Antioxidant and Anti-inflammatory Effects of Extracts from the Flowers of Weigela subsessilis on RAW 264.7 Macrophages
Yung Choon Yoo
1†, Gye Won Lee
2†and Young Ho Cho
2*
1
Department of Microbiology, College of Medicine, Konyang University, Daejeon 302-832, Korea
2
Department of Pharmaceutics & Biotechnology, College of Medical Engineering, Konyang University, Daejeon 302-832, Korea Received November 13, 2015 /Revised December 16, 2015 /Accepted January 5, 2016
This study investigated the antioxidant and anti-inflammatory activity of ethanol extract from the flowers of Weigela subsessilis (WS-E) in lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophages.
The total polyphenol and flavonoid content was 719.19±0.04 μg tannic acid equivalents/ml and 644.87±0.02 μg quercetin equivalents/ml, respectively. The antioxidant activities of WS-E were meas- ured by 1,1-diphenyl-2-picrylhydrazyl (DPPH) and superoxide anion radical scavenging activity. The antioxidant activities of WS-E increased markedly, in a dose-dependent manner. To screen for anti-in- flammatory agents, the inhibitory effects of WS-E on the production of proinflammatory cytokines in the LPS-stimulated RAW 264.7 macrophages was examined. WS-E had no effect on cell viability at a concentration of 100 μg/ml. Nitric oxide (NO) and interleukin (IL)-6 production were inhibited in a dose-dependent manner (p<0.05). WS-E had no effect on the production of tumor necrosis factor (TNF)-α at a concentration of 0.16–20 μg/ml but induced TNF-α at a concentration of 100 μg/ml.
Inducible nitric oxide synthase (iNOS) expression was also inhibited at lower concentrations (p<0.05).
In addition, WS-E reduced the activation of nuclear factor (NF)-κB by inhibition of inhibitoy (I) κB phosphorylation in RAW 264.7 macrophages upon stimulation with LPS (100 ng/ml) for 24 h but not that of mitogen-activated protein kinase (MAPK). These results suggest that WS-E may be a useful antioxidant and anti-inflammatory agent in functional cosmetics.
Key words : Anti-inflammation, antioxidant, nitric oxide, nuclear factor-κB, Weigela subsessilis
†
Authors contributed equally.
*Corresponding author
*Tel : +82-42-600-8503, Fax : +82-42-600-8503
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2016 Vol. 26. No. 3. 338~345 DOI : http://dx.doi.org/10.5352/JLS.2016.26.3.338
서 론
염증반응은 외부의 물리화학적 자극이나 세균감염에 대한 국소적인 방어보호 반응으로 활성 산소종(reactive oxygen species, ROS), inflammatory cytokine, prostaglandin E
2(PGE
2) 등 다양한 매개물질이 관여하는 것으로 알려져 있다 [6]. 염증반응에 관여하는 대표적인 세포 중 하나인 대식세포 는 염증성 매개물질인 interleukin (IL)-6, IL-1β, tumor ne- crosis factor-α (TNF-α) 등과 같은 pro-inflammatory cytokine 과 nitric oxide (NO), inducible nitric oxide synthase (iNOS), PGE
2등의 염증유발인자들을 생성한다[25]. 정상적인 NO는 세균을 죽이거나 종양을 제거하는 중요한 역할을 하지만 외부 요인에 의한 과도한 NO의 생성은 염증을 유발시켜 신경손상, 조직 손상 및 유전자 변이를 일으키는 것으로 알려져 있다[26].
