Analysis of the Anaerobic Bacterial Community in the Earthworm (Eisenia fetida) Intestine
Kwang-Hee Shin, H. Yi1, Jongsik. Chun2, Chang-Jun Cha2, In Seon Kim and Hor-Gil Hur*
Department of Environmental Science and Engineering
1Water Reuse Technology Center, Kwangju Institute of Science and Technology, Gwangju 500-712, Korea
2School of Biological Science, Seoul National University, Seoul 151-742, Korea Received March 16, 2004; Accepted August 5, 2004
Intestinal microbial community structure of earthworm Eisenia fetida was investigated based on 16S rDNA analysis. One hundred different colonies grown on Brain Heart Infusion medium were randomly isolated. Through partial sequence analysis of PCR-amplified 16S rDNA, earthworm intestinal bacteria (EIB) were divided into eight groups, which were further divided into subgroups. Groups EIB 2, EIB 3, EIB 4, EIB 5, EIB 6, EIB 7-1, and EIB 8 showed over 97% similarities to Clostridium bifermentans, C.
butyricum, C. glycolicum, C. celerecrescens, C. lituseburense, Staphylococcus epidermidis, and Propionibacterium acnes, respectively. Group EIB 1 consisting of six subgroups, showed unique pyretic line, found to be most closely related to C. subterminale with 90-95% similarity. Subgroup EIB 7-2 showed 93% similarity to S. epidermidis. Among 100 strains, intestinal microbial community consisted of 49, 13, 13, 5, 4, 2, 11, and 3% EIB 1, EIB 2, EIB 3, EIB 4, EIB 5, EIB 6, EIB 7 and EIB 8, respectively, indications that group EIB 1 was dominant bacterial group in earthworm intestinal bacterial community. Considering earthworm plays key role in improving physical and chemical properties of soil, this study provides valuable information on bacterial community structure of intestine of these ecologically important organisms.
Key words: Earthworm, Eisenia fetida, Microbial community, Intestine, Anaerobic, Clostridium
Earthworms exist in many humid habitats and are considered to be one of the most important organisms among soil invertebrates. They have beneficial effects on soil productivity by improving the physical and chemical properties of soil. Earthworm burrowing creates soil pore spaces, thus increasing water conductivity and air transport.1) They promote soil aggregation by depositing casts rich in glycoproteins, polysaccharides, bacteria, and clay,2) and also affect soil profile formation by mixing litter layers with soil surface layers, which act to deepen humus layers with soil depth.2) Part of the beneficial effects of earthworms on soil structure and development is attributed to their feeding activities. Their consumption pattern involves the breakdown and incorporation of large amounts of mineral soil and organic matter,3) which contain a variety of microorganisms that contribute to the degradation of organic materials. Due to these characteristics, earthworms have been known to be vectors for the dispersal of soil microorganisms4) through lateral and vertical zones, and bioreactors for certain kinds of microorganisms.5) Several studies have shown that earthworms can mediate bacterial transport into different environments.6-8)
Stephens et al. reported that earthworms are important in the transport of beneficial bacteria for use in biological control.
More recently, some bacterial species were shown to survive in egg cocoons of the earthworm species Eisenia fetida9) and were implicated as vectors and contributors for bacterial gene transfer. Recombinant bacterial strains associated with earthworm cocoons were shown to be released into the soil upon hatching and to initiate the biodegradation of the herbicide 2,4-dichlorophenol.10)
In particular, the earthworm Eisenia fetida significantly hastens decomposition of organic matter and rapidly increases its weight on diets high in cellulose, manure, and microorganisms,11) and is a common earthworm together with E. andrei in agricultural ecosystems of Korea12). In addition, E. fetida has been employed as a biological indicator of organic matter stabilization,13) and attention has been given to using the organism for the treatment of biosolid wastes.14) To better understand the role of earthworms in nature and their potential usage as organic waste stabilizers and indicators of soil health, the role of earthworm intestinal microorganisms must be rigorously defined. However, little has been studied on the role of earthworm intestinal microorganisms in terms of the biodegradation of environmental pollutants. In this study, bacteria were isolated from the earthworm intestine under anaerobic conditions, and the indigenous microbial community was characterized. 16S rDNA analysis was used to determine the bacterial community structure of the
*Corresponding author
Phone: +82-62-970-2437; Fax: +82-62-970-2434 E-mail: [email protected]
Abbreviations: EIB, earthworm intestinal bacteria; BHI, Brain Heart Infusion
appropriate inoculants aimed at improving earthworms ability in biodegradation of environmental pollutants.
