CRISPR-Cas9 system 을 이용한 Kluyveromyces marxianus 17694-DH2 균주의 자일로스 소비속도 증진
권덕호1, 박중희1, 정덕열2, 박재범1, 박동민1, 강경곤1, 최서영1, 김수린2, 하석진1*
Application of Genome Editing Method on Kluyveromyces marxianus 17694-DH2 using CRISPR-Cas9 System for Enhanced Xylose Utilization
Deok-Ho Kwon1, Joong-Hee Park1, Deok Yeol Jeong2, Jae-Bum Park1, Dong-Min Park1, Kyoung-Gon Kang1, Seo-Young Choi1, Soo Rin Kim2, and Suk-Jin Ha1*
Received: 4 October 2019 / Revised: 8 November 2019 / Accepted: 10 November 2019
© 2019 The Korean Society for Biotechnology and Bioengineering
Abstract: The third-generation gene editing technology, CRISPR-Cas9 system, is derived from the bacterial immune system. These CRISPR-Cas9 systems have recently been used to mutate yeast gene or replace cleavage sites with other DNA. In this study, CRISPR-Cas9 system was applied to delete PHO13 gene of Kluyveromyces marxianus. As a result, only one strain of three transformants, that the CRISPR-Cas9 sys- tem was applied, was confirmed partial deletion of PHO13 gene. Sequencing of the PHO13 gene revealed the deletion of cytosine at position 457, which resulted in premature termina- tion of translation as compared to that from the parental strain.
The ΔPHO13 strain, K. marxianus 17694-DH2, showed 18.29%
and 21.28% improvement in xylose consumption and ethanol production from xylose, respectively, as compared to those form the parental strain.
Keywords: Kluyveromyces marxianus, CRISPR-Cas9, xylose, PHO13
1. INTRODUCTION
Kluyveromyces marxianus 균주는 잘 알려진 열내성 효모로써, 일반적인 효모가 생육하기 어려운 45
oC의 높은 온도에서도 비교적 성장속도가 빠르다는 장점을 가지고 있다 [1,2]. 이러 한 특징을 가진 K. marxianus는 일반적으로 polyploid 균주로 써 유전자 조작이 다소 어렵다는 단점을 가지고 있다 [3,5].
하지만 형질전환을 위해 삽입하는 DNA 조각들을 무작위적 으로 융합하는 특성을 가지고 있어 특정 유전자의 과발현을 통한 균주 개량을 할 수 있다. 또한 유도적 돌연변이 유발 방 법(directed evolutionary approach)을 이용하여 K. marxianus 36907-FMEL1 균주는 자일로스로부터 자일리톨 생산 능력 이 향상되었으며, K. marxianus SBK1은 글루코스와 자일로 스의 동시 발효를 가능하게되었다 [6,7]. 또한 K. marxianus 17555-JBP2 균주는 mKmXYL1 유전자를 과발현하여 자일리 톨 생산량, 수율, 생산성이 각각 440%, 135%, 440% 증가하 였다 [8]. 그리고 K. marxianus DMB13균주는 KmXYL1, KmXYL3 와 mKmXYL2를 무작위적 융합 방법으로 발현하여 이전 연 구에서 자일로스 대사능력을 향상시킨 DMB3-7 균주와 40
oC 조건에서 비교했을 때 자일로스 소비속도가 320% 증 가하였다 [9,10]. 이와 같은 방법으로 과발현 또는 유도적 돌 연변이를 유도하는 것은 가능하지만 K. marxianus 균주의 non-homologous end-joining (NHEJ) 때문에 특정 유전자의 삭제 또는 homologous recombination (HR)을 사용한 특정 위 치의 유전자 조작을 하기에는 어려움이 따른다 [3].