한편 강한 항산화 활성을 가지는 폴리페놀 화합물은 염증반 응을 매개하는 cytokine, iNOS, cyclooxygenase-2 (COX-2), NO 및 PGE
2의 활성을 저해한다고 보고되었다[8]. 폴리페놀 화합물은 식물계에 존재하는 천연색소로서 주로 세포막에서 유리형, 에스테르형 또는 결합형으로 존재하며, 탄소 수에 따 라 페놀산, 탄닌, 리그난, 안토시아닌, 레스베라트롤 및 플라보 노이드 등의 다양한 물질로 나뉠 수 있다[21]. 특히, 플라보노 이드는 활성산소의 소거 및 산화적 스트레스를 막는 역할을 함으로써 항염증, 항산화, 항노화, 항암, 항고혈압 및 항당뇨 등의 여러 생리적 및 약리적 효능이 있는 것으로 밝혀져, 최근 에는 다양한 식물을 대상으로 ROS를 조절할 수 있는 항산화 활성과 기능성 소재로의 연구가 식품, 의약품, 화장품 등 여러 분야에서 진행되고 있다[4, 18, 20].
병꽃나무(Weigela subsessilis L. H. Bailey)는 인동과(Capri-
foliaceae)에 속하는 낙엽관목으로 주로 한국과 일본에 분포하
고 있고, 높이 2~3 m, 잎은 대생하고 난형 또는 타원형으로
길이 1~7 cm, 폭 1~5 cm이며 양끝은 뾰족하고, 가장자리에
잔 톱니가 있으며 양면에 털이 많이 난다. 꽃은 5월에 피고
황록색이 돌지만 적색으로 변하며 1~2개씩 액생하고 화엽에
털이 있으며, 꽃받침은 선형으로서 밑 부분까지 갈라진다. 열
매는 잔털이 있고 길이 10~15 mm로써 9월에 익으며 종자에
날개가 있다. 우리나라에는 붉은병꽃나무(Weigela florida
(Bunge) A. DC.), 색병꽃나무(Weigela florida for. Alba Rehder), 병꽃나무 및 골병꽃나무(Weigela hortensis (Siebold & Zucc.) K. Koch) 등 4종이 자생하고 있는 것으로 보고되어 있다. 민간 에서는 봄에 채취하여 반 그늘에 말리거나 생 것을 간염, 황달, 소화불량 및 식중독 치료에 사용하였다[15]. 현재까지 병꽃나 무 잎과 줄기에 대한 항산화 연구는 있었으나 병꽃나무 꽃에 대한 화학적 성분연구나 이를 활용한 활성연구는 거의 이루어 지지 않고 있는 실정이다.
따라서 본 연구에서 저자들은 병꽃나무 꽃을 에탄올로 추출 하여 총 플라보이드와 폴리페놀 함량을 조사하고, 항산화 및 항염증 효과를 확인하여 기능성 화장품 소재로의 적용 가능성 을 타진하였다.
재료 및 방법
시약 및 재료
시약으로 사용된 1,1-diphenyl-1-picrylhydrazyl (DPPH), xanthine, xanthine oxidase, nitro blue tetrazolium (NBT), po- tassium persulfate, 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl- tetrazolium bromide (MTT) 등은 Sigma-Aldrich Chemical 사 (St. Louis, MO, USA)에서 각각 구입하여 사용하였다. 그 외 실험에 사용된 모든 시약들은 일급 및 특급 시약을 구입하여 사용하였다. 본 연구에 사용한 병꽃나무 꽃은 소망농원(Seoul, Korea)에서 묘목을 구입하여 심고 키워 개화기에 채집하여 사용하였다.
시료 제조
본 연구에 사용한 병꽃나무 꽃 에탄올 추출물은 묘목에서 꽃잎만 채취하여 음건한 후 무게의 10배량(w/v)의 70% 에탄 올을 가하여 3시간씩 3회 환류 추출 후 냉침하여 Whatman 여과지(No. 5)로 여과하였다. 여과된 추출물을 40℃ 이하에서 감압농축 및 동결 건조한 후 DMSO에 녹여 시료(WS-E)로 사 용하였다.