Materials and Methods
Isolation of bacteria from earthworm intestine.
Earthworms (Eisenia fetida) were collected from soil located at an industrial site, Uiryung, Korea, where organic solvent wastes have been discharged for over 30 years. The surface of each earthworm was washed with 70% ethanol three times and passed through an alcohol lamp three times. The intestinal part of the earthworm was sampled under aseptic conditions and dispersed in a 10-ml dilution blank containing 0.01%
Tween 80 and 0.85% NaCl in an anaerobic glove box (85%
N2: 10% H2: 5% CO2,Coy LAB, Grass Lake, Michigan, USA). After serial dilution, Brain Heart Infusion (BHI) medium was used to grow the intestinal anaerobic bacteria under anaerobic conditions. After a 7-day incubation period, 100 colonies were randomly isolated.
DNA extraction and PCR amplification of 16S rDNA.
Chromosomal DNA was extracted from the 100 bacterial isolates using the procedure of Pitcher15) as modified by Chun and Goodfellow.16) The 16S rDNA was amplified by employing two universal primers, p27F (5'-AGA GTT TGA TCM TGG CTC AG-3') and p1525R (5'-GYT ACC TTG TTA CGA CTT-3').17) The PCR program used for amplification was as follows: 95oC for 15 min, followed by 30 cycles consisting of 94oC for 1 min, 55oC for 1 min, and 72oC for 1 min, and a single final extension step consisting of 72oC for 7 min.
16S rDNA Sequencing. Amplified 16S rDNA fragments were purified using a PCR fragment purification kit (Qiagen, Valencia, CA, USA). Nucleotide sequences were obtained using an ABI 377 XL upgrade DNA Sequencer (Perkin Elmer, Boston, MA, USA) and software provided by the manufacturer. The oligonucleotide primers used in the sequencing reactions have been described previously.17)
Phylogenetic analysis of the microbial community of earthworm intestine. Sequence similarity searches were performed using the BLAST program. The sequence information was then imported into the PHYDIT program18) for assembly and alignment. The 16S rDNA sequences (ca.
500 bp) of each isolate were compared to those of type strains held in the GenBank database. Nucleotide substitution rates were calculated, and phylogenetic trees were constructed by the neighbor-joining method.19) Escherichia coli ATCC 11775T was used as the outgroup.
Physiological and biochemical characterization. The commercial identification kit (API 20A, BioMerieux®, Marcy- lEtoile, France) was used for physiological and biochemical characterizations of the bacterial strains. Experiments were performed as described in the manufacturers manual.
Earthworms are known to play important roles in the structure and chemical properties of soil. Physical movement of the earthworms through soil enables them to mix and granulate the diverse organic material of soil in their gut intestine. Once consumed, organic materials may be subjected to digestive enzymes produced by the earthworms or organisms living in their intestine. This understanding led us to assume that the gut intestine of earthworms may contain a diverse community of bacteria that are most likely living in symbiotic- and/or associative-type relationships. If we better understand the bacterial community structure of the earthworm intestine, we may use the combined biological systems between earthworm and intestinal bacterial structure to effectively manage and improve soil properties. In this study, we were interested in analyzing the culturable bacterial community structure of the earthworm gut intestine.
One hundred earthworm intestinal bacterial (EIB) colonies growing on the BHI medium were randomly isolated. To identify EIB isolates at the species level, molecular phylogenetic trees were constructed based on the 16S rDNA sequences from evolutionary distances by the neighbor- joining method.19) The EIB isolates were divided into eight different groups based upon 16S rDNA sequence analysis, and each group was further divided into subgroups. The subgroups of the eight different groups are located in close proximity to each other on the phylogenetic tree (Figs. 1- 3).
The groups EIB 2, EIB 3, EIB 4, EIB 5, EIB 6, EIB 7-1, and EIB 8 showed more than 97% similarity to Clostridium bifermentans NCIMB10716, C. butyricum ATCC19398, C.
glycolicum DSM1288, C. celerecrescens DSM5628, C.
