4.8%를 나타내어, 실험군과 대조군에
A
B
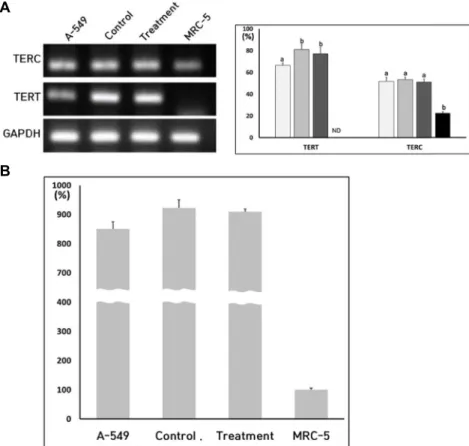
Fig. 6. A, Expression level of TERT and TERC transcripts in A-549 lung cancer (□), untreated control (■), Y-27632 treat- ment (■) and MRC-5 fibroblasts (■).
a and b significant (p<0.05) difference among cell lines, respectively. B, Telo- merase activity in A-549 lung cancer, untreated control iPSCs, Y-27632 treat- iPSCs and normal MRC-5 fibroblasts.
서 OCT-4, NANOG 및 SOX-2의 발현량은 유의적인 차이가 없어, Y-27632를 5일동안 처리하여도 줄기세포 특이 유전자의 발현에는 변화가 없음을 알 수 있었다.
Telomerase 활성 및 TERT와 TERC 발현 분석 단일 세포로 분리한 후 세포의 증식과 관련되는 telomerase 활성을 RQ-TRAP 방법으로 분석하였고 이 telomerase 활성과 관련되는 TERT 및 TERC의 발현을 RT-PCR 방법으로 조사하 였다(Fig. 6). Telomerase의 활성도는 MRC-5 정상체세포에서 100%로 계산하고, 이 값에 대한 상대적인 값으로 산정하였다.
비교군으로 사용한 무한증식을 나타내는 A-549 폐암세포주에 서는 850±25.6%를 나타내어 정상세포의 거의 8배에 이러는 높은 telomerase의 활성도를 나타내었다. 또한, Y-27632를 처 리한 실험군에서 telomerase 활성도는 909±10.1%이었고, Y- 27632를 처리하지 않은 대조군에서 telomerase 활성도는 923±27.6%로 측정되어, Y-27632의 처리가 telomerase의 활성 에 어떠한 변화를 유발하지 않았다. 좀 더, telomerase의 활성 과 관련된 TERT 및 TERC의 발현을 조사하였는데, telomerase 활성을 나타내지 않는 정상체세포에서 TERT의 뱔현은 검출 되지 않았고, TERC의 발현은 23±1.24% 수준으로 측정되었고, 높은 telomerase 활성을 나타내는 A-549 폐암세포의 TERT의 발현은 67±2.1% 수준이었고, TERC의 발현은 52±3.6% 수준으 로 측정되어, TERC의 발현보다 TERT의 발현이 telomerase 활성에 크게 관련됨을 알 수 있었다. 또한, Y-27632를 처리하
지 않은 대조군의 TERT 및 TERC 발현은 81±4.5% 및 53±2.3%
수준이었고, Y-27632를 처리한 실험군의 TERT 및 TERC 발현 은 77±5.6% 및 51±2.8% 수준으로 측정되었다. 정상체세포의 TERT는 0%, TERC는 23±1.2%로 측정되었다. TERT 및 TERC 의 발현은 Y-27632를 처리하여도 유의적인 차이를 나타내지 않았다.
고 찰
유도만능줄기세포를 이용한 세포 치료, 세포 이식 등의 임 상 연구 분야에서나 분화의 기작, 발달과 발생 연구 혹은 외래 형질 도입 등의 다양한 분자세포생물학적인 기초 연구를 하기 위하여 iPSCs를 대량으로 배양할 수 있는 효율적인 체외 배양 기술의 발달이 필요하다. 일반적으로 iPSCs는 인접한 세포들 이 세포군(colony)을 형성하여 자라게 되지만, iPSCs는 단일 세포로 분리된 후 세포군(colony)의 형성하는 것이 매우 어렵 고, 또한 세포의 생존 가능성이 현저히 떨어지게 되어 좀 더 효율적으로 iPSCs의 세포군을 형성하면서 성장을 촉진할 수 있는 세포배양 기술이 절실히 요구된다. 하지만, 이 연구에서 트립신 처리 후, 하나의 단일 세포로 분리된 iPSCs에 ROCK 단백질의 억제제인 Y-27632를 처리하였을 때, iPSCs는 세포군 을 쉽게 형성하면서 세포의 생존율이 현저하게 향상되는 것을 관찰하였다.