세포배양
본 연구에 사용된 마우스 대식세포주인 RAW 264.7세포는 한국세포주은행(Seoul, Korea)에서 분양 받아 사용하였다. 세 포는 10% fetal bovine serum (FBS)과 1% antibiotics (penicil- lin-streptomycin)을 첨가한 DMEM 배지를 사용하여 37℃, 5% CO
2조건에서 배양하였다.
총 폴리페놀 함량 측정
총 폴리페놀 함량은 Folin-Denis 방법[5]을 변형하여 측정 하였다. 즉, 적당하게 희석된 시료 1 ml에 95% ethanol 1 ml와 증류수 5 ml를 첨가하고 1 N Folin-Ciocalteu’s reagent 0.5 ml를 넣어 잘 섞어 주었다. 5분간 방치한 후, 10% Na
2CO
31
ml를 가한 후, 725 nm에서 1시간 이내에 흡광도(UV/Vis spectrophotometer, Thermo Scientific, G10S, MA, USA)를 측 정하였다. Tannic acid를 표준물질로 사용하였으며 총 폴리페 놀 함량은 시료 1 ml 중의 μg tannic acid로 나타내었다.
총 플라보노이드 함량 측정
총 플라보노이드 함량은 Moreno 등[16]의 방법에 의해 측 정하였다. 각 시료 0.5 ml에 10% aluminum nitrate 0.1 ml, 1 M potassium acetate 0.1 ml 및 ethanol 4.3 ml를 가하여 혼합하고 실온에서 40분 동안 정치한 뒤 415 nm에서 흡광도를 측정하였다. Quercetin을 표준물질로 사용하였으며 총 플라보 노이드 함량은 시료 1 ml 중의 μg quercetin으로 나타내었다.
DPPH 라디칼 소거능 측정
DPPH 라디칼에 대한 소거활성은 Blois의 방법[2]을 변형하 여 측정하였다. 각 시료 20 μl에 0.1 mM DPPH 180 μl를 넣고 vortex한 후, 30분 동안 방치한 다음 microplate reader (Tecan, Sunrise, NC, USA)를 사용하여 560 nm에서 흡광도를 측정하 였다. 시료를 첨가하지 않은 대조군과 비교하여 시료의 DPPH 라디칼 소거능을 구하였다.
Superoxide 음이온 라디칼 소거능 측정
Superoxide 음이온 라디칼 소거활성은 Nishikimi의 방법 [19]을 변형하여 각 시료의 xanthine-xanthine oxidase system 에 의해 생성된 supreroxide 음이온 라디칼을 소거하는 효과 를 측정하였다. 3 mM xanthine, 0.6 mM NBT, 15 mM EDTA 및 50 mM potassium phosphate buffer (pH 7.4)를 혼합한 혼합액에 시료 30 μl를 첨가하고 0.1 U xanthine oxidase를 50 μl 가하여 5분간 전 반응(preincubation)시킨 후, 37℃에서 20분간 반응시켰다. 반응이 끝난 후 microplate reader (Tecan) 을 사용하여 560 nm에서 흡광도를 측정하였다. 시료를 첨가하 지 않은 대조군과 비교하여 시료의 superoxide 음이온 라디칼 소거능을 구하였다.
세포독성 측정
세포독성은 MTT 시약을 이용하여 세포 생존율을 측정하는
Mosmann의 방법 [17]을 변형하여 실시하였다. RAW 264.7
세포를 5×10
4cells/well 농도로 96-well plate에 분주한 후 시
료를 0~100 μg/ml의 농도가 되도록 세포배양배지로 희석하
여 첨가하고 12시간 전처리 하였다. 그 후 LPS (E. coli serotype
055:B5)를 100 ng/ml로 세포에 첨가하여 24시간 동안 염증반
응을 유도하였다. MTT 용액(5 μg/ml)을 첨가하고 4시간 후
원심분리하여 상등액을 제거하고 100 μl acid-isopropanol
(0.04 N HCl in isopropanol)을 첨가한 후, 생성된 formazan이
용출되도록 하여 microplate reader (Tecan)로 560 nm에서 흡
광도를 측정하였다. 생존율은 대조군에 대한 흡광도의 차를
Table 1. The total polyphenol and flavonoid contents of extracts from the flowers of Weigela subsessilis (WS-E) Sample Total polyphenol
(μg/ml)
1)Total flavonoid (μg/ml)
2)WS-E 719.19±0.04
3)644.89±0.02
1)
Total polyphenol content (μg/ml) was equivalent as tannic acid as standard.
