관련 문서
Spoilage potential of psychrotrophic lactic acid bacteria (LAB) species: Leuconostoc gelidum subsp. gasicomitatum and Lactococcus piscium , on sweet bell
In this study, we found that Pin1 plays a pivotal role in insulin- induced AP-1 activation through its interaction with p70S6K and prolongs activity of
This study derives the attributes of 'product design identity' from the consumer's point of view, and verified whether the 'product design identity'
The purpose and necessity of this study was to investigate the effects of lactic acid fatigue and body oxidation through myofascial relaxation exercise using small tools
In paragraph 2, this study defined the concept, meaning, and educational significance of art education using play activity, defined games in creativity
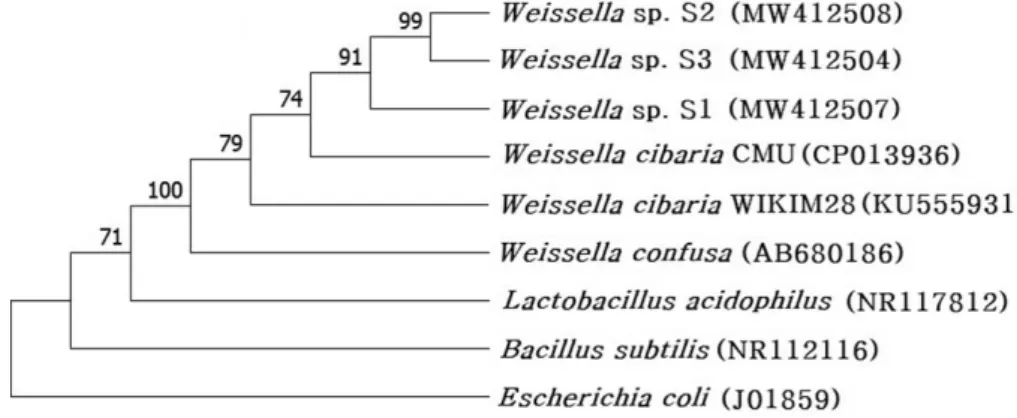
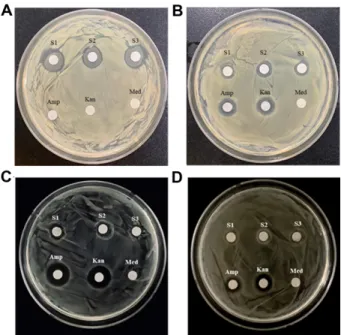
Probiotic properties of lactic acid bacteria isolated from mukeunji, a long-term ripened kimchi. Journal of Food
The object of this study is to analyse the bacteria of periapical bascess and fascial space abscess using multiplex real-time polymerase chain reaction (MRT-PCR) before
The obligate methanotrophic bacteria Methylococcus, Methylomonas, Methylosinus and related bacteria. Schleifer