ISSN: 2233-601X (Print) ISSN: 2093-6516 (Online)
Received: August 5, 2016, Revised: October 5, 2016, Accepted: October 17, 2016, Published online: June 5, 2017
Corresponding author: Jong In Kim, Department of Thoracic and Cardiovascular Surgery, Kosin University Gospel Hospital, Kosin University College of Medicine, 262 Gamcheon-ro, Seo-gu, Busan 49267, Korea
(Tel) 82-51-990-6466 (Fax) 82-51-990-3066 (E-mail) [email protected]
© The Korean Society for Thoracic and Cardiovascular Surgery. 2017. All right reserved.
This is an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/
licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
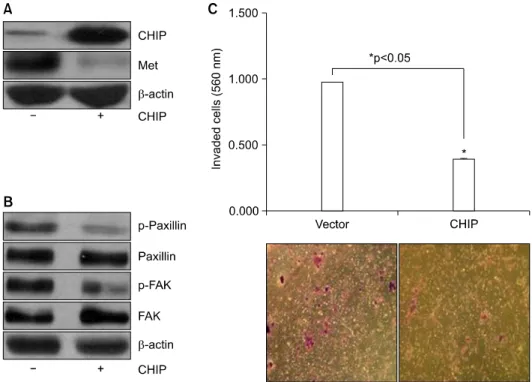
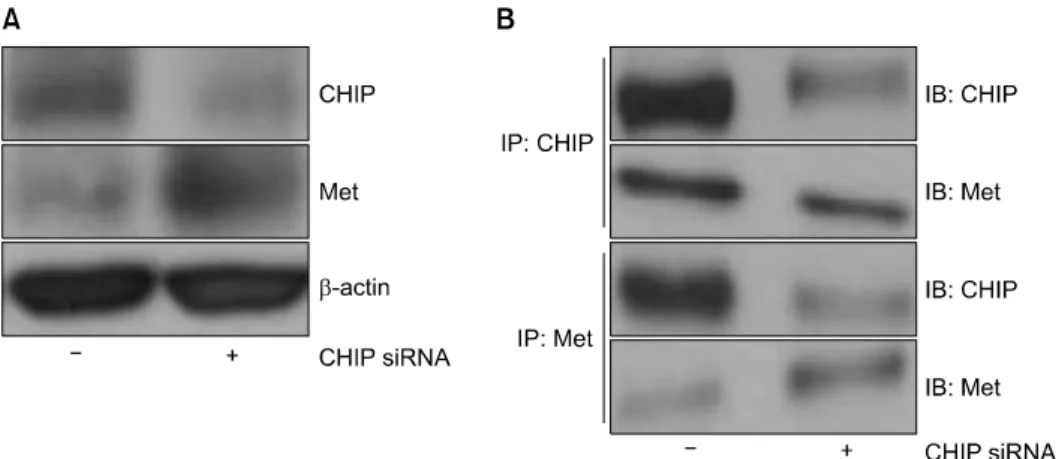
The Antitumor Effect of C-terminus of Hsp70-Interacting Protein via Degradation of c-Met in Small Cell Lung Cancer
Sung Ho Cho, M.D. 1 , Jong In Kim, M.D. 1 , Hyun Su Kim, M.D. 1 , Sung Dal Park, M.D. 1 , Kang Won Jang, Ph.D. 2
1