2)
Total flavonoid content (μg/ml) was equivalent as quercetin as standard.
3)
Each value is mean of triplicate determinations±SD.
백분율로 표시하여 비교 분석하였다.
Nitric oxide (NO) 및 cytokine 생성량 측정
NO 생성량 측정은 Lee 등[14]의 방법에 따라 다음과 같이 측정하였다. RAW 264.7 세포(5×10
4cells/well)를 96-well plate에 분주한 후 시료를 농도별(0.16, 0.8, 4, 20, 100 μg/ml)로 12시간 전 처리한 다음, LPS로 24시간 자극하여 염증을 유발 하였다. 배양액 중에 분비된 NO의 농도는 NO 정량 kit (iNtRON Biotechnology, Seongnam, Korea)를 이용하여 측정 하였다. 또한 세포배양액 중에 분비된 cytokine의 생성량은 enzyme linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN, USA)를 이용하여 측정하였다.
Western bot
iNOS의 세포 내 발현은 Western blot으로 분석하였다. 시 료를 농도별로 전 처리한 RAW 264.7 세포(1×10
6cells/well)에 LPS (100 ng/ml)로 자극한 후 세포를 회수하여 RIPA buffer (protease and phosphatase inhibitor cocktail; Thermo Scien- tific, Waltham, MA, USA)로 4℃에서 30분간 처리하였다. LPS 자극은 iNOS 분석에는 6시간, mitogen-activated protein kin- ase (MAPK)와 nuclear factor (NF)-κB 분석에는 15분간 처리 하였다. 원심분리 한 후 상층액을 얻고 BCA protein assay kit (Bio-Rad, Hercules, CA, USA)를 이용하여 단백질 양을 정량 하였다. 그 후 10% sodium dodecyl sulfate (SDS)-polyacryl- amide gel electrophoresis (PAGE) gel에 각 lane 당 30 μg의 세포 단백질을 넣어 전기영동 하고, gel 내의 단백질을 poly- vinylidene fluoride membrane filter (PVDF, Bio-Rad) 막에 transfer 하였다. PVDF 막을 5% skim milk 용액으로 blocking 하고, IκB, p38, extracellular signal regulated kinase (ERK), c-Jun-N-terminal kinase (JNK), phosphorylated IκB (p-IκB), phosphorylated p38 (p-p38), phosphorylated ERK (p-ERK), phosphorylated JNK (p-JNK) 각각에 대한 항체(R&D Systems)를 실온에서 2시간 반응시켰다. 그 후 각 항체에 대한 2차 항체인 horseradish peroxidase (HRP)-conjugated anti- body (R&D Systems)를 반응시킨 후 ECL kit (Amersham Pharmacia Biotech, Amersham, Buckinghamshire, UK)를 이 용하여 각 band를 발현시켜 관찰하였다. 각 band의 발현은 densitometer를 이용하여 분석하였다.
통계처리
모든 결과는 평균±표준편차로 나타내었으며, 유의성 검사 는 SPSS TM version 20.0 (SPSS Inc., Chicago, USA)를 이용하 여 one-way ANOVA를 실시하였다. 대조군에 대한 시료 처리 군의 통계적 유의성은 Duncan's multiple range test로 검증하 였다.
