맥아당결합 단백질에 융합된 면역결필 바이러스 인테그라제의 생산 및 분석
김도진 • 오유택 ■신차균*
중앙대학교 산업대학 (Received October 25. 1997)
P r o d u c t i o n a n d C h a r a c t e r i z a t i o n o f H u m a n I m m u n o d e f i c i e n c y V i r u s I n t e g r a s e F u s e d
w i t h a M a l t o s e - B i n d i n g P r o t e i n
D o J i n K im , Y o u - T a k e O h a n d C h a - G y u n S h in * College o f In d u stria l Studies, Chung-Ang University, A n su n g 456-756, Korea
Abstract~Retroviral integrase is required for integration of viral DNA into the host cell chromosome.
Human immunodeficiency virus type-1 integrase was partially purified as a part of a fusion protein linked to a maltose-binding protein and characterized in terms of an endonucleolytic activity. The concentration of the fusion protein purified through an amylose column was about 12 mg/m/, in dicating that the solubility of the fusion protein is highly increased by the presence of a maltose- binding protein, considering that the integrase protein alone is poorly solubilized. The en
donucleolytic activity of the fusion protein was detected at 0.1 to 1.0 mM MiV■수 ion. but not at any concentrations tested of Mg"나 ion.
Keywords □ Fusion protein, integrase. maltose-binding protein, endonucleolytic activity.
인간면역결핍바이러스는인간외T-임파구에 침투 한후원형질에서 바이러스의R N A 유전자를D N A 유
전자로견환하고. D N A 유전자가핵으로둘어가서T-
임파구의유전자를끊고그사이에중합하게된다. 이러
한바이러스D N A 유전자의중합은바•이러스의복제와
생성을위하여필요불가결한과정이며, 이과정을중개
하는것이바이러스외특유외효소인integrase이다.*^
바이러스D N A의중합과정에서integrase는직선형
외바이러스D N A유전자의양쪽3' 말단부분에서각기
두개의nucleotide을제거하고, 두개의nucleotide가 짧아진끝을이미사다리형태로절단해놓은숙주세포의 유전자에 연결시킨다. 연결되지않은 5^ 말단은숙주세 포의 유전자복구효소들에의하여두개의nucleotide
• 본 논문에 관한 문의는 이 저자에게로
( 전화) 0334-70-3067 (팩스) 0334-675-0409
가절단된후숙주세포의유전자에연결됨으로써, 바이
러스D N A t 전자가완전히숙주세포의유전자에중합
하게된다."* 바이러스D N A가숙주세포의유전자에중 합될때 , 바이러스D N A가둘어가는숙주세포D N A내 의부위는벌다른특이성이없다. 그러나, 면역결핍바이 러스의integrase가바이러스D N A 끝외두개핵산을 제거하는활성은면역결핍바이러스외 D N A에서만나 타난다.® 면역결핍 바이러스의 integrase의유전자 는바이러스유전자pc)l의끝부분에 존재하며, 감염된 세포에서 바이러스외 다른 효 소둘과 함께 po-
lyprotein외일부분으로생성된다. 바이러스조립과정
에서바이러스외 단백분해효소에 외하여polyprotein
에서절단되어바이러스입자안에포함된다.®'" 박테러 아에서발현시켜 정제한면역결핍 바이러스의 활성은
바이러스D N A 끝외 염기서열을 닮은이중가닥oli-
gonucleotide의한쪽가닥외5' 말단을방사능으로표
46
융합단백질의 일부로써의 인테그라제 톡성 47
식하고 integrase로처러할때. 두개의핵산이 3' 말단 에서 제거되는 것을 이용한 endonucleolytic clea- vage 활성으로측정한다 ®
면역결핍바이러스의 integrase 단백질은구조및기 능상에서 N-말단영역 , 중앙영역 , C-말단영역으로구분 된다〶9■10) N-말단영역에는 각기 두 개의 histidine과 cysteine의 Zn-finger 부분이존재하나, Zn가단백질 의효소적활성이나구조외변화에별다른영향을주지 않는다. 그러나, 이영역은 integrase의 dimerization 에관 하 고 있으며, 이부분이제거되면, 효소적활성을 상실한다® 중앙영역은효소적활성이직접적으로발현 되는부분으로, 이부분외염기서열은유사한다른바이 러스둘의 integrase에서도 거의 일처되고 있다. C-말단영역은 DNA에결합하는부분으로소수성을지 니고었다. 박테리아에서발현하여생성한 integrase는 용해성이 매우낮으며, 용액중에서불안정하여쉽게침 전으로떨어져서활성을상실하여 연구에많은어려움 을주고있다. 본연구에서는용해성이좋은맥아당결합 단백질에 integrase 롤 붙인 융합단백질로써 in- tegrase를생성하고, 이융합단백질이보이는 integra- se의 endonucleolytic 활성을분석하였다.
