․교신저자: 신현철 경북 포항시 남구 대잠동 907-8번지 대구한의대학교 부속 포항한방병원 3내과 TEL: 054-281-0055 FAX: 054-281-7464 E-mail: [email protected]
Mouse의 신장상피세포에서 敗醬추출물이 산화 스트레스 및 NF-κB signaling에 미치는 영향
김현영, 장수영, 최규호, 신현철 대구한의대학교 한의과대학 내과학교실
The Effect of Patriniae Radix on the Oxidative Stress and the NF-κB Signaling in Mouse LLC-PK
1Cell
Hyun-young Kim, Soo-young Jang, Gyu-ho Choi, Hyeon-cheol Shin Dept. of Internal Medicine, College of Oriental Medicine, Dae-gu Haany University ABSTRACT
Objectives :The aims of this study were to investigate the cytoprotective, antioxidative and inflammation genes inhibitory effects of Patriniae Radix on the mouse LLC-PK1cells (renal epithelial cells).
Methods :The cytoprotective effect of Patriniae Radix was evaluated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The antioxidative effect was measured in terms of generation amount of superoxide anion radical (․
O2-) by 2',7'-dichlorodihydrofluorescein diacetate (DCFDA), nitric oxide (NO) by 4,5-diaminofluorescein (DAF-2), peroxynitrite (ONOO-) by dihyldrorhodamine 123 (DHR 123) and prostaglandin E2 (PGE2) by PGE2 immunoassay on H2O2-treated LLC-PK1 cells. For measuring of inflammation genes inhibitory effects, western blot was performed to detect IKK-α, phospho-IκB-α, NF-κB (p50, p65), COX-2, iNOS, IL-1β and VCAM-1 protein level in cytosol fractions from LLC-PK1cells.
Results :Patriniae Radix extract reduced the H2O2-induced cell death and inhibited the amount of H2O2-induced ․O2-, NO, ONOO-, PGE2generation dose-dependently on the mouse LLC-PK1cells in vitro. Also Patriniae Radix extract inhibited the expression of IKK-α, phospho-IκB-α, COX-2, iNOS, IL-1β and VCAM-1 genes dose-dependently by means of decreasing activation of NF-κB.
Conclusions :According to above results, it was identified that Patriniae Radix had the cytoprotective, antioxidative and inflammation genes inhibitory effects. So it was suggested that Patriniae Radix would be effective to the treatment for the inflammatory process and inflammation-related diseases.
Key words :
Patriniae Radix, ROS, ONOO-, NF-κB, COX-2Ⅰ. 서 론
산화 스트레스는 활성산소종 (reactive oxygen
species, ROS)과 활성질소종 (reactive nitrogen species,
RNS)의 발생 및 항산화 방어계 (antioxydant defence
system)간에 심각한 불균형이 초래되는 상황을 말
한다. ROS와 RNS에는 superoxide anion radial
(․O
2-), hydrogen peroxide (H
2O
2), hydroxyl radical
(․OH) 등과 nitric oxide (NO), nitrogen dioxide
(NO
2), nitrous acid (HNO
2), peroxynitrite (ONOO
-)
등이 있으며, 이러한 free radical은 생체막의 주성 분인 인지질을 과산화하여 막기능을 저하시키거나 DNA 과산화를 통해 세포 손상에 관여한다
1,2.
Nuclear transcription factor-kappa B (NF-κB) 는 Sen과 Baltimore
3에 의해 발견된 염증반응 및 세포사에 중요한 전사인자이다. 자극이 없는 상태 에서 NF-κB는 세포질 내에 Inhibitor κB (IκB)와 결합하여 IκB, p50, p65로 구성된 heterotrimer의 상태로 존재하는데, 산화 스트레스 같은 외부자극 이 주어지면 IκB가 인산화되면서 그 결합이 파괴 되어 활성화된 p50, p65가 핵 내로 이동하게 된다.
이렇게 활성화된 NF-κB는 표적 유전자의 promoter regions에 있는 κB 결합자리에 결합하여 cyclooxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS), Tumor necrosis factor-α (TNFα), Interleukin-1β (IL-1β), vascular cell adhesion molecule-1 (VCAM-1) 등 여러 염증 매개 물질들의 전사를 촉진한다
4,5.