일반적으로 동물 세포는 세포의 운동과 형태에 관련된 여러
종류의 세포골격(cytoskeleton)을 가지고 있다. 이러한 세포골 격은 세포 분열, 세포의 신호 전달 등에서도 중요한 역할을 한다. 이 세포골격의 변화는 여러 세포 내 조절물질로 활성이 되는 데, 이 중 GTP 단백질의 한 종류인 Rho 단백질은 이 단백 질과 관련된 protein serine/threonine ROCK1 및 ROCK2 인 산화효소를 자극하게 된다. 활성화된 ROCK 단백질은 세포 내 액틴필라멘트(actin filament) 세포골격과 상호작용하는 미 오신(myosin) 운동단백질의 탈인산화를 억제하여 미오신 인 산화를 계속 유지하게 한다. 세포는 일반적으로 인산화된 미 오신 단백질이 세포의 수축을 유도하는데, 이러한 미오신 단 백질의 탈인산화를 억제하여 인산화된 미오신으로 인하여 세 포는 계속적인 수축을 하게 되어, 세포는 부착하지 못한 상태 가 된다[12, 18]. 즉, 부착성을 상실한 많은 암세포나 세포분열 중인 미부착 정상세포는 이러한 미오신의 활성을 조절하는 ROCK 단백질의 활성이 높다[9, 18, 21, 26]. iPSCs는 분화된 세포를 인위적으로 reprogramming의 과정에서 통하여 생성 된 줄기세포이고, 이 reprogramming 과정에서 역시 이 ROCK 단백질이 활성이 높아지게 된다. 그러므로 iPSCs는 일반적으 로 세포가 세포분열 때처럼 수축된 상태로 배양접시에 부착력 이 감소하게 되고, 미부착된 세포는 세포사멸 단계(anoikis)에 접어들게 된다[27]. 그러나 Y-27632 물질은 세포 이동, 세포 극성 및 세포 분화 등에 관여하는 Rho 단백질의 하위 조절자 인 이 ROCK 단백질을 억제함으로써 세포의 부착을 향상시키 고, 이에 따라 세포의 생존성이 향상될 수 있다고 하며, 특히 전의 다른 연구에서도 iPSCs에 Y-27632를 처리하면 단일 세포 로 분리된 세포의 생존성과 세포군을 형성하는데 도움을 줄 수 있다고 보고하고 있다[19, 28]. Y-27632 화합물은 처음으로 발견된 가장 잘 알려진 ROCK 억제제이기도 하며, Fasudil이 라고 알려진 HA-1077 역시 강력한 ROCK 억제제이다[22]. 이 에 이전의 연구에서 Y-27632와 HA-1077를 각각 ESCs 혹은 iPSCs에 처리하였을 때, 그 세포 성장과 세포군의 형성에 미치 는 그 효과는 유사하다고 하여, ROCK 단백질의 억제는 효율 적으로 iPSCs의 세포군을 형성하는데 도움을 줄 수 있다고 보고하고 있다[22]. 이 연구에서도 역시 5 uM 이상의 농도에서 Y-27632 처리하였을 때 트립신 처리 후 단일세포로 분리된 iPSCs의 생존성이 향상되는 것을 관찰하였다. 그러나 전의 연 구에서 Y-27632의 처리 기간은 대부분 1일 정도 이었으나[14], 이 연구에서는 효소를 사용하여 하나의 단일세포로 분리한 iPSCs에 Y-27632을 노출하였을 때, 노출 시간이 길어질수록 anoikis의 감소로 인한 세포의 생존성이 현저히 향상되는 것을 관찰하였다.