실험방법
시약및사용기기- 제한효소 (BamHI, Hindlll), T4 DNA ligase. T4 polynucleotide kinase (T4 PNK), pMalc2, amylose resin등은 New England Biolabs사에서구입하였다. DNA Sequencing kit는 United States Biochemicals사에서구입하고, Isop- ropylthiogalactopyranoside (IPTG), agarose, am- pidllin등은 Sigma사에서 구입하였다. [ /4지-ATP (3,000 Ci/mmol: lCi = 37GBq)는 머국Amershain 사에서구입하였고. Sephadex G25 column은 Phar- macia사에서구입하였다. 사용된기기로는세포의 파 괴를위하여 미국 Misonix사의초옴파파쇄기 (Soni- cator: XL2020) 을사용하였으며, 박테리아의 배양은 한국 비전과학사의 진탕배양기를 이용하였다. Oli- gonucleotide는한국바이오니아에서합성하였고, 사용 하기직전에 RAGE 정제하였다.
발현벡터의체작및밟현- 발현벡터를제작하기위 하여 면역결핍바이러스 integrase유전자를 함유하는 pQEIN을 제한효소 BamHI과 Hindlll로 처리하고
agarose 전기영동한후, 0.9 b의 DNA 절편을 glass milk 를 사용하여 분리하였다. 벡터 pMalc2 를 BamHI과 Hindlll로처리하여 6.7 Kb의 DNA 절편을 분리하고, integrase유전자와섞고 T4 DNA ligase와 함께 16°C에서 4 시간처러하고반응물을박테러아(£.
coli. XLl-blue)에넣어 42°C에서 3 분간처리하여 형 질전환시켰다. Ampicillin 존재하에 성장하는 균주들 외 폴라스머드를 추출하여 제한효소로 처러하여 in- tegrase유전;^]•틀함유하는균주률선정하고, 재조합된 발현벡터에 integrase유전자가 벡터에 in-frame으로 재조함되었음을 DNA 염기서열분석으로 최종 확인하 였다. 융합단백질외 발현은발현벡터를함유하는균주 틀 100 ug/m/ ampicillin을함유하는 LB배지에서 흡 광도가 595 nm에서 0.6될때까지배양한후, 최종농도 가 0,3 mM이되도록 IPTG를넣고 3 시간더배양하여 유도하였다. 단백질의발현유무는배양액 1 m/을원심 분리하여 얻은균주체률전기영동염색액에 넣고가열 한후, SDS-PAGE 전기영동으로확인하였다.