敗醬은 마타리과 (敗醬科 : Valerianaceae)에 속 한 多年生 草本인 뚝갈 (白花敗醬) Patrinia villosa (Thunb.) Juss와 마타리 (黃花敗醬) P. scabiosaefolia Fisch.의 뿌리 또는 帶根全草를 건조한 것
6,7이다.
<神農本草經>
8에 “主暴熱火瘡 赤氣 疥瘙疽痔 馬鞍 熱氣”라고 처음 수록된 이래 淸熱解毒, 消腫排膿, 祛瘀止痛 등의 효능으로 腸癰, 下痢, 赤白帶下, 産 後瘀滯腹痛, 目赤腫痛, 癰腫疥癬 등의 치료에 상용 되고 있다
6. 敗醬과 관련한 실험연구에 의하면 급 성췌장염 억제 효과
9, 식중독 유발 세균의 증식억 제 효과
10, 항염증 작용
11, 항암작용
12및 간기능 회 복 효과
13등이 보고되었다.
본 연구에서는 敗醬의 이런 효능을 참고하여 산 화 스트레스와 관련한 항산화, 항염증 효과를 규명 하고자 하였다. 이에 mouse의 신장상피세포를 이 용하여 敗醬의 세포 독성 및 H
2O
2에 의한 세포사 보호 효과, ․O
2-, NO, ONOO
-및 prostaglandin E
2(PGE
2) 생성과 COX-2, iNOS, IL-1β, VCAM-1의 발현 및 NF-κB signaling pathway 관련 염증성 단 백질에 미치는 영향을 평가하였다.
Ⅱ. 재료 및 방법
1. 재 료 1) 약 재
敗醬根 (Patriniae Radix)은 대구한의대학교 부 속한방병원 약제과에서 구입하고 정선하여 사용하 였다.
2) 시약 및 기기
시약으로는 Dulbecco's Modifide Eagle Medium Nutrient Mixture F-12 (DMEM/F12), fetal bovine serum (FBS), streptomycin-penicillin 등의 세포배 양용 시약들은 Gibco BRL (Grand Island, USA) 에서, dihydrorhodamine 123 (DHR 123)과 2’,7’- dichloro dihydrofluorescein diacetate (DCFDA)는 molecular Probes (Eugene, OR, USA)에서, carboxy -PTIO sodium salt는 Calbiochem (EMD Biosciences Inc., USA)에서, 4,5-diaminofluorescein (DAF-2)은 Dai ichi Pure Chemical Co. (Tokyo, Japan)에서, sodium dodesyl sulfate (SDS), acrylamide는 Bio-Rad (Hercules, USA)에서, NP-40, CAPS, Tris base, ammonium persulfate, ponceau, glycerol, protease inhibitors, bovine serum albumin은 Sigma Chemical Co. (St. Louis, MO, USA)에서, 1차 항체인 IKK- α , phospho-IκB-α는 Cell Signaling Technology (Cell Signaling, USA)에서, p50, p65, iNOS, COX-2, VCAM-1, IL-1β, β-actin, histone H1과 2차 항체인 anti-mouse, anti-rabbit, anti-goat IgG는 Santa Cruz Biotechnology (Santa Cruz, USA)에서 구입하였고, sodium chloride, hydrogen peroxide, ethanol, methanol 및 기타 시약 은 시중에서 특급품을 구입하여 사용하였다. 기기 는 GENios (GENios-basic, Tecan, Austria), UV-VIS spectrophotometer (UV-2401PC, Shimadzu Co., Japan) 제품을 각각 구입하여 사용하였다.
3) 세 포
Mouse의 신장상피세포인 LLC-PK
1세포 (Renal
epithelial cells)는 경북대학교 의과대학 해부학교실
에서 분양받아 사용하였다.
2. 방 법
1) 검액의 조제
잘게 분쇄한 敗醬根 300 g에 3배 량의 95% methanol 을 가하고 60℃에서 중탕으로 24시간씩 3회 반복 추출하여 추출액을 얻었다. 이 추출액을 실온으로 냉각시키고 여지로 여과한 다음 여액을 회전 감압 농축기를 사용하여 건조시켜 추출물 61.61 g (수율 20.54%)을 얻어 실험에 필요한 농도로 희석하여 사용하였다.
