Original Article: Biochemistry/Molecular Biology
A1E Induces Apoptosis via Targeting HPV E6/E7 Oncogenes and Intrinsic Pathways in Cervical Cancer Cells
Sun Young Ham · Ye Sol Bak · Tae Ho Kwon · Jeong Woo Kang · Kang Duk Choi · Tae Young Han · Il Young Han · Young Yang · Seung Hyun Jung · Do Young Yoon*
Received: 9 September 2013 / Accepted: 8 October 2013 / Published Online: 30 June 2014
© The Korean Society for Applied Biological Chemistry 2014
Abstract A1E is an extract from traditional Asian medicinal plants that has therapeutic activities against cancers, metabolic disease, and other intractable conditions. However, its mechanism of action on cervical cancer has not been studied. In order to ascertain if A1E would have pronounced anti-cervical cancer effect, cervical cancer cells were incubated with A1E and apoptosis was detected by nuclear morphological changes, annexin V-FITC/
PI staining, cell cycle analysis, western blotting, Reverse-transcription polymerase chain reaction, and measurement of mitochondrial membrane potential. Expression of human papiloma virus E6 and E7 oncogenes was down-regulated in A1E-treated cervical cancer cells, while p53 and retinoblastoma protein levels were enhanced.
A1E also perturbed cell cycle progression at sub-G1 and altered cell cycle regulatory factors in SiHa cervical cancer cells. A1E activated apoptotic intrinsic pathway markers such as caspase-9, caspase-3 and poly ADP-ribose polymerase, and down-regulated expression of Bcl-2 and Bcl-xl. A1E induced mitochondrial membrane potential collapse and cytochrome c release, and inhibited phosphatidylinositol 3-kinase (PI3K)/Akt, key factors involved in cell survival signaling. Taken all these results, A1E induced apoptosis via activation of the intrinsic pathway and inhibition of the PI3K/Akt survival-signaling pathway in SiHa cervical cancer cells. In conclusion, A1E exerts anti-proliferative action growth inhibition on cervical cancer cells through apoptosis which demonstrates its anti-cervical cancer properties.
Keywords apoptosis · cervical cancer · human papiloma virus · secondary metabolites · traditional medicinal plant
Introduction
Herbal medicines have been used to treat various diseases since ancient times in many Asian countries. Herbal medicines are extracted from traditional Asian medical plants and have therapeutic activities against cancers, angiogenesis, and metastasis in the absence of observable in vivo side effects (Lee et al., 2011).
It has been also reported that herbal formula inhibits cell proliferation and induces cell death via modulating the autophagy on human colon cancer cells (Yim et al., 2013). A1E is an extract from Asian traditional medicinal plants containing 11 herbal plants (Bak et al., 2013). Asian traditional medicines have been identified as an effective drug for improving function of the digestive system and for strengthening defenses against various infections in Asia (Kishida et al., 2007; Lee et al., 2012). One component of this medicine, ginseng, was reported to possess many anti-cancer effects, as well as enhancing immunity (Kiefer S. Y. Ham · Y. S. Bak · T. H. Kwon · J. W. Kang · D. Y. Yoon
Department of Bioscience and Biotechnology, Bio/Molecular Informatics Center, Konkuk University, Seoul, Republic of Korea
K. D. Choi
Graduate School of Bio & Information Technology, Hankyong National University, Ansung, Kyungki-do, Republic of Korea
T. Y. Han
Banryong Insu Herb Clinic, Nonhyun-dong, Gangnam-Gu, Seoul, Republic of Korea
I. Y. Han
Sunwun Biophysic, Ansung, Kyungki-do, Republic of Korea Y. Yang
Department of Biological Science, Sookmyoung Women’s University, Seoul, Republic of Korea
S. H. Jung
School of Oriental Medicine, Dongguk University, Siksa dong, Ilsan, Republic of Korea
*Corresponding author (D. Y. Yoon: [email protected])
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.
org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
and Pantuso, 2003). Chaga was also reported to exhibit anti- mutagenic and anti-cancer effects (Ham et al., 2009; Nakajima et al., 2009). In this study, we investigated whether A1E would have apoptotic effects on cervical cancer cells.