결과 및 고찰
총 폴리페놀 및 플라보노이드 함량 변화
폴리페놀계 화합물은 대부분 식물들의 항산화능을 부여하 는 2차 대사산물로 식물계에 8,000여 개의 구조를 가진 성분으 로 존재하며 페놀성 화합물이라고도 한다. 한 분자 내에 2개 이상의 페놀성 수산기가 효소 단백질과 같은 거대 분자들과 결합하는 성질이 있기 때문에 항산화 작용, 항암 등의 다양한 생리활성을 가지는 것으로 알려져 있다[7]. 또한, 주된 식물계 폴리페놀 물질인 플라보노이드는 항산화, 항균, 충치예방, 항 암 및 항염증 등이 보고되어 있다[3].
WS-E에 존재하는 총 폴리페놀 및 플라보노이드 함량을 측 정하기 위하여 표준물질로 각각 tannic acid와 quercetin을 사 용하였다. 그 결과 Table 1에서와 같이 WS-E의 총 폴리페놀 함량은 719.19±0.04 μg TAE/ml로 나타났다. 또한, 총 플라보 노이드 함량은 644.87±0.02 μg QE/ml로 나타났다. Jung 등[8]
의 보고에 의하면 소태나무 잎 추출물의 폴리페놀과 플라보노 이드 함량이 각각 367.52 μg/mg과 46.41 μg/mg인 것에 비해 본 연구의 WS-E의 폴리페놀과 플라보노이드 함량은 매우 높 은 것으로 사료된다.
항산화 활성
천연 유래 항산화제의 작용 기작은 유리기와 반응하는 것으 로 라디칼의 소거 활성은 자유 라디칼에 전자를 공여하여 항 산화 효과나 인체에서 노화를 억제하는 척도로 활용되고 있다 [1]. WS-E의 항산화 활성은 DPPH 라디칼 소거능과 super- oxide 음이온 라디칼 소거능을 가지고 측정하였다.
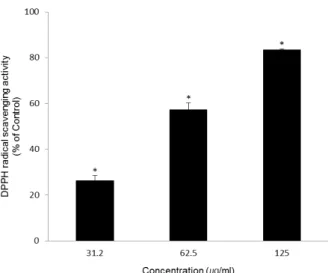
DPPH 라디칼 소거능은 안정한 라디칼인 DPPH를 소거시 키는 항산화 물질의 활성을 측정하는 것으로 짙은 자색을 띄 는 DPPH 라디칼이 방향족 아민류 등에 의해 환원되어 색이 탈색되는 것을 이용하여 항산화 물질을 검색하는 데 이용되고 있다[13]. WS-E의 DPPH 라디칼 소거능을 측정한 결과 Fig.
1에 나타낸 바와 같이 WS-E의 DPPH 라디칼 소거활성은 31.2, 62.5, 125 μg/ml에서 각각 26.27±2.31%, 57.25±3.14%, 83.44±
0.41%로 나타났으며, 농도 의존적으로 활성이 증가하고 있음
을 알 수 있었다. Kang [9]이 보고한 합환피(Albizziae cortex)
Fig. 1. Scavenging activity of WS-E on DPPH radicals. All values were expressed as the average of triplicate samples with SD.
*p<0.05 compared with control.
Fig. 2. Scavenging activity of WS-E on superoxide anion radicals.
All values were expressed as the average of triplicate samples with SD.
*p<0.05 compared with control.
Fig. 3. Effect of WS-E on cytotoxicity in RAW 264.7 macrophages.
RAW 264.7 cells (5×10
4cells/well) were treated with the indicated doses of WS-E for 12 hr prior to 24 hr in- cubation with LPS (100 ng/ml). All values were ex- pressed as the average of triplicate samples with SD.
추출물이 400 μg/ml에서 25.80±2.80%의 소거활성을 보인 것 과 비교할 때 WS-E의 DPPH 라디칼 소거능이 우수함을 알 수 있다.