최근 유전자 조작 기술로 각광을 받고 있는 3세대 유전자
1강원대학교 생물공학과
1Department of Bioengineering and Technology, Kangwon National University, Chuncheon, Korea
Tel: +82-33-250-6278, Fax: +82-33-243-6350 e-mail: [email protected]
2경북대학교 식품공학부
2School of Food Science and Biotechnology, Kyungpook National University, Daegu 41566, Korea
Research Paper
조작 기술인 clustered regularly interspaced short palindromic repeat (CRISPR)-Cas9 system은 박테리아의 면역 체계에서 발견되었으며, 유전자의 변형을 일으키거나 절단 부위를 다 른 DNA로 대체하기 위해 사용되고 있다 [11,13]. 이전 연구 에서 에틸 아세테이트와 에탄올의 생합성을 이해하기 위해 K. marxianus CBS6556 균주에 CRISPR-Cas9 system을 이용 하여 KmAdh1~7과 KmATF 유전자를 각각 부분 결실시켜 단 백질을 비활성화 하였다 [14]. 그 결과 KmAdh2 유전자가 호 기 또는 혐기 조건에서 에탄올 생산에 중요한 역할을 한다는 것과 alcohol-O-acetyltransferase의 비활성화가 에틸 아세테 이트 생산을 완전히 억제하지 못하는 것을 확인하였다. 또한 K. marxianus 균주의 NHEJ 특성을 억제하기 위해 NHEJ 단 백질의 유전자인 Nej1 또는 Dnl4를 CRISPR-Cas9 system으 로 돌연변이를 만들어 NHEJ의 활성을 억제하고 HR의 효과 를 증진시킨 연구가 있다 [3].
자일로스를 이용할 수 없는 Saccharomyces cerevisiae 균주 에 자일로스의 초기 대사경로인 xylose reductase, xylitol dehydrogenase, xylulose kinase)를 도입하기 위해 XYL1, XYL2, XYL3 유전자를 발현한 후 PHO13 유전자를 삭제하였 을 때 자일로스 소비 속도가 증가하는 것이 확인되었다 [15].
따라서 본 연구에서는 자일로스로부터 에탄올 생산이 증가 된 돌연변이 균주 K. marxianus 17694-DH1을 모균주로 하여 KmPHO13 유전자를 CRISPR-Cas9 system으로 제거하여 자 일로스 소비량 증가와 에탄올 생산 효율 향상을 확인하였다.
2. MATERIALS AND METHOD
2.1. 균주 및 발효 조건
본 연구에 모균주로 사용된 균주는 이전 연구에서 개량한 유 도적 돌연변이 방법으로 개량된 돌연변이 균주 K. marxianus 17694-DH1 이며, 자일로스로부터 에탄올 생산이 향상된 균 주이다 [16]. 전 배양은 YPD (Yeast extract 10 g/L, Peptone 20 g/L, Dextrose 20 g/L) 배지를 사용하여 30
oC 배양 온도와 200 rpm 교반 조건에서 진행하였다. 자일로스 발효 결과를 비교하기 위한 본 배양은 YPX (Yeast extract 10 g/L, Peptone 20 g/L, Xylose 80 g/L) 배지에서 수행하였으며 30
oC 배양 온 도조건에서 교반속도를 100 rpm과 200 rpm의 두 가지 조건 으로 달리하여 발효를 수행하여 비교하였다.
2.2. CRISPR-Cas9 system 의 적용과 형질전환 균주의 선 별 방법
K. marxianus 17694-DH1 균주에 CRISPR-Cas9 system을 이 용하기 위한 Cas9 단백질의 발현은 single-copy plasmid인 pRS41N vector (p414-TEF1p-Cas9-CYC1t)를 이용하였으며 selection marker로 clonNAT을 사용하였다. 또한 특정 유전자 인식을 위해서 multi-copy plasmid이며 gBlock과 gRNA (NGG와 PAM 염기서열 포함)가 포함된 pRS42H vector를 이 용하였고 selection marker로 hygromycin B를 사용하였다
[15]. pRS42H vector는 PHO13 유전자의 특정 염기서열에 결 합하여야 하므로 overlap PCR 방법을 통해 제작되었으며 overlap PCR에 사용된 primer는 다음과 같다. gRNA-PHO13- F (5 ′-CAATACTTCGCCATTCCTTAGTTTTAGAGCTAGAA ATA GCA AG-3′), gRNA-PHO13-R (5′-TAAGGAATGGCG AAGTATTGGATCATTTATCTTTCACTGCG-3′).