웅함단백질의 정제- Integrase률 함유하는융합단 백질의대량정제를위하여발현벡터룰함유하는균주의
500m/ 배양액을위에서와같은조건에서 발현을유도
하고. 발현이확인된균주의 pellet을 5 ml의완충용액 A (10mM sodium phosphate, [pH 7.2], ImM p- mercaptoethanol (P-MeOH), Im M EDTA, 1M NaCl)에현탁한후, 초옴파파쇄기을이용해서 세포률 파괴하였다. 파괴된세포시료틀 4°C에서 40,000 X g로 20 분간원심분리을하여 4.5 m/ 정도의상등액을얻은 다옴. 같은부피의완층용액 B (10 mM sodium phos
phate, (pH 7.2], 1 mM p-MeOH, 1 mM EDTA)로 섞어 희석하였다. 머러 준버한 0.3m/의 amylose resin을함유하는컬럼에희석액을통과시키고, 5 m/의 완층용액 C (10 mM sodium phosphate, [pH 7.2], 1 mM P-MeOH, 0.5 M NaCl, 1 mM EDTA, 0.25 %
Tween 20)로서 컬럼을층분히 씻어주었다. 150u/의 완층용액 EKlOmM sodium phosphate, (pH 7.2], 1 mM p-MeOH, 0.5 M NaCl, 1 mM EDTA)을 step- wise형태로 10번통과하여 융함단백질을 추출하였다.
수집된 각 분획에 존재하는 단백질의 분포는 SDS-
PAGE 전기영동으로확인하고, 융합단백질이다량으로
있는부분만을따로모아, Biospin30을통과시켜서 완 충용액에존재하는 maltose와고농도외 염룰제거하였 다. 단백질의양을 Bradford assay 방법으로즉정하고,
정제된융합단백질을소분하여 -70T:에보관하였다. 융 합단백질의 integrase활성과의 비교를위하여 사용된 histidine 표식된 integrase는본실험설에서 이미발표 한방범에따라정제하였다.
이 중 가 닥 oligonucleotide 기 질 의 제 작 - 융합단백질 에존재히는 integrase의활성의측정에사용될기질로 써, 방사능으로표식된이중가닥 oligonucleotide 기질 을 다옴과같이 제작하였다. 먼저 면역결핍바이러스의 U5-LTR 끝부분과같은염기서열을갖는 20 mer 크기 의 2 개의 oligonucleotide (K16 [U5-LTR, + strand], 5-TGTGGAAAATCTCTAGCAGT-3^ : K17 [U5- LTR, -strand], 5-ACTGCTAGAGATTTTCCACA- 30를 PAGE 정제하였다. 정제된 K16을방사능으로표 식하기위하여, 15pmol K16, 50uCi (y*-P]-ATP 및 1 unit T4 polynuclotide kinase을 40 u/의반응용액 (70 mM Tris-HCl [pH 7.4], 10 mM MgCU,5 mM dithiothreitol)에서 15 분간 37°C에서 반응시키고, 최 종농도로 10 mM EDTA. 100 mM NaCl이 되도록 EDTA와 NaCl를첨가하고, 85°C에서 15분간가열하였 다. 상보적인염기서열을갖는 K17 (30 pmol)을첨가한 후. 3 분간끓인다옴천천히 식혀서, 상보적인 결합이 일어나게하였다. 방사능으로표식된기질둘과반응하지 않은방사능은 Sephadex G25 컬럼를통과시켜서분리 하였다.