Superoxide 음이온 라디칼 소거능은 xanthine-xanthine ox- idase 반응으로 uric acid가 생성되는 과정에 생성되는 super- oxide 음이온 라디칼을 WS-E와 반응시켜 NBT로 확인하는 방 법이다[12]. WS-E의 xanthine oxidase에 의해 생성되는 super- oxide 음이온 라디칼 소거능을 측정한 결과 Fig. 2에 나타낸 바와 같이 WS-E는 투여 농도 의존적으로 superoxide 음이온 라디칼 소거작용을 나타내었다. 즉, 31.2, 62.5, 125 μg/ml의 농도에서 각각 53.12±10.48%, 65.61±9.31%, 76.11±5.22%의 소 거능을 보였다. Rhim 등[23]의 연구에서 소리쟁이(Rumex
crispus) 추출물의 항산화 활성 연구 결과 0.1, 0.5, 1 mg/ml농도에서 각각 26.06%, 33.13%, 43.17%의 소거효과를 보였다 는 보고와 비교할 때 WS-E의 superoxide 음이온 라디칼 소거 능은 소리쟁이 추출물 보다 우수한 것으로 사료된다.
세포독성
MTT assay를 이용하여 WS-E의 RAW 264.7 대식세포에 대 한 세포독성을 측정한 결과 100 μg/ml의 농도까지 세포의 증 식에 별다른 영향을 주지 않는 안전한 농도인 것으로 확인되 었다(Fig. 3). 따라서 전혀 독성을 나타내지 않는 안전한 농도 인 100 μg/ml이하의 농도로 WS-E를 처리하여 다음 실험을 수행하였다.
NO 생성 억제
NO는 L-arginine을 기질로 하여 NO 합성효소에 의해 생성
되는 무기 유리체로 지속적인 염증의 진행에 의한 종양 형성
촉진, 면역 반응, 혈관 이완 등 여러 생물학적인 과정에 관여하
는 인자 중의 하나로 인식되고 있다[10]. WS-E의 항염증 활성
을 측정하기 위하여 먼저 LPS로 전 처리한 RAW 264.7 대식세
포에 WS-E를 농도별로 처리한 후 염증매개인자인 NO 생성
정도를 정량하였다. 그 결과 Fig. 4에 나타낸 바와 같이 WS-E
를 농도별로(0.16, 0.8, 4, 20, 100 μg/ml) 처리하였을 때 8.98,
8.15, 7.42, 6.48, 4.71 μM로 투여 농도 의존적으로 NO의 생성
을 억제하였다(p<0.05). 또한 염증반응의 자극에 의해 유도되
는 NO를 생합성 하는 효소로 알려진 iNOS의 세포내 발현량
을 Western blot으로 측정하여 NO 분비 억제에 관한 작용기
전을 확인하였다. 그 결과 WS-E를 전 처리한 경우에 있어서
iNOS의 세포 내 발현이 현저하게 억제되었으며, 이 억제효과
는 WS-E의 농도에 의존하는 것으로 나타났다(Fig. 5). 또한
Fig. 4. Inhibitory effect of WS-E on NO production from LPS- stimulated RAW 264.7 macrophages. The level of NO was determined by NO detection kit. All values were expressed as the average of triplicate samples with SD.
*
p<0.05 compared with control.
A
B
Fig. 5. Effect of WS-E on iNOS expression in LPS-stimulated RAW 264.7 macrophages. Total cellular proteins were obtained at 6 hr after LPS stimulation for detection of iNOS. The level of expression of iNOS was measured by Western blot (A). Actin was used as an internal con- trol in a densitometric analysis (B). This result is the representative of almost same results from two in- dependent experiments.