Cervical cancer is the second leading cause of female cancer deaths worldwide. Infection with high-risk human papilloma viruses (HPVs), such as HPV 16, 18, 31, and 33, play a central role in the development of almost 99.5% of cervical cancers (Munagala et al., 2011). HPV is implicated in virtually all cervical cancers worldwide, with HPV 16 being the most common high- risk type. The E6 and E7 oncoproteins have been shown to be the main mediators of the development of HPV-induced cervical carcinoma. High-risk E6 and E7 oncoproteins drive cell proliferation and contribute to neoplastic progression. E6-mediated p53 degradation prevents the normal repair of chance mutations in the cellular genome (Doorbar, 2006). The E7 protein associates with the retinoblastoma (RB) protein and interferes with the production of E2F/RB complex, that result lead to E2F activation and gene transcription (Sima et al., 2007). When expressions E6 and E7 genes are inhibited, tumor suppressor proteins such as retinoblastoma protein (pRb) and p53 are normally recovered.
Cell death takes 2 distinct forms: necrosis and apoptosis.
Necrosis is a degenerative phenomenon that follows irreversible injury. Apoptosis, in contrast, is an active process requiring protein synthesis for its execution (Walker et al., 1988). Apoptosis can easily be distinguished from necrosis through its diverse alterations to the cell, including chromatin condensation, DNA fragmentation, cytoplasmic shrinkage, and the formation of apoptotic bodies (Bak et al., 2011). In particular, the apoptotic intrinsic pathway involves a cascade of molecular events occurring entirely within cells. Mitochondria are central to the intrinsic apoptotic pathway.
On receipt of an apoptotic signal, a pro-apoptotic member of the B-cell lymphoma-2 (Bcl-2) family, such as Bcl-2-associated X (Bax), oligomerizes and inserts into the mitochondrial membrane, permeabilizing it and allowing cytochrome c to redistribute into the cytoplasm (Franklin, 2011). p53 can translocate to the cytosol and associate with mitochondria. Direct effects of p53 on proteins of the Bcl-2 family have been proposed (Lucken-Ardjomande et al., 2005). Intrinsic pathway-related p53 protein is also degraded by HPV E6 protein (Tan et al., 2012). The Phosphatidylinositide 3-kinases/Akt (PI3K/Akt) pathway can induce cell cycle progression by modulating the protein stability of cyclin D (Muise-Helmericks et al., 1998). Akt can phosphorylate pro-apoptotic Bad, and allow excess Bcl-2 or Bcl-XL to out-compete Bax (Chang et al., 2003).
In this study, we assessed the molecular mechanisms involved in the anti-cancer effects of A1E, an Asian medicinal plant extract.
We examined the role of the cytotoxic and apoptotic pathways in A1E activity in the cervical cancer cell line SiHa. We assessed the potential of A1E as a novel chemotherapeutic agent in human cervical cancer. To this end, we investigated the effects of A1E on apoptosis mediated via intrinsic pathways and on PI3K/Akt survival signaling pathways in SiHa cells.
Matrials and Methods
Reagents. CellTiter 96 AQueous One solution Cell Proliferation Assay Reagent (MTS; 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxy- methoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) was purchased from Promega (USA). Phenazine methosulfate (PMS), Hoechst stain solution and propidium iodide (PI) were purchased from Sigma (USA). An FITC-annexin V Apoptosis Detection Kit I was purchased from BD Biosciences (USA). Antibodies specific to cyclin D were purchased from BD Biosciences. Antibodies specific to poly ADP-ribose polymerase (PARP), caspase-3, caspase-9, caspase-8, Bcl-2, Bcl-xl, pRb, p53, p-p53, cytochrome c, and anti- mouse IgG-horseradish peroxidase were purchased from Cell Signaling Technology (USA). Antibodies specific to p21, p27, GAPDH, and anti-goat IgG- horseradish peroxidase (HRP) were from Santa Cruz Biotechnology (USA). A caspase-9 inhibitor (Z- IETD-fmk) was from R&D Systems (USA). qPCR reagent iQTM and SYBR® Green Supermix were both from Bio-Rad (USA). JC- 1 (5,5',6,6'-tetrachloro-1,1',3,3'-tetraethyl benzimidazolylcarbocy- anine iodide) was purchased from Enzo (USA).