웅 함 단 백 질 의 integrase활 성 의 측 정 - 융합단백질의 integrase외활성은바이러스 LTR끝외염기서열을닮 은 20 bp의이중가닥 oligonucleotide중한쪽가닥의 3' 말단의 2 개의 nucleotide가 제거되는 endonudeo- lytic 활성을측정하여조사하였다. 방사능으로표식된 0.1 pmol 이중가닥 oligonucleotide 기질과 30 pmol 융 합단백질 (MBP-IN)을 10 U;부피의기본 반응완충액 (10 mM Tris-HCl [pH 7.4))에각실험에서표시된성 분을가감하여서 90분간 33/에서반응시켰다. 반응정지 용액 (95% formamide, 20 mM EDTA, 0.05% bro- mophenolblue, 0.05% xylene cyanol FF) 4 u/을첨 가하여반응을정지시켰다. 반응한생성물의확인은반 응물을 20 % 번성 acrylamide gel에서전기영동하고. 젖은 gel을 X-ray 필림에노출하여측정하였다.*®
결 과 및 고 찰
발 현 벡 터 의 제 작 및 밝현 - 면역결핍바이러스의 in-
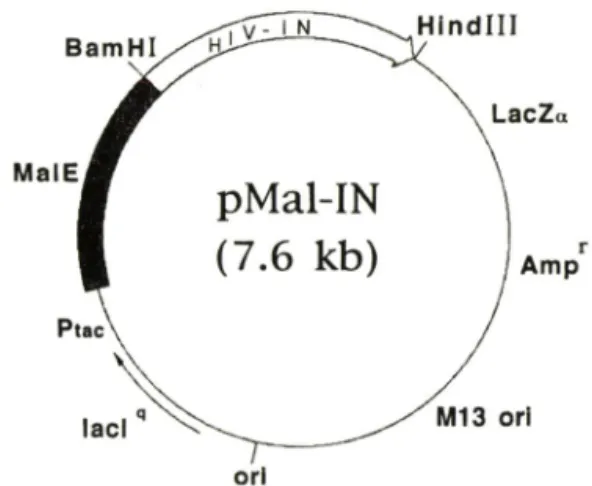
Fig. 1 — Genetic map of a ftision protein expression vec
tor (pMal-IN) coding a maltoseHbinding protein and a HIV-1 integrase.
tegrase 유전자는바이러스 pol 유전자의 3' 말단지역 에존재하며, 바이러스자가복제할때에 GarPol po- lyprotein의 C-말단에 일부분으로써 생산되어 바이러 스조럽과정에서단백분해효소에의하여절단됨으로써 활성을 갖는다.®본연구에서 integrase외 유전자는 이미 본실험실에서 integrase 발현벡터로써 발표한 pQEIN에서얻었다.*^* 맥아당결합단백질에융합된 in- tegrase의 발현벡터는 pQEIN을 BamHI 및 Hin- dlll로처러하여 얻은 0.9Kb의 DNA 절편을 pMalc 2에옮겨서제작하고재조합된발현벡터틀 pMal-IN으 로명명하고, 최중적으로 DNA 염기서열을분석하여
integrase 유전자가맥아당단백질외끝부분에 정확하
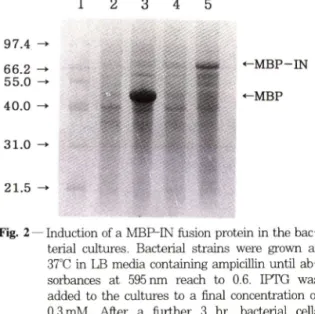
게 연결되었음을 확인하였다 (Fig. 1). 이벡터는 in- tegrase유전자가맥아당결합단백질의유전자 (MalE) 의뒤부분에직접연결되어 있어분자량 74,000Da되 는융합단백질이 IPTG가처리된균주에서생성되게한 다. pMal-IN이융합단백질을잘발현하는가를조사하 기위하여 맥아당결합단백질외 유전자만을함유하는 발현벡터 (pMalc2)와융함단백질의 벡터 (pMal-IN) 외균주들을각기 IPTG를첨가한것과안한것의단백 질생성을비교하였다 (Fig. 2). IPTG를첨가하지않 은 배양에서는 맥아당결합단백질이나, 용합단백질이 거의 생성되지않아단백질의 발현이 lac repressor에 의하여잘억제되고있으나 (lanes 2 and 4), IPTG를 첨가한배앙에서는맥아당결합단백질과융합단백질이
매우높은수준으로생성되고있옴을보여준다 (lanes
3 and 5).
/. Pharm. Soc. Korea
융합단백질의 일부로써의 인테그라제 톡성 49
1 2 3 4 5 9 10 11 12
97.4 66.2 55.0 40.0
31.0
21.5
-MBP-IN
-MBP
Fig. 2 — Induction of a MBP-IN fusion protein in the bac
terial cultures. Bacterial strains were grown at 37°C in LB media containing ampicillin until ab
sorbances at 595 nm reach to 0.6. IPTG was added to the cultures to a final concentration of 0.3 mM. After a further 3 hr. bacterial cells were collected and analyzed. Lane 1, protein molecular size markers in kDa: lanes 2 and 3.
strain containing pMalc2: lanes 4 and 5. strain containing pM aH N: lanes 3 and 5. IPTG added.