그러나, 이 연구에서 Y-27632 처리가 단일세포로 분리된 iPSCs가 세포군 형성으로 인하여 세포의 생존성이 향상되었 지만, Y-27632를 장기간 처리하였을 때, 줄기세포의 특성에 변화를 줄 수 있어, 10 uM의 Y-27632로 5일간 처리된 iPSCs에 서 줄기세포 특이 전사인자의 발현과 세포사멸없이 줄기세포
의 무한 증식과 밀접하게 관련되는 telomerase 활성을 분석하 였다. 전의 몇몇 연구에서도 OCT-4, NANOG 및 SOX-2 같은 줄기세포 전사인자는 줄기세포의 기본적인 세포 특성에 가장 중요한 요인이라고 보고하고 있다[10, 25]. 현재의 연구에서도 Y-27632를 처리 후, OCT-4, NANOG 및 SOX-2 줄기세포 특이 전사인자의 발현은 유의적인 변화를 나타내지 않았다. 또한, 배아줄기세포와 같이 유도만능줄기세포는 세포의 노화가 일 어나지 않아 무한 증식이 가능하고, iPSCs의 무한 증식 능력은 또한 줄기세포의 자가재생 능력(self-renewal capacity)과 밀접 하게 관련되어 있다[1]. 이러한 무한 증식 능력은 말단소립 (telomere)의 길이와 관련되는데, iPSCs의 reprogramming 과 정에서 telomerase의 활성도 증가하게 된다. 이 연구에서도 암세포와 같이 무한 증식을 하는 세포와 비교하게 telomerase 의 활성이 나타났고, 분화된 정상체세포와 비교하여 상당히 높은 수준의 telomerase 활성을 나타내었다. 반대로 무한 증식 을 나타내는 암세포에서 telomerase 활성에 영향을 주는 여러 물질로 telomerase 활성을 줄이게 되면 telomere의 길이는 짧 아지면서 세포사멸(apoptosis)에 이른다고 보고하고 있다[11].
이 연구에서 Y-27632를 처리 한 다음, iPSCs에서 telomerase의 활성은 변화가 없어, Y-27632 처리를 하였을 때, 세포의 무한 증식과 자가 재생 능력에 유의적인 차이가 없을 것이라고 생 각된다. 또한 telomerase 활성에 가장 크게 영향을 미치는 두 유전자 TERT 및 TERC의 발현의 발현과 관련된다고 하고 있 다[13]. 본 연구에서도 Y-27632 처리 후, telomerase 활성에 영 향을 주는 TERT 및 TERC의 발현은 처리하지 않았을 경우와 비교하여 유의적인 차이를 나타내지 않았다.
이 실험을 통해 단백질 분해 효소를 이용하여 유도만능줄기 세포를 쉽게 단일 세포로 분리하여 Y-27632 ROCK 단백질 억 제제의 처리를 통해, 유도만능줄기세포의 세포군 형성과 더불 어 세포의 생존성이 향상되는 것을 관찰하였다. 또한, Y-27632 를 처리 후 줄기세포 특이 전사인자와 telomerase 활성은 유의 적인 변화를 나타내지 않아 Y-27632 처리가 줄기세포의 특징 에 유의적인 변화를 나타내지 않은 것으로 알 수 있었다. 이에 이 방법을 통해 좀 더 효율적으로 유도만능줄기세포의 체외 배양 시술에 적용할 수 있을 것으로 판단된다. 그러나, Y- 27632를 처리하지 않은 iPSCs의 세포군은 동그란 모양으로 성장하게 되지만, Fig. 2C와 2D에서 나타난 것처럼, Y-27632의 처리 후 iPSCs의 세포군 형태는 종종 분화된 섬유아세포 (fibroblast) 비슷하게 가장자리 부분이 길고 얇은 중간엽줄기 세포와 비슷한 모양으로 관찰되어, Y-27632 처리가 세포의 분 화에 미치는 영향을 좀 더 심도 있는 조사가 필요할 것으로 판단된다.
References
1. Allsopp, R. 2012. Telomere length and iPSC re-program-
ming: survival of the longest. Cell Res. 22, 614-615.
2. Amit, M., Carpenter, M. K., Inokuma, M. S., Chiu, C. P., Harris, C. P., Waknitz, M. A., Itskovitz-Eldor, J. and Thomson, J. A. 2000. Clonally derived human embryonic stem cell lines maintain pluripotency and proliferative potential for prolonged periods of culture. Dev. Biol. 227, 271-278.
3. Beers, J., Gulbranson, D. R., George, N., Siniscalchi, L. I., Jones, J., Thomson, J. A. and Chen, G. 2012. Passaging and colony expansion of human pluripotent stem cells by en- zyme-free dissociation in chemically defined culture con- ditions. Nat. Protoc. 7, 2029-2040.
4. Capetian, P., Azmitia, L., Pauly, M. G., Krajka, V., Stengel, F., Bernhardi, E. M., Klett, M., Meier, B., Seibler, P., Stans- lowsky, N., Moser, A., Knopp, A., Gillessen-Kaesbach, G., Nikkhah, G., Wegner, F., Döbrössy, M. and Klein, C. 2016.
Plasmid-based generation of induced neural stem cells from adult human fibroblasts. Front. Cell Neurosci. 10, 245.