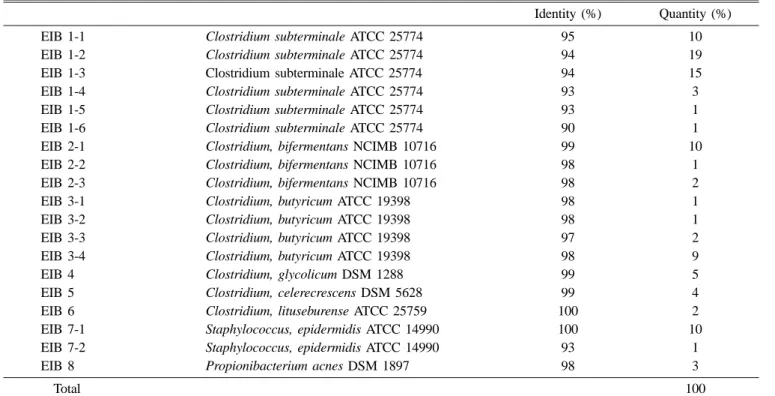
lituseburense ATCC25759, Staphylococcus epidermidis ATCC14990, and Propionibacterium acnes DSM1879, respectively. Ninety-seven percent similarity is considered to be the cutoff value indicating species identity when compared to reference strains.20) The group EIB 1, on the other hand, was divided into six subgroups, which were closest in similarity to C. subterminale ATCC25774 with 90-95% similarity.
Subgroup EIB 7-2 showed 93% similarity to Staphylococcus epidermidis ATCC14990. 16S rDNA sequence similarities of EIB 1-1, EIB 1-2, EIB 1-3, EIB 1-4, EIB 1-5, and EIB 1-6 to their closest reference strains were 95, 94, 94, 93, 93, and 90%, respectively (Table 1). It is clear from phylogenetic analysis that group EIB 1 is a new species of the Clostridium subphylum, which is the nearest neighbor to C. subterminale ATCC 25774. Additionally. The subgroup EIB 7-2 was a member of the Staphylococcus subphylum and was the nearest neighbor to S. epidermidis ATCC14990 (Fig. 3), showing 93% 16S rDNA sequence similarity (Table1).
Although the 16S rDNA sequence analysis is effective for the identification of organisms on the levels of genus and species, this method cannot differentiate strains at the
subspecies level and is, therefore, not an appropriate method to measure intraspecies relationships.20) Biochemical identification of EIB was carried out using a commercial identification kit to overcome the limitation of 16S rDNA-based analysis.
Although each EIB group showed a high similarity of 16S rDNA and were located in close proximity in the phylogenetic tree, all EIB groups showed different substrate utilization patterns (Table 2). Six subgroups of the EIB 1 group were classified into five types according to their biochemical properties. The EIB 1-1 subgroup showed the same results
with the EIB 1-5 subgroup, except that glycerol acidification was positive for the EIB 1-5 subgroup. EIB 1-2 and EIB 1-4 subgroups only showed a positive result for the hydrolysis reaction of gelatine from among the biochemical tests employed. The EIB 1-3 subgroup was shown to utilize L- tryptophan that is known to be related to indole formation.
The EIB 1-6 subgroup showed a positive reaction for the acidification of D-maltose and D-xylose, and the hydrolysis of gelatine. The EIB 2 group was also classified into three subgroups according to their biochemical reactions. The EIB Fig. 1. Phylogenetic position of the isolate Clostridium sp. and related taxa. Similarity and distance matrices were constructed based on available 16S rDNA sequences. The scale bar represents the expected number of changes per sequence position.
2-1 subgroup had the same biochemical properties as the EIB 2-3 subgroup, except that the EIB 2-3 subgroup showed acidification of D-cellobiose. The EIB 2-2 subgroup underwent H2O2 catalase activity with utilization of L- tryptophan, urea, D-glucose, and D-saccharose. The EIB 3 group was also divided into two subgroups based on H2O2
catalase activity. The EIB 3-1 and EIB 3-2 subgroups showed positive H2O2 catalase activity, whereas EIB 3-3 and EIB 3-4 did not. The EIB 4, 5, 6, and 8 groups, which were not divided into subgroups, showed unique substrate utilization patterns, differentiating them from the others. The EIB 7-1 and EIB 7-2 subgroups showed the same biochemical reaction patterns except that acidification of D-cellobiose and L-rhamnose
occurred only in EIB 7-1 (Table 2). Following phylogenetic and biochemical analyses using the BHI medium under anaerobic conditions, the spectrum of the microbial community structure in the earthworm intestine obtained was unexpectedly narrow. Most of the EIB groups belonged to Clostridium that has been also known to be the predominant species in human beings in addition to Bacteroides, Bifidobacterium, Eubacterium, Peptococcus, Peptosteptococcus, and Ruminococcus, which outnumber aerobic bacteria (facultative anaerobes) by a factor of 100-1,000.21,22) Based on the closest relative evaluated by 16S rDNA analysis, the anaerobic microbial community in the earthworm E. fetida was composed of C. subterminale (49%), C. bifermentans Fig. 2. Phylogenetic position of the isolate Staphylococcus sp. and related taxa. Similarity and distance matrices were constructed based on available 16S rDNA sequences. The scale bar represents the expected number of changes per sequence position.