Oriental medicinal herbs and formulation of A1E. A1E is composed of 11 oriental medicinal herbs. The 11 ingredients and their proportions (w/w) were determined as recently reported (Bak et al., 2013) and the same batch has been also used and analyzed in this study.
15.8% Ginseng (Korea), 15.8% Chaga (Russia), 10.6% Pinellia tuber (China), 5.2% Sparganium rhizome (China), 5.2% Alpinia rhizome (China), 5.2% Alpinia katsumadai seed (Vietnam), 5.2%
Astragalus Root (Korea), 5.2% Cinnamon bark (Vietnam), 10.6%
Dolichos seed, 10.6% Psoraleae semen (India), and 10.6% Arisaema rhizome (China). The herbal ingredients were obtained from the Oriental Medical Hospital, Dongguk University (Korea) and kindly authenticated by Dr. Seonghyun Jeong (Department of Oriental Herbal Materials, Dongguk University). Ethanol extracts from the above listed plants were prepared as follows. The dried and pulverized medicinal herbs were mixed together, and 1 kg batches were soaked in 40% ethanol (3 L). The ethanol extract was concentrated using a rotary evaporator, lyophilized, and then reconstituted in distilled water for the in vitro studies described below.
Cell culture. HPV 16-positive SiHa and CaSki cervical carcinoma cells and the HPV-negative cervical cancer cell line C33A were obtained from the American Type Culture Collection (USA). The cells were cultured in Dulbecco's Modified Eagle Medium (DMEM) medium (Hyclone Laboratories, USA) supplemented with 2 mM L-glutamine and 10% fetal bovine serum (Hyclone Laboratories USA), and incubated under humidified conditions at 37oC with 5% CO2.
Cytotoxicity assay. In order to test the effect of A1E on cell growth, cell viability was quantified using MTS. Cervical cancer cells (1×104) were seeded in 100µL of medium in 96-well plates and incubated overnight. After 20 h, the cells were treated with
various concentrations of A1E for 24 h. Media samples (100µL) were removed and washed one time. The cells were incubated with 20µL MTS (2 mg/mL) and PMS mixed solution in DMEM medium for 1–2 h at 37oC. The optical absorbance was measured at 492 nm using a spectrophotometer (Apollo LB 9110, Berthold Technologies GmbH, Germany).
Hoechst staining. Apoptotic nuclear morphology was observed using Hoechst staining and fluorescence microscopy (×100) (Latt et al., 1976). SiHa cervical cancer cells were seeded on coverslips in 6-well plates, and treated with various concentrations of A1E for 24 h. The coverslips were then washed twice with phosphate- buffered saline (PBS) and fixed with 4% paraformaldehyde for 1 h at room temperature. After washing 3 times with PBS, the cells were stained with the Hoechst staining solution at 37oC. The coverslips were then washed with PBS a further 3 times, dried completely, and mounted on microscope slides with mounting solution. The slides were observed using fluorescence microscopy.
Detection of apoptosis. Apoptotic cells were quantified by flow cytometry (Vermes et al., 1995). SiHa cells were plated in 6-well plates at a concentration of 2×105/well, incubated overnight to adhere, and treated with various concentrations of A1E for 48 h before harvesting and washing with PBS. Staining was performed with the FITC-Annexin V Apoptosis Detection Kit I (PharMingen, USA), in accordance with the manufacturer’s instructions (BD, USA). The percentage of late apoptotic cells was calculated from the Annexin-V-positive signal and the PI-positive signal.
Cell cycle analysis by flow cytometry. Cell cycle distribution was quantified using flow cytometry. The cervical cancer cells (2×105/well) were seeded in 6-well plates and incubated overnight to adhere. The cells were exposed to various concentrations of A1E for 24 h. The cells were then harvested, washed with PBS, and fixed with ice-cold 70% EtOH at −20oC. After fixation, the cells were washed with PBS and stained for 30 min with PBS containing 50µg/mL PI and 100 µg/mL RNaseA for 20 min in the dark. The DNA content was analyzed using a FACSCalibur instrument and CellQuest software (BD Bioscience). The sub-G1 cell population was counted, with their hypodiploid (2 N) fragmented DNA, indicating ongoing apoptosis.