용합단백질의정제- 융합단백질의정제는융합단백 질의일부인맥아당결합단백질이 amylose잔기에결합 하는성질을이용하였다.융합단백질의 생성이 확 인된배양외박테리아펠렛을 1 M NaCl를함유하는인 산완충용액에서 초음파파쇄기로파괴하여세포내의단 백질돌이 세포밖으로나오도록하고 원심분리후 상등 액을 amylose resin을통과시켜서결합된단백질들을 맥아당용액으로 추출하였다 (Fig. 3). 융합단백질의 amylose 잔기에대한결합은매우효과적으로, 원심분 리후 상등액에 있던 많은융합단백질돌은 amylose resin을통과한여과액에서는상당부분결합하고, 소량 만이남아었다 (lanes 1 and 2). Amylose resin을중
분히세척후 10mM 맥아당용액으로결합된단백질들
을추출하였을때, 두번째및세번째분획에서가장많 이추출되었다 (lanes 4 and 5). 세번째분획외 경우
섞여었는약 5 % 정도의박테리아의 단백질을포함하
여농도가약 12mg/m;이다. 이러한용해도는기존에 integrase의생산에 이용되는다른방법들에서의 in- tegrase외용해성과 비교하여 약 4 배높온 것이다. Integrase는일반용액에서 용헤성이 매우낮다. 따라 서. 여러번의컬럼을거처는전형적인생화학적인방법 에의한정제는매우어렵다. 이러한이유에서현재많 은연구자들은사용하는방법흔두가지특정이있다. 첫
Fig. 3 — Purification of a MBP-IN fusion protein using an amylose affinity chromatography. A bacterial pel
let from a 500 m/ culture treated with IPTG was resuspended and sonicated. The clear su
pernatant was collected and passed through an amylose column. Lane 1. supernatant: lane 2, flow-through; lane 3. washout: lanes 4-12, elu- ate in 10 mM maltose.
째, histindine 표식을이용한 affinity 정제방범으로, integrase 단백질의 앞 또는뒷부분에 몇 개외 his-
tidine잔기를붙여서니켈이붙은컬럼을이용하여 한
번의 컬럼 정제과정만으로도상당히 순수한단백질을 얻는다. 둘째, 박테리아를분쇄하는용액에 계면 활성제로 CHAPS를첨가하였다. 이계면활성제는 his- tidne표식을 함유하는 integrase의 용해성을 높여준 다. CHAPS 이외의다른많은계면활성제들이 조사되 었으나, 단백질을번성시키지않고 CHAPS보다용해성 을높이는것은보고되지않고있다/ ^ 그러나. 이런방 법도 integrase의용해성을 3 mg/m/ 이상높이기어렵 기때문에단백질구조연구에많은어려움이있다. 또한 integrase의활성억제제의탐색이나단백질외구조연
구를위한대량생산에는 CHAPS가매우고가이기때
문에 histidine표식을이용하는방범의 사용이 제한되 고었어, 새로운방법의 개발이요구되고있다. 따라서, 용해성이좋은맥아당단백질을이용하여 CHAPS를사 용하지않고 integrase를고농도외 융합단백질의 형태 로생산하는것도이러한문제점을극복하는하나의선 텍이될수었다. 본실험에서는융합단백질의효소적활 성을조사하기위하여세번째분획의단백질들을 2 mg/
m/로희석하여융합단백질의 integrase의활성을분석 하였다.