Fig. 3. Phylogenetic position of the isolate Propionibacterium sp. and related taxa. Similarity and distance matrices were con- structed based on available 16S rDNA sequences. The scale bar represents the expected number of changes per sequence position.
(13%), C. butyricum (13%), C. glycolicum (5%), C.
celerecrescens (4%), C. lituseburense (2%), S. epidermidis (11%), and Propionibacterium acnes (3%) (Table 2). Among the bacterial strains, the Clostridium strain, which is Gram- positive, endospore-forming, and a strict anaerobic rod, was shown to be the dominant anaerobic species in the earthworm
intestinal bacterial community. This is the first report on the community structure of culturable anaerobic bacteria isolated from the earthworm intestine. More work, therefore, is needed to compare the microbial community structure in the surrounding soil environment of earthworms with that in the earthworm intestine.
Table 1. Sequence similarities of 100 isolates to closest relatives and portion of each group
Identity (%) Quantity (%)
EIB 1-1 Clostridium subterminale ATCC 25774 95 10
EIB 1-2 Clostridium subterminale ATCC 25774 94 19
EIB 1-3 Clostridium subterminale ATCC 25774 94 15
EIB 1-4 Clostridium subterminale ATCC 25774 93 3
EIB 1-5 Clostridium subterminale ATCC 25774 93 1
EIB 1-6 Clostridium subterminale ATCC 25774 90 1
EIB 2-1 Clostridium, bifermentans NCIMB 10716 99 10
EIB 2-2 Clostridium, bifermentans NCIMB 10716 98 1
EIB 2-3 Clostridium, bifermentans NCIMB 10716 98 2
EIB 3-1 Clostridium, butyricum ATCC 19398 98 1
EIB 3-2 Clostridium, butyricum ATCC 19398 98 1
EIB 3-3 Clostridium, butyricum ATCC 19398 97 2
EIB 3-4 Clostridium, butyricum ATCC 19398 98 9
EIB 4 Clostridium, glycolicum DSM 1288 99 5
EIB 5 Clostridium, celerecrescens DSM 5628 99 4
EIB 6 Clostridium, lituseburense ATCC 25759 100 2
EIB 7-1 Staphylococcus, epidermidis ATCC 14990 100 10
EIB 7-2 Staphylococcus, epidermidis ATCC 14990 93 1
EIB 8 Propionibacterium acnes DSM 1897 98 3
Total 100
Table 2. Phenotypic characteristics of EIB isolated from the earthworm intestine
Substrate EIB group based on 16S rDNA analysis
1-1 1-2 1-3 1-4 1-5 1-6 2-1 2-2 2-3 3-1 3-2 3-3 3-4 4 5 6 7-1 7-2 8
Tryptophane - - + - - - + - + - - - - - + - - - -
Urea + - - - + - - + - - - - - - - - - - -
Glucose + - - - + - + - + + + + + + + - + + +
Mannitol - - - - - - - - - + + + + - - - - - -
Lactose - - - - - - - - - + + + + - + - - - -
Saccharose + - - - + - - + - + + + + - + - + + -
Maltose + - - - + + + + + + + + + + + + + + +
Salicin - - - - - - - - - + + + + - + - - - -
Xylose - - - - - + - - - + + + + + + - - - -
Arabinose - - - - - - - - - + + + + + + - - - -
Gelatin + + - + + + + + + - - - - - + + + + +
Ferric citrate - - - - - - + + + + + + + - + - - - -
Glycerol - - - - + - - - - + + + + + + - - - -
Cellobiose - - - - - - - - + + + + + - + - + - -
Mannose + - - - + - + + + + + + + + + - + + +
Melezitose - - - - - - - - - + + + + + - - - - -
Raffinose - - - - - - - - - + + + + + + - - - -
Sorbitol - - - - - - - - - + + + + + - - - - -
Rhamnose - - - - - - - - - + + + + + + - + - -
Trehalose - - - - - - - - - + + + + + + - - - -
H2O2 calalase + - - - + - - + - + + - - - + - + + -
his criticism and edition on this manuscript. This work has been supported by BioGreen 21 Project of Rural Development Administration, Korea.