2) 세포 배양
LLC-PK
1세포를 100 ㎜ dish (Corning Co., New York, USA)에 배양하였다. 이때 5% fetal bovine serum, L-glutamine (5.84 ㎍/㎖)과 L-sodium bicarbonate (2.438 g/㎖)를 함유한 DMEM/F12 배지에 penicillin -streptomycin (100 U/㎖)을 첨가하여 사용하였고, 37℃, 5% CO
2incubator에서 배양하였으며, 2일에 1회씩 subculture하여 세포주를 유지하였다.
3) MTT assay
敗醬의 세포 독성 및 H
2O
2로 유발시킨 세포사에 대한 敗醬의 세포 보호 효과를 알아보기 위해, Tada 등의 방법
14으로 측정하였다. 노란색의 3- (4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) 용액은 살아 있는 세포에서는 mitochondria의 succinyl dehydrogenase에 의해 비 수용성인 보라색의 formazan으로 환원된다. 이 formazan 의 발색 정도를 spectrophotometer로 측정하여 세 포의 생존율을 측정함으로써 敗醬의 독성 및 세포 보호 효과를 측정할 수 있다. LLC-PK
1세포를 96 well plate에 1×10
4/well로 접종하여 24시간 배양하 고 serum free 배지로 바꿔준 후, 敗醬 추출물을 농 도별로 가하여 1시간 전처리한 다음 敗醬의 세포 독성을 측정하였다. LLC-PK
1세포를 96 well plate 에 1×10
4/well로 접종하여 24시간 배양하여 敗醬 추출물을 1시간 전처리하고 H
2O
2를 최종 농도 100 mM이 되도록 가하여 5시간 더 배양한 후 敗醬의 세포 보호 효과를 측정하였다. 측정시에는 배지를 제거하고 0.5 ㎎/㎖ MTT 용액을 세포에 가하여 4
시간 추가 배양하고 solublization 용액 (dimethyl sulfoxide:ethanol, 1:1)을 가하여 24시간 배양한 후, 595 nm에서 발색 정도를 측정하였다.
4) LLC-PK
1세포에서 ․O
2-, NO 및 ONOO
-소 거능 측정
LLC-PK
1세포를 96 well plate에 1×10
4/well로 분주하여 하루 동안 배양한 다음 敗醬 추출물을 농도별로 전처리한 후 1시간 동안 배양하고, 각각 H
2O
2100 mM을 처리하여 5시간 동안 배양한 후 각 소거능을 측정하였다.
․O
2-소거능은 DCFDA assay
15로 측정하였다.
지용성의 DCFDA가 esterase 또는 산화적 가수분 해를 받아 비형광성인 DCFH로 탈아세틸화되며, DCFH는 활성산소에 의해 산화되어 강한 형광을 나타내는 2’,7’-dichlorofluorescein (DCF)이 된다.
따라서 96 well microplate의 배지를 제거하고 각 well당 free media 200 ㎕와 50 mM phosphate buffer (pH 7.4)로 희석한 0.125 mM DCFDA 50
㎕를 첨가한 후 여기파장 485 nm와 방출파장 535 nm에서 5분 간격으로 7회 측정하였다.
NO 소거능은 Nagata 등의 방법
16으로 측정하였 다. 특이적인 NO의 indicator인 4,5-diamino fluorescein (DAF-2)은 2개의 아미노기 사이에 NO를 포집하 여, 490-495 nm의 여기파장에서 green의 형광을 방 출하는 triazolofluorescein을 생성하며 형광의 세기 는 DAF-2에 의해 포집된 NO의 양에 의존한다. 따 라서 96 well microplate의 배지를 제거하고 각 well 당 free media 150 ㎕와 Dimethyl sulfoxide 414.5
㎕에 DAF-2 1 ㎎이 녹아 있는 것을 50 mM phosphate buffer (pH 7.4)로 1:400배로 희석한 DAF-2 50 ㎕ 를 첨가한 후 여기파장 485 nm와 방출파장 535 nm에서 3분 간격으로 7회 측정하였다.
ONOO
-소거능은 Kooy 등의 방법
17에 의해 측
정하였는데 96 well microplate의 배지를 제거하고
각 well당 free media 20 ㎕와 90 mM NaCl, 5 mM
KCl 및 100 μM diethylenetriaminepenta acetic acid
와 10 μM DHR 123을 함유하는 sodium phosphate
완충액 (pH 7.4) 180 ㎕를 가하여 5분간 방치한 후 여기파장 485 nm와 방출파장 535 nm에서 측정하 였다.