Reverse-transcription polymerase chain reaction (RT-PCR) and real-time quantitative PCR. The cells were harvested and lysed using the easy-BLUE Total RNA Extraction Kit (iNtRon Biotechnology, Korea) according to the manufacturer’s instructions (Eckardt, 2010). RNAs (5µg) were reverse transcribed using M- MuLV reverse transcriptase (New England Biolabs, USA). RT- PCR analysis was performed using a PCR thermal cycler Dice instrument (TaKaRa, Japan) with the following primer sets: E6:
5'-ATG CAC CAA AAG AGA ACT GCA-3' (forward) 5'-TTA CAG CTG GGT TTC TCT ACG-3' (reverse); E7: 5'-ACA AGC AGA ACC GGA CAG AG-3' (forward), 5'-TGG GGC ACA CAA TTC CTA GT-3' (reverse); GAPDH: 5'-TGA TGA CAT CAA CAA GGT GGT-3' (forward), 5'-TCCTTG GAG GCC ATG TAG GCC-3' (reverse). Fas: 5'-AGG GAT TGG AAT TGA GGA AG-3' (forward), 5'-ATG GGC TTT GTC TGT GTA CT-3' (reverse) Fas-associated protein with death domain (FADD): 5'-
ACC TCT TCT CCA TGC TGC TG-3' (forward) 5'-CAC ACA GGT CTT CTT CCC CA-3' (reverse) TRAIL: 5'-GTC TCT CTG TGT GGC TGT AA-3' (forward) 5'-TGT TGC TTC TTC CTC TGG CT-3' (reverse). GAPDH was used as an internal control.
Real-time quantitative PCR was performed with a relative quantification protocol using the Chromo 4 Real-Time PCR system and iQ SYBR Green Supermix (both from Bio-Rad). All the target genes were normalized to the expression of the housekeeping gene GAPDH. Each sample was run with the following primer sets: E6 qPCR: 5'-TTG CTT TTC GGG ATT TAT GC-3' (forward), 5'-GGT TTG TTG TAT TGC TGT TC-3' (reverse); E7 qPCR: 5'-ATG CAT GGA GAT ACA CCT ACA TTG-3', 5v-TTA TGG TTT CTG AGA ACA GAT GGG-3';
GAPDH qPCR: 5'-GGC TGC TTT TAA CTC TGG TA-3' (forward), 5'-TGG AAG ATG GTG ATG GGA TT-3' (reverse);
FADD: 5'-ACC TCT TCT CCA TGC TG-3' (forward), 5'-CAC ACA GGT CTT CCC CA-3' (reverse). The fold changes in expression represent the ratio of E6 and E7 expression in the A1E- treated cervical cancer cells compared to the untreated control.
Western blot analysis. Cells were harvested using a scraper and collected by centrifugation (7 2×g, 5 min, 4oC). The cells were then washed with ice-cold PBS and centrifuged (1,890×g, 5 min, 4oC). The pellets were suspended in a lysis buffer containing 0.1%
sodium dodecyl sulfate (SDS), 0.1% sodium deoxycholate, 1%
Triton X-100, 1 mM EDTA, 0.5 mM ETDA, 140 mM NaCl, 10 mM Tris-HCl (pH 8.0), and a protease inhibitor cocktail. The cell lysates were incubated on ice for 2 h and then clarified by centrifugation at maximum speed for 30 min at 4oC. Equal quantities of protein samples (50µg) were separated by SDS- polyacrylamide gel electrophoresis (PAGE) and then transferred to polyvinylidene difluoride membranes. The membranes were blocked in 5% non-fat dried milk dissolved in Tris-buffered saline containing Tween-20 (2.7 M NaCl, 53.65 mM KCl, 1 M Tris- HCl, pH 7.4, and 0.1% Tween-20) for 1 h at room temperature before incubation with the specific primary antibodies for 2 h at room temperature. After the membraines were washed three times, the secondary antibodies (HRP-conjugated α-rabbit or α- mouse IgG) were incubated with the membranes for 1 h at room temperature. After washing 4 times, the signals were visualized using the Westzol plus Western Blot Detection System (iNtRON Biotechnology, Korea).