융합단백절의integrase 활성- 에서리트로
TI 0
N
20mer
ISm er
Fig. 5 — Effect of various buffer components on en- donucleolytic activity of a MBP-IN fusion pro
tein. The basic reactions were formulated in the presence of Im M Mn'*■나' as described in Fig. 4. Additional components were added to the reaction as follows: lane 1. oligonucleotide only: lane 2. histidine-tagged IN: lanes 3-13.
MBP-IN: lane 4. Mr/'*' omitted; lanes 5-7. 10, 20 and 50 mM -MeOH ; lanes 8-10, 10. 20 and 50 mM CaCl2- lanes 11-13, 1, 3 and 5 mM EDTA respectively.
/. Pharm. Soc. Korea
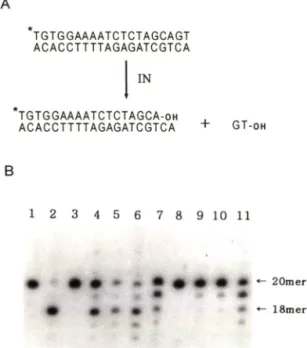
Fig. 4 — Endonucleolytic activity of a MBP-IN fusion protein in the presence of metal ions. (A) Schematic diagram of endonucleolytic reaction.
(B) Endonucleolytic activity of a MBP-IN. Pro
teins (MBP-IN or hisitine-tagged IN) of 30 pmol were incubated at 33"C for 90 min with radiolabelled duplex oligonucleotide of 0.1 pmol in 10 mM TrisHCl (pH 7.4) containing the com
ponents indicated below: lane 1. oli
gonucleotide only: lane 2, histidine-tagged IN with 1 mM Mn 니: lane 3-11. MBP-IN : lanes 4- 7. 0.1. 0.5. 1 and 10mM M n" " : lanes 8-11. 0.1.
0.5. 1 and 10 mM respectively.
바이러스 integrase외활성은바이러스 DNA외염기 서열을닮은이중가닥의 oligonucleotide를이용하여 즉정 할수있다.^ ^' uffro에서 integrase는이중가 닥 oligonucleotide중한쪽가닥의 3' 말단에 있는두 개외잔기를선택적으로쌀라낸다. 본연구에서는쌀라 질가닥의 oligonucleotide외 5' 말단을 방사능으로 표식하고 3' 말단외두잔기가쌀라져서짧아지는 oli
gonucleotide 의크기변화를관측함으로써 융합단백 질외 integrase 활성을측정하였다 (Fig. 4A). 러트 로바이러스 integrase의조효소로알려진 망간과마 그네슘의 농도를 변화시키면서 integrase외 endo
nucleolytic 활성을 조사하였을 때. 0.1 — 1.0 mM 에서는 상당한 활성을 나타내었으나. 의 경우 어떠한 농도에서도 활성을 보이지 못하고있다 (Fig 4B: lanes 4-11). 이결과는융합단백질의 in-
A
TGTGGAAAATCTCTAGCAGT ACACCTTTTAGAGATCGTCA
IN
TGTGGAAAATCTCTAGCA-oh
ACACCTTTTAGAGATCGTCA + GT-oh
2 3 4 5 6 7 8 9 10 11
tegrase 활성은 망간을조효소로 필요로하여. his- tidine표식된 integrase와 같은 성질을 보여주고있 다."하또한 10 mM ■"또는 Mg우■"에서는 oligonu- cleotide의 잔기듈이 하나썩 쌀려짐으로써. inte- grase의활성이억제된가운데섞여있는 nuclease의 활성이 작용함을 보여준다(Fig. 4B: lanes 7 and 11). 이러한비특이적인 절단은본연구에 이용된 in-
tegrase들이한단계과정만으로정제되어박테라아에
서 유래된 nuclease가 integrase시료속에 포함되어 나타나는 것으로사료된다. 융합단백질의 integrase 활성의 적합한발현조건을조사하기 위하여활성측 정반응에 P-mercaptoethanol, CaCla,또는 EDTA 등을 첨가해 보았을 때. p-mercaptoethan이을 20
mM 수준까지 첨가한경우가더좋은활성을보여준다
(Fig. 5: lanes 5-7). 최중농도로 1 mM EDTA의첨 가는 부분적 활성을 보여주며, 그이상의 농도에서는 반응용액중의망간이온을완전히 제거하여 반응을억 제한다.