5) Prostaglandin E
2측정
세포내에서 생성되는 염증물질인 prostaglandin E
2(PGE
2)의 양은 배지를 사용하여 PGE
2immunoassay kit (R&D SYSTEMS)의 실험 방법에 따라 측정 하였다.
6) Western blot analysis
LLC-PK
1세포를 100 ㎜ dish에 24시간 배양한 후 敗醬 추출물을 농도별로 1시간 전처리하고 H
2O
2100 mM을 가하여 5시간 더 배양한 후 PBS로 세 척하고 scraper로 모은 LLC-PK
1세포에서 cytosol, nuclear 단백질을 분리하여 염증인자의 단백질 발 현을 관찰하였다. 동일 량의 단백질을 sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) 로 분리시킨 후, nitrocellulose membrane에 단백질 들을 전이시켜 주었다. 이 membrane을 분리한 후 5% skim milk로 단백질이 결합하지 않은 부분의 비특이적 antibody 결합을 차단시키고 각 타깃 단 백질의 1차 antibody를 2시간 이상 처리하였다. 이 membrane을 0.1% Tween 20을 함유한 TBST로 60 분간 세척한 다음 2차 antibody를 2시간 이상 반응 시키고, TBST로 60분간 세척한 다음 membrane에 ECL solution을 반응시켜 발생된 형광을 X-ray film에 감광시켰다.
7) 단백질 정량
단백질의 정량은 Lowry 등의 방법
18과 Bradford protein assay kit를 사용하여 정량하였다.
8) 통계 처리
실험 성적의 분석은 각 실험군 간의 평균치와 평균오차로 표시하고 각 실험군 간의 유의성 검정 은 student's t -test를 이용하여 통계 처리하였다.
Ⅲ. 결 과
1. 세포 독성
敗醬을 농도별 (25, 50, 100, 200, 400, 800, 1000
㎍/㎖)로 처리하였을 때 각각의 농도에서 98.24, 96.34, 87.47, 85.66, 61.95, 42.33, 31.29%의 생존율을 나타내어, 200 ㎍/㎖까지는 세포 독성이 거의 없는 것으로 나타났다(Fig. 1).
Fig. 1. Cytotoxic Effect of Patriniae Radix in LLC-PK
1Cells.
LLC-PK1cells were exposed to Patriniae Radix (25, 50, 100, 200, 400, 800, 1000 ㎍/㎖). After 24-hr exposure, the cell viability was evaluated by MTT assay. Each value is the mean±S.D.
Significantly different from the normal (** : p <
0.01).
2. H
2O
2처리에 대한 세포 보호 효과
H
2O
2를 처리하였을 때의 세포 생존율은 58.79%
였으나, 敗醬 추출물을 25, 50, 100, 200, 400, 800,
1000 ㎍/㎖로 전처리한 세포의 생존율은 각각
60.33, 73.24, 79.45, 83.37, 64.22, 55.38, 39.85%로 나
타나 200 ㎍/㎖까지는 농도 의존적으로 H
2O
2에 의
한 세포사를 저해하는 것으로 나타났다(Fig. 2).
Fig. 2. Cytoprotective Effect of Patriniae Radix on H
2O
2-induced Cell Death in LLC-PK
1Cells.
LLC-PK1cells were exposed to H2O2(100 mM) in the presence and absence Patriniae Radix (25, 50, 100, 200, 400, 800, 1000 ㎍/㎖). After 5hr exposure, the cell viability was evaluated by MTT assay. Each value is the mean±S.D.
Significantly different from the normal (** : p
< 0.01, * : p < 0.05). : normal, : control, : experimental
3. H
2O
2처리에 의한 ․O
2-, NO 및 ONOO
-생성에 미치는 영향
․O
2-는 정상 세포에서 126.38±6.88 fluorescence/
min인데 비하여 H
2O
2처리에 의해 191.28±10.01 fluorescence/min으로 생성이 증가하였으나, 敗醬 추출물을 전처리한 경우에 농도 의존적으로 감소 하여 50 ㎍/㎖ 농도에서 166.21±8.88 fluorescence/min, 100 ㎍/㎖ 농도에서 149.38±8.13 fluorescence/min, 200 ㎍/㎖ 농도에서 144.64±8.11 fluorescence/min으 로 유의성 있게 감소하는 것으로 나타났다(Fig. 3).