Nuclear and cytoplasmic fractionation. SiHa cells were treated with A1E for 24 h prior to harvesting and fractionation with NE- PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific Inc, USA) according to the manufacturer’s instructions.
Briefly, cells were collected by centrifugation at 1,980×g, 4oC, 5 min, washed with PBS and re-centrifuged. The cell pellet was suspended with buffer I, vortexed, and incubated on ice for 10 min. Buffer II were added and incubated for 1 min prior to centrifugation at 16,000×g, 4oC, 5 min, to produce a cytoplasmic extract supernatant. The insoluble pellet was then suspended with buffer III, incubated on ice for 40 min and centrifuged at 16,000×g for 10 min to produce a nuclear extract supernatant.
Equal quantities of protein from these extracts (50µg) were
separated by SDS-polyacrylamide gel electrophoresis.
Analysis of mitochondrial transmembrane potential (MMP).
The MMP (∆øm) was measured using JC-1 (5,5',6,6'-tetrachloro- 1,1',3,3'-tetraethyl-benzimidazolyl carbocyanine chloride) staining and flow cytometry. The cervical cancer cells were plated at a
seeding density of 3×105 cells/well, treated with various concentrations of A1E for 24 h, and harvested by trypsin-EDTA. Cells were then incubated with 5µg/mL of JC-1 for 10 min at 37oC in darkness.
Stained cells were washed with PBS and detected by FACS with a 488-nm laser.
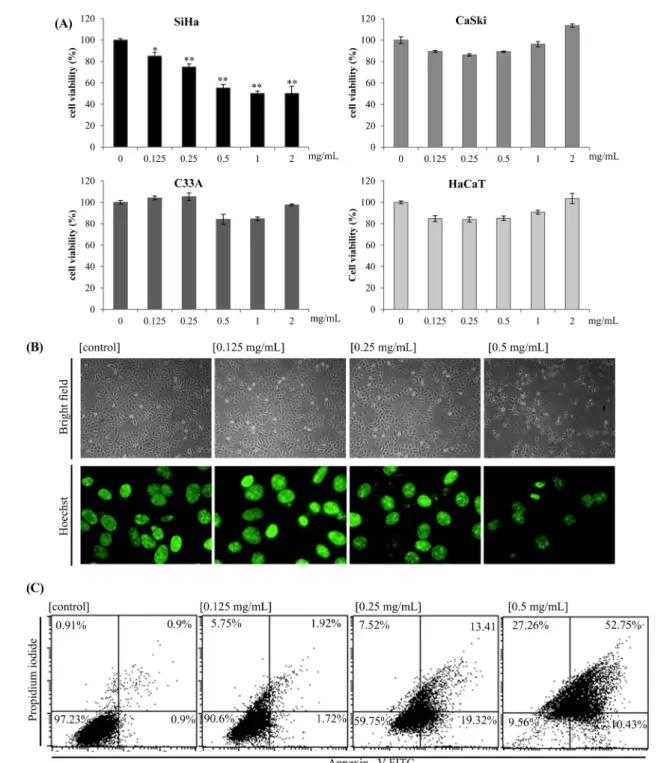
Fig. 1 Cytotoxic effect of A1E on cervical cancer cells. (A) SiHa, CaSki, C33A, and HaCaT cells were treated with A1E for 24 h, and the cell viabilities were measured by MTS assay. Data represent average percentages ± SD from 5 well experiments. *p <0.05, **p <0.01, and ***p <0.001, as compared to untreated control cells. (B) SiHa cells were exposed to the level of A1E indicated and imaged using phase-contrast microscopy (×40) and Hoechst nuclear staining using fluorescence microscopy (×100). (C) Cells were stained with AnnexinV-FITC/PI and analyzed by flow cytometry. The lower left quadrant (AnnxV/PI) represents viable cells, whereas the lower right quadrant (AnnxV+/PI) represents early-phase apoptotic and the upper right quadrant (AnnxV+/PI+) represents late-phase apoptotic/necrotic cells.
Statistical analysis. Data presented are the mean ± SEM of results from at least 3 independent experiments. Statistical significance was assessed with Student’s t-test. *p <0.05, **p <0.01 and ***p
<0.001 were considered statistically significant.