형질전환에 사용될 모균주는 30
oC 배양 온도와 200 rpm 교 반조건에서 YPD 배지를 사용하여 배양하였으며, 접종 후 흡 광도를 측정하여 OD
6003.0 이 되었을 때, 각각의 pRS41N vector와 pRS42H vector를 poly ethylene glycol (PEG3350;
Sigma, St. Louis, MO)과 lithium acetate (Sigma, St. Louis, MO) 그리고 carrier DNA를 이용한 방법으로 transformation 을 실시하였다 [17]. 균주에 vector를 삽입한 후 clonNAT (100 ug/mL) 과 hygromycin B (600 µg/mL)를 첨가된 YPD agar 배지를 이용하여 30
oC 온도조건에서 배양하여 형질전 환체 균주를 선별하였다.
2.2. KmPHO13 유전자의 염기서열 분석
CRISPR-Cas9 system 을 이용하여 선별된 3개의 형질전환 균 주의 genomic DNA를 각각 추출한 후 KmPHO13_F (5′- ATGACTGGTACGAAAGCGAAT-3′)와 KmPHO13_R (5′- CTATTGATGGGTGAATTCATAAAGGTC-3 ′) primer를 사용 하여 KmPHO13 유전자를 증폭하기 위한 PCR을 수행한 후 T-vector 를 이용하여 클로닝을 수행하였다. 이 후 각 균주의 KmPHO13 유전자가 삽입된 T-vector를 이용하여 유전자 염 기서열을 분석하였다. 유전자 염기서열 분석은 ㈜마크로젠 (Seoul, Korea) 에 의뢰하였으며 염기서열 분석 프로그램인 Lasergene (DNASTAR, Inc., WI, USA)을 사용하여 분석하였다.
2.3. 형질전환 균주의 성장 속도 및 대사산물 분석 효모의 성장은 24시간 간격으로 일정한 양을 취해 흡광광도 계 (Biomate 5, Thermo, NY, USA)를 사용하여 600 nm 파장 에서 흡광도를 측정하여 분석하였다. 효모의 대사 산물인 자 일로스, 자일리톨, 에탄올 등은 고성능 액체크로마토그래피 (HPLC 1200 series, Agilent Technologies, Santa Clara, CA, USA)를 이용하여 분석하였으며, 컬럼은 Rezex ROA-Organic acid H+ (Phenomenex, Torrance, CA, USA) 를 사용하였다. 컬 럼과 refractive index detector의 온도는 50℃를 유지하였으며, 0.6 mL/min의 유속으로 0.005 N의 황산용액을 이동상으로 사용하였다.
3. RESULTS AND DISCUSSION
3.1. 다양한 K. marxianus 균주의 항생제 clonNAT과 hygromycin B 에 대한 내성 확인
유전자 조작 기술인 CRISPR-Cas9 system을 K. marxianus 균
주에 사용하기 위해서 pRS41N vector와 pRS42H vector의
selection marker로 사용되는 clonNAT과 hygromycin B에 대
한 내성 유 · 무를 확인하였다. 항생제 내성을 확인하기 위해 서 100 µg/mL의 clonNAT과 600 µg/mL의 hygromycin B가 첨가된 YPD agar 배지를 각각 준비한 후 3종류의 K. marxianus 균주 (K. marxianus KCTC17555, K. marxianus KCTC17694, K. marxianus ATCC63907)를 평판 도말법을 사용하여 접종 하였다. 각각의 항생제 clonNAT과 hygromycin B가 첨가된 YPD agar 배지에서 K. marxianus KCTC17555 균주와 K.
marxianus ATCC36907 균주는 정상적인 균주의 성장을 나타 내어 clonNAT과 hygromycin B에 대한 내성이 있는 것으로 확인되었으나, K. marxianus KCTC17694 균주는 전혀 성장 하지 못하여 각각의 항생제에 대한 내성이 없는 것으로 확인 되었다 (Table 1). 따라서 CRISPR-Cas9 system을 적용하여 형질전환체를 선별하기 위해서는 항생제에 대한 내성이 없 는 K. marxianus KCTC17694 균주만이 이용이 가능할 것으 로 확인되었다. 이후 K. marxianus KCTC17694를 모균주로 하여 유도적 돌연변이 방법을 통해 선별된 K. marxianus 17694- DH1 균주의 clonNAT과 hygromycin B에 대한 내성을 측정 한 결과 모균주와 같이 내성이 없는 것으로 확인이 되어 CRISPR-Cas9 system을 적용하기 위한 실험은 K. marxianus 17694-DH1을 모균주로 하여 실험을 진행하였다.