A
B
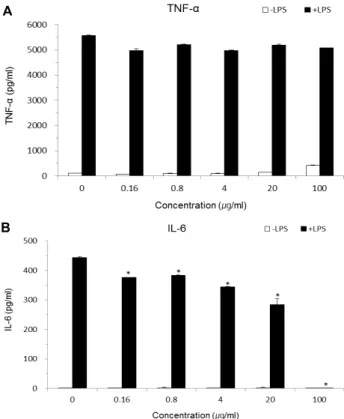
Fig. 6. Inhibitory effect of WS-E on TNF-α and IL-6 production from LPS-stimulated RAW 264.7 macrophages. The level of TNF-α (A) and IL-6 (B) in the supernatants of RAW 264.7 cells was determined by ELISA kits. All values were expressed as the average of triplicate samples with SD.
*p<0.05 compared with control.
Western blot에 의한 결과를 정량적으로 관찰하기 위해 densi- tometer를 이용하여 분석한 결과에서도 iNOS의 발현이 현저 하게 억제되는 것으로 관찰되었다. Kim 등[11]은 NO 저해 활성이 높게 나타난 왕쥐똥나무(Ligustrum ovalifolium) 잎 에 탄올 추출물이 iNOS의 발현을 효과적으로 저해한다고 보고하 였으며, 이는 WS-E의 NO 생성 억제와 iNOS 발현 저해양상과 유사한 결과로 사료된다. 따라서 LPS로 자극한 대식세포에
대한 WS-E의 NO 생성 억제는 NO 생합성 효소인 iNOS의 세포 내 발현 억제에 의한 것으로 사료된다.
TNF-α 및 IL-6 측정
주로 단핵세포와 대식세포에서 생산되는 TNF-α는 면역 반
응과 염증 반응을 유도하는 염증유도 매개 cytokine으로 세포
의 성장과 분화, apoptosis, necrosis 등의 기능에 관여하며,
또한 대식세포와 T-세포에 의해서 생산되는 IL-6는 inflamma-
tory와 pro-inflammatory cytokine으로 알려져 있다[27]. 본 연
구에서는 염증반응에서 대식세포로부터 분비되는 중요한 in-
flammatory cytokine으로 알려진 TNF-α와 IL-6의 분비에 대
한 WS-E의 억제활성을 조사하였다. 그 결과 LPS로 자극된 대
식세포로부터 분비되는 TNF-α에 대해서는 WS-E 전 처리에
의한 억제효과가 관찰되지 않았다(Fig. 6). WS-E가 NO의 분비
를 억제하는 활성을 보임에도 불구하고 TNF-α의 분비를 억제
하지 않는 이유를 분석하기 위하여 WS-E가 대식세포에 대해
TNF-α의 생성을 유도하는 가를 조사하였다. 그 결과 비교적
높은 농도인 100 μg/ml의 농도에서부터 WS-E는 대식세포로
부터 TNF-α의 분비를 유도하는 것으로 관찰되었다(data not
shown). 결과적으로 WS-E의 항염증 활성에서 TNF-α의 분비
A
B C
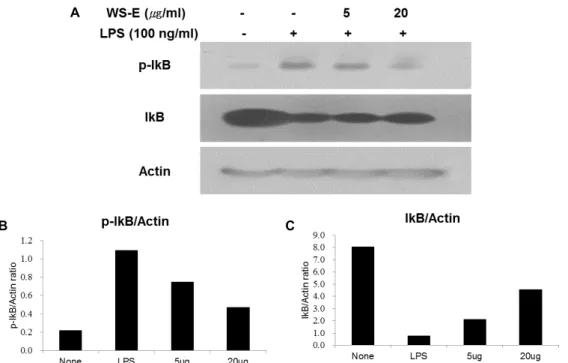
Fig. 8. Inhibitory effect of NF-κB activation by WS-E in LPS-stimulated RAW 264.7 macrophages. NF-κB activation in LPS-stimulated RAW 264.7 cells was measured by IκB phosphorylation. Equal amounts of total protein were resolved by SDS-PAGE (A).