러트로바이러스 integrase는끝에두개외 nucleo- tide가제거된 바이러스 DNA를숙주세포의 DNA나 자신의바이러스 DNA외비특이적부위를끊고연결시
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3
#
• • •
• - •
융합단백질의 일부로써의 인테그라제 특성 51
키는 중합활성이 있다. In uiYro에서 이활성은 en
donucleolytic 활성에의하여두개외 nucleotide가제 거되고, 방사능으로 표식된 oligonucleotide를 다른 oligonucleotide의중간부위에 비톡이적으로끊고. 연 결함으로써 방사능으로표식된 oligonucleotide의크 기가증가하는것으로측정할수있다. 맥아당결합단백 질과의 융함단백질로존재하는 integrase는 histidi- ne으로표식된 integrase에비하여 endonucleolytic 활성이뛰어나지는못하다. 그러나. 맥아당결합단백질 과의융합단백질을사용한반응들에서 DNA기질의 20 mer 위부분에중합된생성물이더강하게나타나므로. integrase외 endonucleolytic 활성에 외하여 절단된 oligonucleotide가다른 DNA에둘어가는중함활성은 융함단백질이더강한것으로사료된다(Fig. 5: lane 2 vs lanes 3. 5-7. and 11).
면역결핍바이러스의 integrase의효소활성은단백질 외중앙영역에서 나타난다. 그러나, N-말단영역이나, C-말단영역을부분적으로제거한돌연변이들도효소활 성을쉽게상실하여 전체적인단백질외 구조가중요하 다16,19) 이것은효소활성이두개이상외 integrase가 결합된중함체에서만나타나기때문으로. 중합체외 형 성을위한단백질사이외결합은 N-말단영역을통하여 일어난다.^^ 따라서. 융합단백질이나 histidine이표식 된 integrase의경우중합체 형성을위한 N-말단구조 외순수성이 펄요하다. 현재많이사용되는 histidine 으로표식된 integrase의경우 N-말단에 12^20개정 도의아미노산이 추가되나. 효소활성의 발현에는거의 영향을주지않고있다."" "® ""* 분자량이 42,700 Da인맥 아당결합단백질이 N-말단에결합된 integrase의경우 정상적인중합체의형성이어려울것으로사료되나. 부 분적인 효소활성을보이는 것으로보아융합단백질외 구조에많은흥미를갖게한다. 또한, 이러한결과는 in- tegrase가 감염된 바이러스에서 생성될 때 polyp-
rotein의일부로생성되어바이러스조럽과정에단백분
해효소에외하여절단되어활성을갖는것으로알려지 고있으나. polyprotein외형태에서나. 부분적으로절단 된형태에서도활성을나타낼수도있옴을시사한다.
감 사 의 말 씀
본연구는 1996년도중앙대학교학술연구지원버에외
하여이루어졌기에이에감사드럽니다.
문 헌
1) Brown. P. O. : Integration of retroviral D N A . Curr. Topics Microbiol Im m unol. 157, 19 (1990).
2) Bushm an, F. D.. Fugiwara, T. and Craigie, R. ■ R e tro v iral D N A in te g ratio n directed by i n tegration protein in vitro. Science 249, 1555 (1990).
3) Craigie, R.. Fugiwara, T. and B ushm an , F. D. '■
The IN protein of M oloney m u rin e leukem ia virus processes the viral D N A ends a n d ac
complishes their integration in vitro. Cell 62, 829 (1990).
4) Katz, R. A.. M erkel G. W .. Kulkosky, J., L^is, J.
and Skalka. A. M. : The avian retroviral IN pro
tein is both necessary a n d sufficient for in tegrative recombination in vitro. Cell 63, 87 (1990).
