285
<원례보저
>
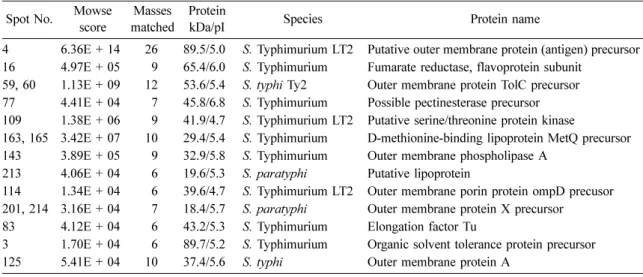
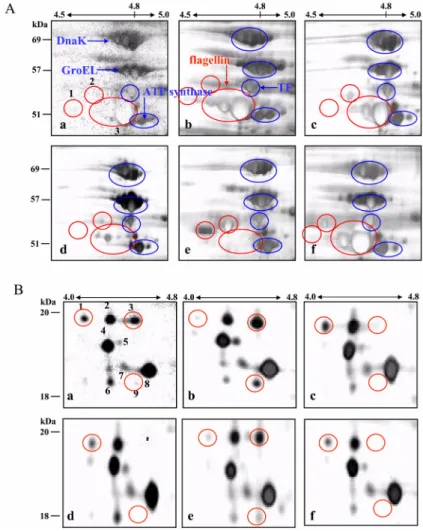
Proteome analysis between diverse phenotypes of Salmonella enterica subspecies enterica serovar Typhimurium ( S. Typhimurium)
Gee-Wook Shin
1, In-Seok Cha
2, Woo-Won Lee
3, Seong-Won Nho
2, Seong-Bin Park
2, Ho-Bin Jang
2, Yong-Hwan Kim
2, Tae-Sung Jung
2,*
1
College of Veterinary Medicine, Chonbuk National University, Jeonju 561-756, Korea
2
College of Veterinary Medicine, Gyeongsang National University, Jinju 660-701, Korea
3