Results
A1E reduced proliferation and induced apoptosis in cervical cancer cells. The effects of A1E on the viability of several cervical cancer cell lines (SiHa, CaSki, and C33A) and HaCaT normal keratinocyte cell lines were determined by MTS assay (Fig. 1A). A1E exerted the cytotoxicity against SiHa cells in vitro with an estimated significant effect on cell viability at 0.125 mg/
mL and this reached a maximum of approx. 55.27% cell death in the presence of 0.5 mg/mL (Fig. 1A). However, CaSki, HPV 16- negative C33A, and HaCaT normal keratinocyte cells were only slightly affected by A1E. Therefore, we focused on SiHa cells.
Morphological changes were apparent under inverted phase- contrast microscopy. Treatment of SiHa cells with A1E for 24 h resulted in significant chromosomal condensation, detected by Hoechst staining (Fig. 1B). To determine whether the cytotoxic effect of A1E was associated with the induction of apoptosis, SiHa cells were treated with A1E, and then analyzed by flow
cytometry for the early apoptotic marker annexin V. The annexin V and PI staining results, in conjunction with the observed nuclear condensation, provided experimental evidence that A1E-induced cell death was caused by apoptosis (Fig. 1C).
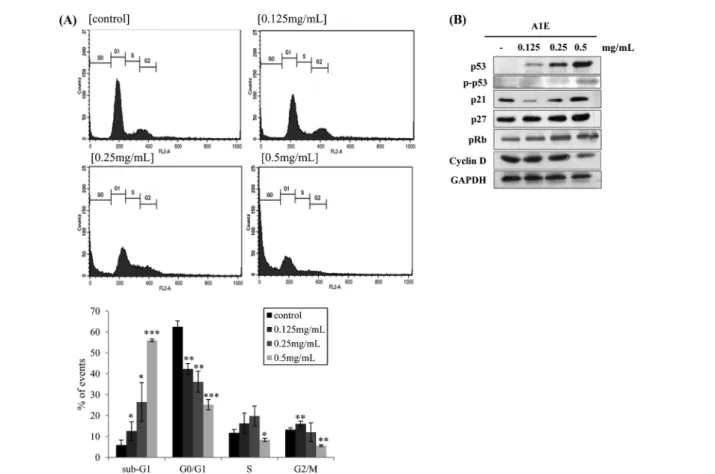
A1E induced sub-G1 phase and S phase arrest by modulating p53 and pRb and their targeting proteins involved in cell cycle progression. SiHa cells treated with A1E (0.5 mg/mL) for 24 h showed a significant alteration in cell cycle progression. A1E- treated SiHa cells showed an increase in the sub-G1 phase, indicating cleavage of nuclear DNA. The number of cells in the sub-G1 phase increased in a dose-dependent manner (Fig. 2A).
The S phase was also delayed up to the concentration of A1E (~0.25 mg/mL). However, almost cells were arrested by sub-G1 phase and the number of cells in the S phase decreased at high concentration of A1E (0.5 mg/mL) (Fig. 2A). The SiHa cervical cancer cell line has previously been shown to express E6 and E7 oncoproteins that degrade p53 and inactivate pRb, respectively (Furumoto and Irahara, 2002). The present study revealed that cell cycle modulating factors such as p53, p21, and p27 were increased in A1E treated SiHa cells (Fig. 2B). pRb protein levels also increased, while levels of cyclin D1 were down-regulated, supporting the finding that the cell cycle was arrested at the G1 phase and slightly arrested S phase, due to modulation of p53, pRb, and cyclin D1 expression (Fig. 2B).
Fig. 2 Effects of A1E on cell cycle and expression of p53 and Rb. (A) Effect of A1E on cell cycle. SiHa cells were stained with PI and analyzed by flow cytometry to measure cell cycle. Data represent average percentages ± SEM from 3 independent experiments. *p <0.05, **p <0.01, ***p <0.001 as compared to the control. (B) Effect of A1E on expression of p53, pp53, pRb, and cyclin D1 in SiHa cells.