5. Gilmore, A. P. 2005. Anoikis. Cell Death Differ. 12 Suppl 2, 1473-1277.
6. Ha, H. Y., Jang, S. H. and Jung, J. W. 2011. The use of pluri- potent stem cell for personalized cell therapies against neu- rological disorders. J. Biomed. Biotechnol. 2011, 520816.
7. Heng, B. C., Ye, C. P., Liu, H., Toh, W. S., Rufaihah, A. J., Yang, Z., Bay, B. H., Ge, Z., Ouyang, H. W., Lee, E. H. and Cao, T. 2006. Loss of viability during freeze-thaw of intact and adherent human embryonic stem cells with conven- tional slow-cooling protocols is predominantly due to apop- tosis rather than cellular necrosis. J. Biomed. Sci. 13, 433-445.
8. Huh, Y. J. and Kim, D. W. 2011. Stem cells: general in- formation and perspectives. J. Kor. Med. Assoc. 54, 450-453.
9. Itoh, K., Yoshioka, K., Akedo, H., Uehata, M., Ishizaki, T.
and Narumiya, S. 1999. An essential part for Rho-associated kinase in the transcellular invasion of tumor cells. Nat. Med.
5, 221-225.
10. Jeon, B. G., Jang, S. J., Park, J. S., Subbarao, R. B., Jeong, G. J., Park, B. W. and Rho, G. J. 2015. Differentiation poten- tial of mesenchymal stem cells isolated from human dental tissues into non-mesodermal lineage. Anim. Cell. Syst. 19, 321-331.
11. Kim, Y. D., Jang, S. J., Lim, E. J., Ha, J. S., Shivakumar, S.
B., Jeong, G. J., Rho, G. J. and Jeon, B. G. 2017. Induction of telomere shortening and cellular apoptosis by sodium meta-arsenite in human cancer cell lines. Anim. Cell. Sys.
21, 241-254.
12. Kureishi, Y., Kobayashi, S., Amano, M., Kimura, K., Kanaide, H., Nakano, T., Kaibuchi, K. and Ito, M. 1997. Rho-asso- ciated kinase directly induces smooth muscle contraction through myosin light chain phosphorylation. J. Biol. Chem.
272, 12257-12260.
13. Lee, H. J. and Jeon, B. G. 2018. Comparison of telomere length and telomerase activity in human normal and cancer cell lines. Contemporary Educational Research 30, 185-200.
14. Li, X., Meng, G., Krawetz, R., Liu, S. and Rancourt, D. E.
2008. The ROCK inhibitor Y-27632 enhances the survival rate of human embryonic stem cells following cryopreserva- tion. Stem Cells Dev. 17, 1079-1085.
15. Li, X., Meng, G., Krawetz, R., Liu, S. and Rancourt, D. E.
2009. ROCK inhibitor improves survival of cryopreserved serum/feeder-free single human embryonic stem cells.
Hum. Reprod. 24, 580-589.
16. Little, D., Ketteler, R., Gissen, P. and Devine, M. J. 2019.
Using stem cell-derived neurons in drug screening for neu- rological diseases. Neurobiol. Aging 78, 130-141.
17. Maldonado, M., Luu, R. J., Ramos, M. E. and Nam, J. 2016.
ROCK inhibitor primes human induced pluripotent stem cells to selectively differentiate towards mesendodermal lin- eage via epithelial-mesenchymal transition-like modulation.
Stem Cell Res. 17, 222-227.
18. Matsuoka, T. and Yashiro, M. 2014. Rho/ROCK signaling in motility and metastasis of gastric cancer. World J. Gas- troenterol. 20, 13756-12766.
19. Noritake, J., Watanabe, T., Sato, K., Wang, S. and Kaibuchi, K. 2005. IQGAP1: a key regulator of adhesion and migration.
J. Cell Sci. 118(Pt 10), 2085-2092.
20. Olson, M. F. and Sahai, E. 2008. The actin cytoskeleton in cancer cell motility. Clin. Exp. Metastasis 26, 273-287.
21. Olson, M. F. 2008. Applications for ROCK kinase inhibition.
Curr. Opin. Cell Biol. 20, 242-248.
22. Pakzad, M., Totonchi, M., Taei, A., Seifinejad, A., Hassani, S. N. and Baharvand, H. 2010. Presence of a ROCK inhibitor in extracellular matrix supports more undifferentiated growth of feeder-free human embryonic and induced pluri- potent stem cells upon passaging. Stem Cell Rev. 6, 96-107.