Actin was used as an internal control in a densitometric analysis (B; pIκB/Actin, C; IκB/Actin). This result is the representative of almost same results from two independent experiments.
Fig. 7. Effect of WS-E on MAPK activation in LPS-stimulated RAW 264.7 macrophages. Total cellular proteins were harvested at 15 min after LPS stimulation. Phosphoryla- tion of p38, ERK, and JNK was determined by Western blot. This result is the representative of almost same re- sults from two independent experiments.
억제가 인정되지 않았던 것은 WS-E가 단독으로 TNF-α를 유 도하는 것과 관련이 있는 것으로 사료된다.
한편, 또 다른 inflammatory cytokine인 IL-6의 경우에는 NO에서와 동일하게 WS-E의 전 처리에 의해 IL-6의 분비가
유의하게 억제되는 것을 확인하였다(Fig. 6). 이상의 결과로부 터 WS-E는 LPS에 의한 대식세포의 염증반응에 있어서 NO와 inflammatory cytokine인 IL-6의 분비를 억제하는 활성이 인 정되었으나 TNF-α의 분비에 대해서는 억제효과가 없는 것으 로 확인되었다.
MAPK 및 NF-κB 억제활성
LPS 자극에 의해 대식세포가 염증작용을 일으킬 때에는 다 양한 세포 내 신호전달을 통해 염증이 유도된다. 특히 p38, JNK 및 ERK와 같은 MAPK는 이러한 염증반응에 있어서 매우 중요한 신호전달 분자로서 이들 분자의 인산화에 의해 염증반 응이 유발되는 것으로 알려져 있다[24]. 따라서 WS-E의 항염 증 활성이 MAPK의 조절여부에 의해 나타나는 지 알아보기 위하여 평가한 결과, Fig. 7에 나타난 바와 같이 LPS 자극에 의한 p38, ERK 및 JNK 등의 MAPK 활성을 억제하지 못하는 것으로 나타났다.
한편 NF-κB는 염증반응에 있어서 iNOS 및 COX-2의 발현
을 조절하는 전사 조절인자로 LPS 자극에 의한 대식세포의
염증유발 기전에는 NF-κB의 활성화에 의한 핵 내 전위가 중요
하다. 평상시 NF-κB는 세포질에서 IκB와 결합되어 핵 내 전위
가 불가능한 불활성화 상태로 존재하나 염증성 자극에 의하여
IκB kinase가 활성화 되어 IκB가 인산화 될 경우 IκB로부터
유리되어 핵 내로 이동하여 iNOS, COX-2 및 inflammatory
cytokine의 합성을 촉진한다고 알려져 있다[22]. 따라서 WS-E
의 항염증 활성이 NF-κB 활성화 억제에 의한 것인지를 알아보 기 위하여 WS-E를 전 처리 하고, LPS로 자극한 대식세포에서 IκB의 인산화에 대한 영향을 측정하였다. 그 결과 Fig. 8에 나 타낸 바와 같이 WS-E는 LPS에 의한 IκB의 인산화를 농도 의존 적으로 억제하는 것으로 확인되었다.
이상의 결과를 종합해 보면 병꽃나무 꽃 에탄올 추출물인 WS-E의 항염증 활성은 MAPK 억제보다는 NF-κB의 활성 억 제, 그리고 iNOS의 세포 내 발현 억제를 통한 NO 생성의 저 해, inflammatory cytokine인 IL-6의 억제 등과 같은 일련의 기전을 통해 일어나는 것으로 추정되었다. 향후 TNF-α의 분비 에 관한 활성에 대해 면밀한 검토가 필요할 것으로 사료된다.
감사의 글
본 연구는 지식경제부와 한국산업기술진흥원의 지역산업 기술개발사업(No. A001100081)으로 수행된 연구결과입니다.
또한 본 연구는 부분적으로 2014년도 중소기업청 기술개발사 업(No. C0248870)의 지원으로 수행되었습니다.
References