3.2. CRISPR-Cas9 system 을 이용한 K. marxianus 17694- DH1 균주의 KmPHO13 유전자 편집
CRISPR-Cas9 system은 Cas9 단백질과 single guide RNA (sgRNA)로 이루어져 있으며 유전 정보를 바탕으로 gRNA가 protospacer adjacent motif (PAM) site를 기준으로 5’ 방향으 로 17~20개 정도의 염기서열을 인식하도록 디자인하게 되면 Cas9 단백질이 PAM site를 기준으로 3~5번째 염기서열의
DNA를 절단하게 된다 [18-20]. DNA가 절단되면 이를 스스 로 복구하기 위해 NHEJ 또는 homology-directed repair (HDR) 방법을 사용하여 절단 부위를 고치게 되는데 이때 유 전자가 변형 되거나 다른 DNA로 교체 될 수 있다 [21]. 이러 한 특징으로 인해 CRISPR-Cas9 system은 S. cerevisiae, Candida glabrata, K. marxianus 등의 여러 효모의 유전자 조작에 응용 되고 있다 [3,22,23]. K. marxianus 17694-DH1 균주의 KmPHO13 유전자 조작을 위해 pRS41N vector를 도입한 이후 clonNAT 이 100 µg/mL이 포함된 YPD agar 배지에서 선별하였으며 KmPHO13 유전자에 gRNA가 특이적으로 결합하도록 개량 된 pRS42H vector를 추가로 도입하여 clonNAT 100 µg/mL와 hygromycin B 600 µg/mL이 동시에 첨가된 YPD agar 배지에 서 최종 선별하였다.
실험 결과 총 3개의 colony가 선별되었으며, clonNAT 100 µg/
mL와 hygromycin B 600 µg/mL를 함유한 YPD 배지에서 24 시간씩 5번 연속적으로 계대배양하여 최종적으로 균주를 선 별하였다. 선별된 3개의 균주와 모균주의 KmPHO13 유전자 의 염기서열을 비교한 결과 3개의 균주 중 한 개의 균주만이 KmPHO13 유전자에 결실이 있는 것을 확인하였으며 유전자 조작이 성공한 K. marxianus 17694-DH1-ΔKmPHO13-2 균주 를 K. marxianus 17694-DH2로 명명하였다. K. marxianus 17694-DH2 균주의 KmPHO13 유전자 결실 부위는 PAM site 로부터 4번째 위치한 시토신이며 전체 염기서열에서 457번 째 위치한 것을 확인하였다 (Fig. 1). 또한 시토신의 결실로 인하여 KmPHO13의 154번째 아미노산인 라이신이 아르기 닌으로 치환되고 168번째에 아미노산에 종결코돈이 생김으 로 인하여 KmPHO13 단백질이 본래의 활성을 가지지 못할 것으로 예상된다.
3.3. 모균주와 K. marxianus 17694-DH2 균주의 자일로스 소비 속도 비교
유전자 조작 기술로 선별된 균주의 자일로스 소비 속도를 비 교하기 위하여 모균주 (K. marxianus 17694-DH1)와 K. marxianus 17694-DH2 균주의 발효를 30
oC 온도 조건에서 100 rpm과 200 rpm의 교반속도 조건으로 각각 나누어 발효를 수행하였 다. 자일로스가 80 g/L 포함된 YP 배지에서 교반속도를 100 rpm 으로 유지하고 발효를 진행한 결과, 모균주의 경우 엔 발효 168시간에 자일로스를 53.98 ± 1.89 g/L 소비하여 에 Table 1. Evaluation of antibiotics resistances against ClonNAT or
Hygromycin B from three K. marxianus strains
Strains Cell growth
ClonNAT (100 μg/mL) Hygromycin B (600 μg/mL) K. marxianus
KCTC17555 О О
K. marxianus
KCTC17694 Х Х
K. marxianus
ATCC36907 О О
Fig. 1. Alignment of nucleotide sequences of KmPHO13 from K. marxianus 17694-DH1, K. marxianus 17694-DH1ΔKmPHO13-1, K.
marxianus 17694-DH1ΔKmPHO13-2 and K. marxianus 17694-DH1ΔKmPHO13-3.