NO는 정상 세포에서 55.48±2.04 fluorescence/min 인데 비하여 H
2O
2처리에 의해 92.57±3.03 fluorescence/
min으로 생성이 증가하였으나, 敗醬 추출물을 전 처리한 경우에 농도 의존적으로 감소하여 50 ㎍/㎖
농도에서 72.34±3.93 fluorescence/min, 100 ㎍/㎖
농도에서 68.26±2.11 fluorescence/min, 200 ㎍/㎖
농도에서 64.27±3.44 fluorescence/min으로 유의성 있게 감소하는 것으로 나타났다(Fig. 4).
Fig. 3. Effect of Patriniae Radix on ․O
2-Generation in LLC-PK
1Cells.
LLC-PK1cells were exposed to H2O2(100 mM) in the presence and absence Patriniae Radix (25, 50, 100, 200 ㎍/㎖). The generation of ․O2-
was measured by DCFH-DA fluorescence probe.
Results are mean±S.D. a) Significantly different from the normal. b) Significantly different from the control (** : p < 0.01, * : p < 0.05). : normal, : control, : experimental
Fig. 4. Effect of Patriniae Radix on NO Generation in LLC-PK
1Cells.
LLC-PK1cells were exposed to H2O2(100 mM) in the presence and absence Patriniae Radix (25, 50, 100, 200 ㎍/㎖). The generation of NO was measured by DAF-2 fluorescence probe.
Results are mean±S.D. a) Significantly different from the normal. b) Significantly different from the control (** : p < 0.01). : normal, : control, : experimental
ONOO
-는 정상 세포에서 142.27±4.73 fluorescence
/min인데 비하여 H
2O
2처리에 의해 311.43±10.01 fluorescence/min으로 생성이 증가하였으나, 敗醬 추출물을 전처리한 경우에 농도 의존적으로 감소 되어 50 ㎍/㎖ 농도에서 251.41±12.55 fluorescence /min, 100 ㎍/㎖ 농도에서 234.64±12.04 fluorescence /min, 200 ㎍/㎖ 농도에서 217.57±11.09 fluorescence /min으로 유의성 있게 감소하는 것으로 나타났다 (Fig. 5).
Fig. 5. Effect of Patriniae Radix on ONOO
-Generation in LLC-PK
1Cells.
LLC-PK1cells were exposed to H2O2(100 mM) in the presence and absence Patriniae Radix (25, 50, 100, 200 ㎍/㎖). The generation of ONOO- was measured by DHR 123 fluorescence probe.
Results are mean±S.D. a) Significantly different from the normal. b) Significantly different from the control (** : p < 0.01). : normal, : control, : experimental
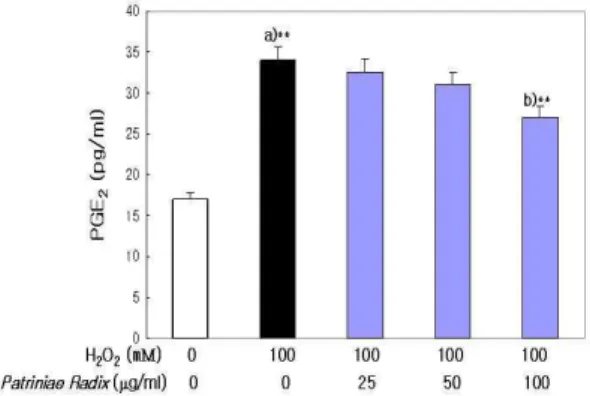
4. H
2O
2처리에 의한 PGE
2생성에 미치는 영향 PGE
2는 정상 세포에서 17.23±0.88 pg/㎖인데 비 하여 H
2O
2처리에 의해 34.92±1.82 pg/㎖로 유의성 있게 증가하였으나, 敗醬 추출물을 전처리한 실험 군에서는 100 ㎍/㎖ 농도에서 27.08±1.33 pg/㎖로 유의성 있게 감소하였다(Fig. 6).
Fig. 6. Effect of Patriniae Radix on PGE
2Level in LLC-PK
1Cells.