23. Pyle, A. D., Lock, L. F. and Donovan, P. J. 2006. Neurotro- phins mediate human embryonic stem cell survival. Nat.
Biotechnol. 24, 344-350.
24. Sathananthan, H., Pera, M. and Trounsin, A. 2002. The fine structure of human embryonic stem cells. Reprod. Biomed.
Online 4, 56-61.
25. Schmidt, R. and Plath, K. 2012. The roles of the reprogram- ming factors Oct4, Sox2 and Klf4 in resetting the somatic cell epigenome during induced pluripotent stem cell gen- eration. Genome Biol. 13, 251.
26. Thumkeo, D., Watanabe, S. and Narumiya, S. 2013. Physio- logical roles of Rho and Rho effectors in mammals. Eur. J.
Cell Biol. 92, 303-315.
27. Walker, A., Su, H., Conti, M. A., Harb, N., Adelstein, R.
S. and Sato, N. 2010. Non-muscle myosin II regulates surviv- al threshold of pluripotent stem cells. Nat. Commun. 1, 71.
28. Watanabe, K., Ueno, M., Kamiya, D., Nishiyama, A., Matsu- mura, M., Wataya, T., Takahashi, J. B., Nishikawa, S., Muguruma, K. and Sasai, Y. 2007. A ROCK inhibitor per- mits survival of dissociated human embryonic stem cells.
Nat. Biotechnol. 25, 681-686.
29. Xu, R. H., Peck, R. M., Li, D. S., Feng, X., Ludwing, T. and Thomson, J. A. 2005. Basic FGF and suppression of BMP signaling sustain undifferentiated proliferation of human ES cells. Nat. Methods 2, 185-190.
30. Yamanaka, S. 2012. Induced pluripotent stem cells: past, present, and future. Cell Stem Cell. 10, 678-684.
31. Zhong, X. and Rescorla, F. J. 2012. Cell surface adhesion molecules and adhesion-initiated signaling: understanding of anoikis resistance mechanisms and therapeutic opportu- nities. Cell Signal. 24, 393-401.
초록:ROCK 억제제를 통한 사람 치유두 조직 유래 단일 사람 유도만능줄기세포의 생존성 향상
심유진1․강영훈2․김현지1․김미정1․이현정3․손영범3․이성호4․전병균1,5*
(1경상대학교 생물교육과, 2경상대학교 치과학교실, 3경상대학교 수의학과, 4경상대학교 생명과학부, 5경상대학교
교육연구원)
이 연구는 단일 세포로 분리된 유도만능줄기세포(induced pluripotent stem cells, iPSCs)에 anoikis 세포사멸을 억제할 수 있는 Rho-associated protein kinase (ROCK)의 억제제를 처리하여 iPSCs의 세포 생존성을 향상하고자 하였다. Episomal plasmid 방법으로 확립된 iPSCs를 단일세포로 분리한 후, ROCK 억제제 Y-27632 dihydro- chloride (Y-27632)를 0 uM, 0.5 uM, 1 uM, 2.5 uM, 5 uM, 7.5 uM 및 10 uM 농도별로 5주일 동안 각각 처리하였 을 때, 5 uM 이상의 농도에서 세포의 생존율이 유의적으로 향상되었고, 10 uM의 Y-27632을 0일, 1일, 2일, 3일, 4일 및 5일 동안 처리하였을 때, Y-27632의 노출 기간이 길어질수록 세포의 생존율이 유의적으로 향상되는 것을 관찰하였다. 그러나, Y-27632의 노출 후, iPSCs의 형태학적 분화가 관찰되어 10 uM의 Y-27632에서 5일 동안 iPSCs 에 처리 한 후, 줄기세포학적인 특성을 비교 조사하였다. 우선, octamer-binding transcription factor 4 (OCT-4), homeobox protein NANOG (NONOG) 및 SRY-box 2 (SOX-2) 줄기세포 특이 유전자의 발현은 Y-27632를 처리한 실험군은 Y-27632를 처리하지 않은 대조군에서 서로 유의적인 차이를 나타내지 않았다. 또한, Y-27632를 처리한 실험군은 Y-27632를 처리하지 않은 대조군과 비교하여 telomerase 활성과 이것의 활성과 관련된 telomerase re- verse transcriptase (TERT) 및 telomerase RNA component (TERC)의 유전자 발현에는 유의적인 차이가 없었다.
이상의 결과로 보아, iPSCs에 Y-27632를 처리하였을 때, iPSCs의 줄기세포의 특정을 유지하면서 anoikis에 의한 세포사멸을 감소시켜 세포 생존율이 증가한다는 것을 알 수 있었다.