탄올을 14.54 ± 0.20 g/L 생산하였으며 이때 에탄올의 수율이 0.26 g/g임을 확인하였다 (Fig. 2(A)). K. marxianus 17694- DH2 의 경우엔 발효 216시간에 자일로스를 64.26 ± 1.17 g/L 소비하여 에탄올을 16.08 ± 0.50 g/L 생산하였으며 에탄올의 수율이 0.25 g/g임을 확인하였다 (Fig. 2(B)). 따라서 자일로 스 소비량과 에탄올 생산량이 각각 약 19% 그리고 10% 증가 하였지만 최대 에탄올 생산을 위한 발효시간이 168시간에서 216 시간으로 증가함에 따라 생산 속도가 크게 낮아지는 결 과를 확인하였다. 이러한 결과에 따르면 100 rpm의 비교적 낮은 교반조건에서 형질전환체 세포의 생장 속도가 모균주 보다 느린 것으로 판단되어 이후 균주의 성장 속도를 높이기 위해 보다 호기적인 조건에서 실험을 수행하였다.
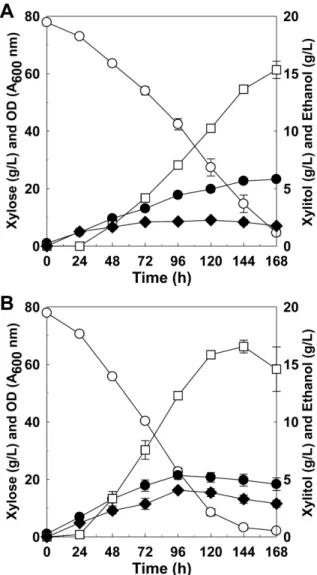
자일로스가 80 g/L 포함된 YP 배지에서 교반속도를 200 rpm 으로 유지하고 발효를 수행한 결과, 모균주의 경우 144시간 에 63.14 ± 3.07 g/L의 자일로스를 소비하고 13.65 ± 0.28 g/L 의 에탄올을 생산하였으며, 에탄올 수율과 생산 속도는 각각 0.21 g/g과 0.09 g/L·h였다 (Fig. 3(A)). 반면에 K. marxianus 17694-DH2 균주는 144시간에 74.69 ± 0.78 g/L의 자일로스 를 소비하여 16.55 ± 0.55 g/L의 에탄올을 생산하였으며, 에 탄올 수율과 생산성은 각각 0.22 g/g과 0.11 g/L·h 였다. 모균 주와 비교하여 자일로스 소비량은 18.29%, 에탄올 생산량은 21.28% 증가한 것을 확인하였다 (Fig. 3(B)).
100 rpm과 200 rpm의 교반조건에서 각각 발효를 수행한 결 과 비교적 호기적인 조건인 200 rpm 교반조건에서 KmPHO13 유전자를 조작한 균주 (K. marxianus 17694-DH2)의 자일로 스 대사 능력이 증진된 것을 확인하였다. 이전 연구에서 PHO13 유전자가 자일로스 대사 과정에서 xyloulose-5-phosphate 의 탈인산화에 관여하여 xylulose kinase에 의한 xylulose 인 산화 과정이 반복되는 대사 에너지의 낭비를 일으킬 수 있다 는 연구결과 [15,24]가 있으며, 또한 PHO13 유전자 결실에 의해 호기적인 조건에서 NADH 생산이 향상된다는 연구결 과가 있다 [25]. 따라서 KmPHO13 유전자가 조작된 K.
marxianus 17694-DH2 균주는 대사 에너지의 낭비를 줄이고
호기적인 조건에서 자일로스 대사에 필요한 co-factor (NADH)의 재생이 유리하므로 모균주에 비해 자일로스 소비 와 에탄올 생산 결과가 크게 향상되었다고 사료된다.