LLC-PK1cells were exposed to H2O2(100 mM) in the presence and absence Patriniae Radix (25, 50, 100 ㎍/㎖). Results are mean±S.D. a) Significantly different from the normal. b) Significantly different from the control (** : p
< 0.01). : normal, : control, : experimental
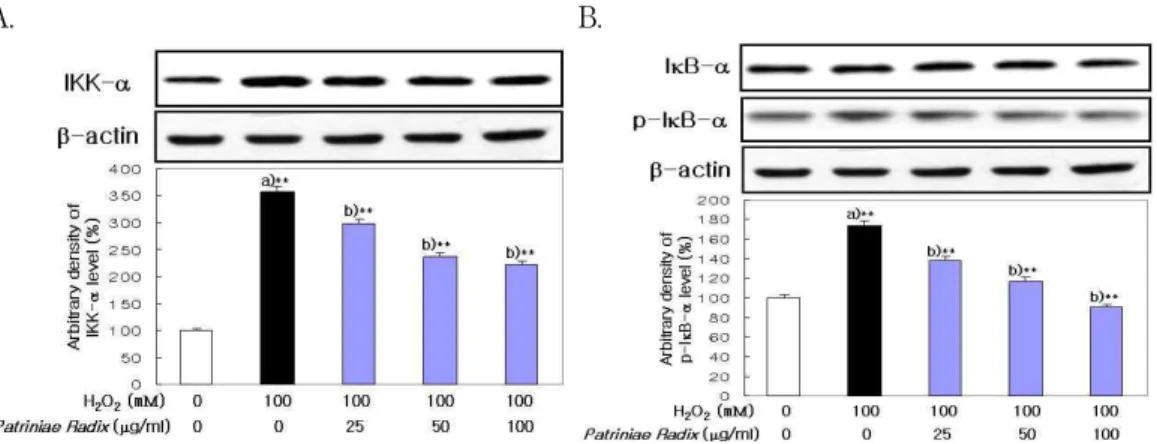
5. H
2O
2처리에 의한 IKK-α 및 phospho-IκB-α에 미치는 영향
IKK-α는 정상 세포에 비하여 H
2O
2처리에 의 해 증가하였으나, 敗醬 추출물을 전처리한 실험군 에서는 감소하는 경향을 보였다(Fig. 7A).
Phospho-IκB-α는 정상 세포에 비하여 H
2O
2처 리에 의해 증가하여 인산화가 진행되었으나, 敗醬 추출물을 전처리한 실험군에서는 감소하는 경향을 보였다(Fig. 7B).
6. H
2O
2처리에 의한 NF-κB 단백질 양에 미치는 영향
敗醬 추출물이 ROS 감수성 전사인자인 NF-κB
의 활성에 미치는 영향을 검토하기 위하여 핵 내
의 p50, p65 양을 조사하였다. NF-κB의 subunit인
p50, p65 핵내 양을 관찰한 결과 정상 세포에 비하
여 H
2O
2처리에 의해 증가하였으나, 敗醬 추출물을
전처리한 실험군에서는 농도 의존적으로 감소하는
경향을 보였다(Fig. 8).
A. B.
Fig. 7. Effect of Patriniae Radix on IKK-α and phospho-IκB-α Expression in LLC-PK
1Cells.
Western blot was performed to detect IKK-α and phospho-IκB-α protein level in cytosol fractions from LLC-PK1. Cells were exposed to H2O2(100 mM) in the presence and absence Patriniae Radix (25, 50, 100 ㎍/
㎖). Results are mean±S.D. a) Significantly different from the normal. b) Significantly different from the control (** : p < 0.01). A : IKK-α protein, B : phospho-IκB-α protein. : normal, : control, : experimental
A. B.
Fig. 8. Effect of Patriniae Radix on NF-κB Expression in LLC-PK
1Cells.
Western blot was performed to detect NF-κB protein level in nuclear fractions from LLC-PK1. Cells were exposed to H2O2 (100 mM) in the presence and absence Patriniae Radix (25, 50, 100 ㎍/㎖). Results are mean±S.D. a) Significantly different from the normal. b) Significantly different from the control (** : p < 0.01).