5) V ink. C.. v an G ant. D. C., Elgersma, Y. and Plasterk. R. H . A. '■ H u m a n immunodeficiency virus integrase protein requires a subterm inal position of its viral D N A recognition sequence for efficient cleavage. I Virol 65. 4636 (1991).
6) B ushm an. F. D. and Craigie, R. : Activities of h u m a n im m un ode ficien cy v iru s (H IV ) in te gration protein in vitro: Specific cleavage an d integration of H IV D N A . Proc. N a tl Acad. Sci.
U SA 88. 1339 (1991).
7) Sherm an. P. A. and Fyffe. J . A. ■ H u m a n im m unodeficiency v irus integration protein ex
pressed in Escherichia coli possesses selective D N A cleaving activity. Proc. N atl. Acad. Sci.
U SA 87. 5119 (1990).
8) LaFemina, R. L.. Callahan. P. L. and Cordingley.
M. G. ■ Substrate specificity of recombinant H u m an immunodeficiency virus integrase protein. /.
Virol. 65. 5624 (1991).
9) Kulkosky, J.. Jones. K. S., Katz, R. A., Mack. J.
P. G. and Skalka. A. M. • Residues critical for re
troviral integrative recom bination in a region th a t is highly conserved am ong retroviral/re- trotransposon integrases and bacterial sequence transposases. M o l Cell Biol. 12. 2331 (1992).
10) Vink, C., Groeneger, A. A. M . an d Plasterk. R.
H . A. ■ Identification of the catalytic and DNA- binding region of the h u m a n immunodeficiency virus type 1 integrase protein. Nucleic A cid Res.
21 1419 (1993).
11) V ink, C. and Plasterk, R. H. A. ■ Activities of fe
line immunodeficiency virus integrase protein produced in Escherichia coli. }. Virol 68, 1468 (1994).
12) Jenkins. T. M., Hickman. A. B.. Dyda. F.. Ghir- lando, R .. Davies. D. R. and Craigie, R. : C a
talytic domain of h u m an iiimiunodeficiency virus type 1 integrase: Identification of soluble m utant by systematic replacement of hydrophobic resi
dues. Proc.. N a tl Acad. Sci. U SA 92. 6057 (1995).
13) O h . J.-W . and Shin. C.-G. '■ Purification and characterization of the H um an immunodeficiency virus type 1 integrase expressed in Escherichia coli M o l Cells. 6, 96 (1996).
14) K ellerm an , T. C. an d Ferenci, T. : M altose b in d in g protein from E. coli. M ethods in E n - zymology 90. 459 (1982).
15) G uan, C.. Li. P.. Riggs. P. D. and Inouye. H. ; Vectors th a t facilitate the expression and p u rification of foreign peptides in Escherichia coli by fusion to maltose-binding protein. Gene 67. 21
(1987).
16) B u sh m an . F. D .. E ngelm an. A .. Palmer. L.
Wingfield. P. and Craigie. R. : Domains of the integrase protein of h u m an immunodeficiency v irus type 1 responsible for polynucleotidyl transfer and zinc binding. Proc. Natl. Acad. Sci.
U SA 90. 3428 (1993).
17) K atzm an. M. and S u d o l M. - In vitro activities of purified V isna virus integrase. J. Virol. 68.
3558 (1994).
18) Balakrishnan, M .. Zastrow, D. and Jonsson. C.
B. : C atalytic activities of the h u m a n T-cell leukem ia virus type II integrase. Virology 219, 77 (1996).
19) Drelich. M .. W ilhelm , r. and Mous, J. : Iden
tification of am ino acid residues critical for en
donuclease and integration activities of HIV-1 IN protein in vitro. Virology 188. 459 (1992).
20) Jones. K. S.. Coleman. J .. Merkel, G. W., Laue.
T. M . and Skalka, A. M . : Retroviral integrase functions as a m ultim er and can tu rn over ca- talytically. /. Biol Chem. 267, 16037 (1992).
/. Pharm. Soc. Korea