4. CONCLUSION
K. marxianus 균주에 CRISPR-Cas9 systems을 적용하여 PHO13 유전자의 편집에 성공하였으며, 형질전환에 성공한 K. marxianus 17694-DH2 균주를 모균주와 비교 분석하였다.
형질전환된 균주의 자일로스 소비량과 에탄올 생산량은 200 rpm 교반조건에서 모균주와 비교하여 각각 18.29%, 21.28% 향상됨을 확인하였으며, 이는 PHO13 유전자의 결 실로 인한 자일로스를 대사하는 능력이 향상된 것으로 사료 된다.
Fig. 2. Comparisons of xylose fermentation results by K. marxianus 17694-DH1 and K. marxianus 17694-DH2 using YP medium containing 80 g/L xylose at 30
oC with 100 rpm. Symbols: xylose ( ■ ), ethanol ( □ ) and ethanol yield (▨).
Fig. 3. Time profiles of xylose fermentation by K. marxianus 17694-DH1 (A) and K. marxianus 17694-DH2 (B) in YP medium containing 80 g/L xylose at 30oC with 200 rpm. Symbols: xylose (○), xylitol (
◆
), optical density (●
) and ethanol (□
).Acknowledgements
이 논문은 2019년 대학혁신지원사업 도전 연구비지원 프로 그램의 지원을 받아 수행됨. 2018년도 정부(교육부)의 재원 으로 한국연구재단의 지원을 받아 수행된 기초연구사업임 (No. 2018R1A6A1A03025582).
REFERENCES
1. Anderson, P., K. McNeil, and K. Watson (1986) High-efficiency carbohydrate fermentation to ethanol at temperatures above 40 C by Kluyveromyces marxianus var. marxianus isolated from sugar mills. Appl. Environ. Microbiol. 51: 1314-1320.
2. Hughes, D., N. Tudroszen, and C. J. Moye (1984) The effect of temperature on the kinetics of ethanol production by a thermotoler- ant strain of Kluveromyces marxianus. Biotechnology letters. 6: 1-6.
3. Nambu-Nishida, Y., K. Nishida, T. Hasunuma, and A. Kondo (2017) Development of a comprehensive set of tools for genome engineering in a cold-and thermo-tolerant Kluyveromyces marx- ianus yeast strain. Scientific reports. 7: 8993.
4. Hoshida, H., N. Murakami, A. Suzuki, R. Tamura, J. Asakawa, B.
M. Abdel‐Banat, S. Nonklang, M. Nakamura, and R. Akada (2014) Non‐homologous end joining‐mediated functional marker selection for DNA cloning in the yeast Kluyveromyces marx- ianus. Yeast. 31: 29-46.
5. Ribeiro, O., A. K. Gombert, J. A. Teixeira, and L. Domingues (2007) Application of the Cre-loxP system for multiple gene dis- ruption in the yeast Kluyveromyces marxianus. Journal of biotech- nology. 131: 20-26.
6. Kim, S.-B., D.-H. Kwon, J.-B. Park, and S.-J. Ha (2019) Allevia- tion of catabolite repression in Kluyveromyces marxianus: the thermotolerant SBK1 mutant simultaneously coferments glucose and xylose. Biotechnology for biofuels. 12: 90.
7. Kim, J.-S., J.-B. Park, S.-W. Jang, and S.-J. Ha (2015) Enhanced xylitol production by mutant Kluyveromyces marxianus 36907- FMEL1 due to improved xylose reductase activity. Applied bio- chemistry and biotechnology. 176: 1975-1984.
8. Park, J.-B., J.-S. Kim, D.-H. Kweon, D.-H. Kweon, J.-H. Seo, and S.-J. Ha (2019) Overexpression of Endogenous Xylose Reductase Enhanced Xylitol Productivity at 40°C by Thermotolerant Yeast Kluyveromyces marxianus. Applied biochemistry and biotechnol- ogy. 1-12.
9. Suzuki, T., T. Hoshino, and A. Matsushika (2019) High-tempera- ture ethanol production by a series of recombinant xylose-ferment- ing Kluyveromyces marxianus strains. Enzyme and Microbial Technology. 109359.
10. Goshima, T., K. Negi, M. Tsuji, H. Inoue, S. Yano, T. Hoshino, and A. Matsushika (2013) Ethanol fermentation from xylose by metabolically engineered strains of Kluyveromyces marxianus.