A : p50 protein, B : p65 protein. : normal, : control, : experimental
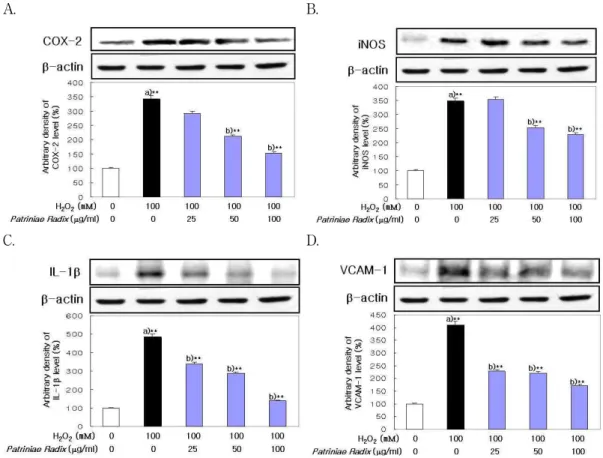
7. H
2O
2처리에 의한 염증 촉진 인자의 발현에 미 치는 영향
敗醬 추출물이 염증 촉진 인자인 COX-2, iNOS, IL-1β 및 VCAM-1의 단백질 발현양에 미치는 영
향을 관찰한 결과 정상 세포에 비하여 H
2O
2처리
에 의해 증가하였으나, 敗醬 추출물을 전처리한 실
험군에서는 농도 의존적으로 감소하는 경향을 보
였다(Fig. 9).
A. B.
C. D.
Fig. 9. Effect of Patriniae Radix on COX-2, iNOS, IL-1β and VCAM-1 Expression in LLC-PK
1Cells.
Western blot was performed to detect COX-2, iNOS, IL-1β and VCAM-1 protein level in cytosol fractions from LLC-PK1. Cells were exposed to H2O2(100 mM) in the presence and absence Patriniae Radix (25, 50, 100 ㎍/
㎖). Results are mean±S.D. a) Significantly different from the normal. b) Significantly different from the control (** : p < 0.01). A : COX-2 protein, B : iNOS protein, C : IL-1β protein, D : VCAM-1 protein. : normal,
: control, : experimental
Ⅳ. 고 찰
敗醬은 마타리과 (敗醬科 : Valerianaceae)에 속 한 多年生 草本인 뚝갈 (白花敗醬) Patrinia villosa (Thunb.) Juss와 마타리 (黃花敗醬) P. scabiosaefolia Fisch. 또는 그 근연식물의 뿌리 또는 帶根全草를 일반적으로 일컫는다
6,7. <神農本草經>
8에 “敗醬味苦 性平 主暴熱火瘡 赤氣 疥瘙疽痔 馬鞍熱氣”라고 처 음으로 언급하였으며, <本草綱目>
19에서는 “敗醬味
微苦甘性平 入手足陽明厥陰經 善排膿破血 故仲景 治癰 及古方婦人科皆用之 乃易得之物而後人不知用 蓋未遇識者耳”라고 하여 解熱, 消腫排膿, 祛瘀止痛 등의 효능으로 腸癰, 産後惡露不止, 産後腰痛, 赤 眼, 瞕痛, 努肉攀睛, 癰疽腫毒, 赤白痢 등의 치료에 쓰이고 있다
6,20.
敗醬의 약리성분으로 白花敗醬의 全草에는 정유
성분, 건조 果枝에는 sinigrin, 根과 根莖에는
morroniside, lognin, villoside 등이 있으며, 黃花敗
醬의 根과 根莖에는 oleanolic acid, hederagenin (mukurosigenin), β-sitosterol, β-D-gluciside와 patrinoside C․D․C1․D1, scabioside A․B․C․D․E․F․
G 등의 다양한 saponin 및 정유 8%, alkaloid, tannin, 전분 등을 함유하고 있다
20. 그 밖에 敗醬의 약리 성분에 대한 연구로 우
21는 triterpenoid와 coumarin 류의 성분을 분리 정제하였으며, 김
22은 마타리에서 Rutin, α-hederin, kalopanax saponin B, hederagenin, 23-hydroxyursolic acid 성분이 분 리되었다고 보고한 바 있다. 한편 敗醬과 관련한 실험연구로는 임
11이 proteinase 활성수용체-2 유발 rats 발바닥 부종에 敗醬根 물추출물이 항염증 효 과가 있다고 하였으며, Wei
12는 敗醬이 adjuvant로 유발된 감염 및 통각과민 rats에 대해 항암작용이 있다고 하였고, 박
13은 肝兪穴 敗醬약침의 처리가 흰쥐의 간기능 회복에 효과가 있음을 시사한 바 있다.
본 실험에서는 우선 敗醬 methanol 추출물의 세 포 독성을 측정해 본 결과 200 ㎍/㎖까지는 세포 독성이 거의 없었으나, 400 ㎍/㎖ 이상에서는 농도 의존적으로 비교적 강한 세포 독성을 나타내었다.