References
1. Edwards, C. A. and Lofty, J. R. (1972) Biology of earth- worms. Chapman Hall, London.
2. Furlong, M. A., Sigleton, D. R., Coleman, D. C. and Whit- man, W. B. (2002) Molecular and culture-based analyses of prokaryotic communities from an agricultural soil and the burrows and casts of the earthworm Lumbricus rubellus.
Appl. Environ. Microbiol. 68, 1265-1279.
3. Piearce, T. G. (1978) Gut contents of some lumbricid earth- worms. Pedobiologia 18, 153-157.
4. Madsen, E. L. and Alexander, M. (1982) Transport of Rhizobium and Psedomonas through soil. Soil Sci. Soc. Am.
J. 46, 557-560.
5. Parle, J. N. (1963) Micro-organisms in the intestines of earthworms. J. Gen. Microbiol. 31, 1-11.
6. Daane, L. L., Molina, J. A. E., Berry, E. C. and Sadowsky, M. J. (1996) Influence of earthworm activity on gene trans- fer from Psedomonas fluorescens to indigenous soil bacte- ria. Appl. Environ. Microbiol. 62, 515-521.
7. Daane, L. L., Molina, J. A. E. and Sadowsky, M. J. (1997) Plasmid transfer between spatially separated donor and recipient bacteria in earthworm containning soil micro- cosms. Appl. Environ. Microbiol. 63, 679-686.
8. Stephens, P. M., Davoren, C. W., Ryder, M. H. and Doube, B. M. (1993) Influence of the lumbricid earthworm Apor- rectodea trapezoides on the colonization of wheat roots by Pseudomonas corrugata strain 2140R in soil. Soil Biol. Bio- chem. 25, 1719-1724.
9. Fischer, O. A., Matlova, L., Bartl, J., Dvorska, L., Svas- tova, P., Maine, R. D., Melicharek, I., Bartos, M. and Pav- lik, I. (2003) Earthworms (Oligochaeta, Lumbricidae) and mycobacteria. Vet. Microbiol. 91, 325-338.
10. Daane, L. L. and Haggblom, M. M. (1999) Earthworm egg capsules as vectors for the environmental introduction of biodegradative bacteria. Appl. Environ. Microbiol. 65, 2376-
earthworm Eisenia foetida on microorganisms and cellu- lose. Soil Biol. Biochem. 16, 491-495.
12. Hong, Yong, Kim T. H. and Na Y. E. (2001) Identity of two earthworms used in vermiculture and vermicomposting in Korea: Eisenia andrei and Perionyx excavatus. Korean Journal of Soil Zoology. 17, 185-190.
13. Hartenstein, R. (1981) Sludge decomposition and stabiliza- tion. Science 212, 743-749.
14. Priya, K. and Garg, V. K. (2003) Vermicomposting of mixed solid textile mill sludge and cow dung with the epigeic earthworm Eisenia foetida. Bioresource Technology 90, 311-316.
15. Pitcher, D. G., Saunders, N. A., and Owen, R. J. (1989) Rapid extraction of bacterial genomic DNA with guanid- ium thiocyanate. Lett. Appl. Microbiol. 8, 151-156.
16. Chun, J. and Goodfellow, M. (1995) A phylogenetic analy- sis of the genus Nocardia with 16S rRNA gene sequences.
Int. J. Syst. Bacteriol. 45, 240-245.
17. Lane, D. J. (1991) 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics, Stackebrandt, E.
and Goodfellow, M. (ed.) pp. 115-175, Wiley, Chichester 18. Chun, J. (1995) Computer-assisted classification and identi-
fication of actinomycetes. Ph.D. Thesis, University of New- castle, Newcastle upon Tyne, UK.
19. Saitou, N. and Nei, M. (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406-425.
20. Stackebrandt, E. and Goebel, B. M. (1994) Taxonomic note:
a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacte- riology. Int. J. Syst. Bacteriol. 44, 846-849.
21. Simon, G. L. and Gorbach, S. L. (1984) Intestinal flora in health and disease. Gastroenterology 86, 174-193.
22. Salminen, S., Bouley, C., Boutron-Ruault, M. C., Cum- mings, J. H., Franck, A., Gibson, G. R., Isolauri, E., Moreau, M. C., Roberfroid, M. and Rowland, I. (1998) Functional food science and gastrointestinal physiology and function. Br. J. Nutr. 80, S147-171.