Journal of bioscience and bioengineering. 116: 551-554.
11. Brouns, S. J., M. M. Jore, M. Lundgren, E. R. Westra, R. J. Sli- jkhuis, A. P. Snijders, M. J. Dickman, K. S. Makarova, E. V. Koo- nin, and J. Van Der Oost (2008) Small CRISPR RNAs guide
antiviral defense in prokaryotes. Science. 321: 960-964.
12. Guy, C. P., A. I. Majerník, J. P. Chong, and E. L. Bolt (2004) A novel nuclease-ATPase (Nar71) from archaea is part of a pro- posed thermophilic DNA repair system. Nucleic acids research.
32: 6176-6186.
13. Makarova, K. S., L. Aravind, N. V. Grishin, I. B. Rogozin, and E.
V. Koonin (2002) A DNA repair system specific for thermophilic Archaea and bacteria predicted by genomic context analysis.
Nucleic acids research. 30: 482-496.
14. Löbs, A.-K., R. Engel, C. Schwartz, A. Flores, and I. Wheeldon (2017) CRISPR–Cas9-enabled genetic disruptions for understand- ing ethanol and ethyl acetate biosynthesis in Kluyveromyces marx- ianus. Biotechnology for biofuels. 10: 164.
15. Kim, S. R., H. Xu, A. Lesmana, U. Kuzmanovic, M. Au, C. Flor- encia, E. J. Oh, G. Zhang, K. H. Kim, and Y.-S. Jin (2015) Dele- tion of PHO13, encoding haloacid dehalogenase type IIA phosphatase, results in upregulation of the pentose phosphate pathway in Saccharomyces cerevisiae. Appl. Environ. Microbiol.
81: 1601-1609.
16. Kwon, D.-H., J.-B. Park, E. Hong, and S.-J. Ha (2019) Ethanol production from xylose is highly increased by the Kluyveromyces marxianus mutant 17694-DH1. Bioprocess and biosystems engi- neering. 42: 63-70.
17. Schiestl, R. H., and R. D. Gietz (1989) High efficiency transforma- tion of intact yeast cells using single stranded nucleic acids as a carrier. Current genetics. 16: 339-346.
18. Sternberg, S. H., S. Redding, M. Jinek, E. C. Greene, and J. A.
Doudna (2014) DNA interrogation by the CRISPR RNA-guided endonuclease Cas9. Nature. 507: 62-67.
19. Anders, C., O. Niewoehner, A. Duerst, and M. Jinek (2014) Struc- tural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease. Nature. 513: 569-573.
20. Szczelkun, M. D., M. S. Tikhomirova, T. Sinkunas, G. Gasiunas, T.
Karvelis, P. Pschera, V. Siksnys, and R. Seidel (2014) Direct obser- vation of R-loop formation by single RNA-guided Cas9 and Cas- cade effector complexes. Proceedings of the National Academy of Sciences. 111: 9798-9803.
21. Doudna, J. A., and E. Charpentier (2014) The new frontier of genome engineering with CRISPR-Cas9. Science. 346: 1258096.
22. Enkler, L., D. Richer, A. L. Marchand, D. Ferrandon, and F. Jossi- net (2016) Genome engineering in the yeast pathogen Candida glabrata using the CRISPR-Cas9 system. Scientific reports. 6:
35766.
23. Tsai, C. S., I. I. Kong, A. Lesmana, G. Million, G. C. Zhang, S. R.
Kim, and Y. S. Jin (2015) Rapid and marker‐free refactoring of xylose‐fermenting yeast strains with Cas9/CRISPR. Biotechnol- ogy and bioengineering. 112: 2406-2411.
24. Kim, S. R., J. M. Skerker, W. Kang, A. Lesmana, N. Wei, A. P.
Arkin, and Y.-S. Jin (2013) Rational and evolutionary engineering approaches uncover a small set of genetic changes efficient for rapid xylose fermentation in Saccharomyces cerevisiae. PloS one.
8: e57048.
25. Bommer, G. T., F. Baldin, and E. Van Schaftingen (2016) Accumu- lation of metabolic side products might favor the production of ethanol in Pho13 knockout strains. Microbial Cell. 3: 495.