敗醬의 세포 보호 효과를 측정해본 바에서도 200
㎍/㎖까지는 농도 의존적으로 H
2O
2에 의한 세포사 를 저해하는 것으로 나타났다. 이런 敗醬의 특성으 로 안
23은 敗醬 증류수 추출물의 10%까지는 자궁 경부암 세포인 HeLa cell의 증식 억제에 유효하다 고 하였고, 김
24은 증류수 敗醬 추출물의 처리로 남 성호르몬 의존형 전립선암 세포인 LNCaP cell의 생존율 감소가 敗醬 자체의 세포사 효과임을 보고 한 바 있다. 이로써 敗醬은 과도한 농도에서는 자 체에 세포 독성이 있을 수 있으며, 한편으로는 H
2O
2에 의한 세포사에 대해 세포 보호 효과가 있 음을 시사하였다. 따라서 본 연구에서는 敗醬 자체 의 세포 독성을 고려하여 ․O
2-, NO, ONOO
-생성 에 관한 실험에서는 敗醬의 농도를 각각 25, 50, 100, 200 ㎍/㎖로, PGE
2, COX-2, iNOS, IL-1β, VCAM-1, NF-κB 등에 미치는 영향에 관한 실험
에서는 25, 50, 100 ㎍/㎖로 설정하였다.
ROS는 산소기 (oxygen radical) 뿐만 아니라 산 소의 비자유기 유도체 (nor-radical derivatives of O
2)를 총칭하며 정상 산소성 개체 (aerobic organism) 는 ROS의 발생과 항산화 방어계가 서로 균형을 이루고 있다. Superoxide anion radical (․O
2-), hydrogen peroxide (H
2O
2) 및 lipid peroxyl radical (․LOO) 등을 포함한 ROS는 염증반응
25, 고혈압
26및 고지 질 혈증
27에서 세포 신호 전달과정을 매개하고 직 접적으로 조직 손상을 유발하며 혈관 기능을 저하 시킨다. 또한 다양한 염증성 매개물들은 산화제 생 성을 활성화하여 조직 항산화능력을 감소시키므로 조직의 기본적인 대사 구조적 인자에 손상을 주어 독성을 유발한다. 이러한 다양한 세포와 조직에서 의 산화 작용은 독특한 산화 물질을 생성하게 되며 복잡한 ROS의 이차적인 조직 반응이 시작된다
1.
NO는 다양한 세포에서 발생되어 정상 세포 기 능에 중요한 역할을 하고 있다. 염증성 매개 물질 에 노출된 세포에 의해서 생성된 NO의 증가는 NO가 생체 방어와 free radical에 의해 유발된 조 직 손상에 기여한다는 것을 나타낸다
1. 하지만 다 양한 급성 또는 만성 염증반응시 NO의 생성이 증 가하고 조직 손상에 기여하며 조직의 NOS 발현과 NO
2-/NO
3-가 다양한 염증성 질환시 증가되어 있다
28,29. ONOO
-는 강력한 생물학적 산화제이고 hydroxyl radical 같은 반응성, thiol과 lipid의 산화, 그리고 지질과 단백질의 nitrosation/nitration을 포함하는 신호 전달물질이다
30,31. ․O
2-와 NO의 반응이 SOD 가 ․O
2-를 제거하는 것보다 훨씬 빠른 반응이기 때문에 조직의 NO 농도가 SOD의 농도에 접근하 면 언제라도 ONOO
-가 형성될 것이다. 게다가 ONOO
-는 이차적인 nitrated species를 발생하므로 염증 신호 전달물질로 작용한다
1.
본 연구에서 이런 ․O
2-, NO 및 ONOO
-의 염증
신호 전달물질로서의 특성을 참고하여 敗醬 추출
물을 전처리한 LLC-PK
1세포에 H
2O
2처리 후 활
성산소 및 활성질소 생성계를 유도한 결과 ․O
2-,
NO, ONOO
-의 생성을 50, 100, 200 ㎍/㎖에서 농도 의존적으로 유의성 있게 억제하였다.
NF-κB는 쥐의 B 림프구에서 kappa light-chain 발현을 조절하는 전사인자로 발견되었으며
32, 이후 다양한 세포들에서 면역 및 염증반응에서 중요한 역할을 하는 범발적인 전사인자임이 확인되었다
33-35