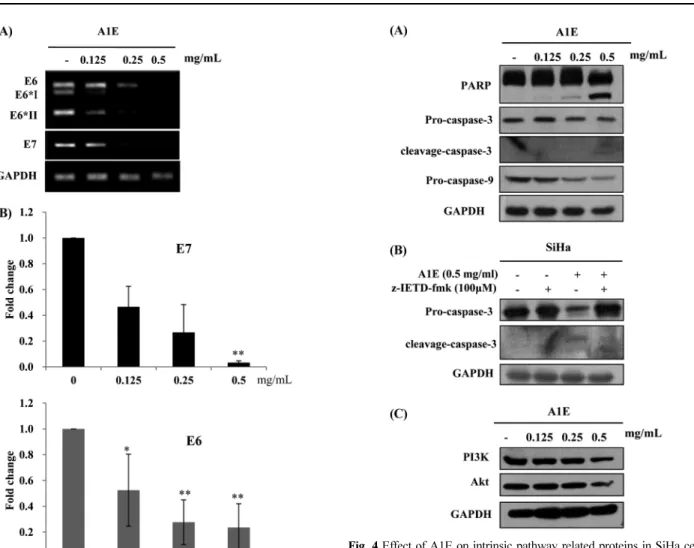
A1E inhibited HPV oncogene expression. Inhibition of HPV E6 and E7 gene expression in cervical cancer cell lines induces apoptosis. In order to assess whether A1E-induced cell death was associated with reduced expression of E6 and E7 oncogenes, their expression was analyzed in SiHa cells using PCR and qPCR.
These analyses revealed that A1E treatment resulted in reduced levels of both E6 and E7 as shown in Fig. 3. In particular, E6 full length as well as spliced forms (E6*I and E6*II) were inhibited by A1E.
The effect of A1E on apoptosis-related factors in SiHa cells.
Western blotting indicated that levels of pro-caspase-3 and -9 were decreased in A1E-treated SiHa cells, whilst cleaved PARP was detectable at high A1E doses (Fig. 4A). SiHa cells treated with
A1E in the presence of a caspase 9 inhibitor, z-IETD-fmk, showed suppressed activity of caspase-3, which is downstream of caspase- 9 (Fig. 4B). The PI3K signaling pathway, which plays pivotal roles in the signal transduction of cell survival, was also down- regulated in A1E-treated cells, where levels of both PI3K and Akt proteins were reduced (Fig. 4C).
The extrinsic pathway was not involved A1E-induced apoptosis.
Death receptor-related apoptosis occurs via the extrinsic pathway.
Death receptor-related gene expression of Fas, FADD, and TRAIL was analyzed in A1E-treated cells. Expression of Fas, FADD, and TRAIL were decreased in a dose-dependent manner (Fig. 5A). We performed real-time PCR to confirm these RT-PCR data, and these analyses also showed decreased Fas and FADD mRNA expression in A1E-treated cells (Fig. 5B). These results suggested that A1E-treated cervical cancer cells showed reduced susceptibility to death receptor-mediated apoptosis.
Role of mitochondria in A1E-induced cell death. A collapse in mitochondrial membrane potential is a critical step in the intrinsic apoptotic pathway. Using JC-1 staining, we detected A1E-induced Fig. 4 Effect of A1E on intrinsic pathway related proteins in SiHa cells.
(A) Representative western immunoblots of proteins of interest in the presence of increasing concentrations of A1E. (B) Representative western immunoblots of caspase-3 in the presence and absence of the caspase-9 inhibitor z-IETD-fmk. (C) The effect of A1E on PI3K and Akt protein levels in SiHa cells, determined by western immunoblotting.
Fig. 3 Effect of A1E on expression of E6/E7 mRNAs in SiHa cells. (A) Cells were treated with A1E for the indicated times and then harvested for mRNA extraction and PCR as described in Methods. Two micrograms of total mRNA, oligo dT, and reverse transcriptase were used for cDNA synthesis PCR using each primer. (B) E6 and E7 were both down-regulated by A1E in a dose-dependent manner, as compared to GAPDH. Three independent qPCR experiments were performed. The gene expression level obtained from real-expressed by comparing with the values non- treated SiHa cells. *p <0.05, **p <0.01 as compared to the control.
mitochondrial membrane potential breakdown. A1E-treated SiHa cells showed a clear shift in membrane potential, as shown in Fig.
6A. Disruption of the mitochondrial membrane potential is usually associated with the release of cytochrome c into the cytosol.
Fractionation of A1E-treated SiHa cells was used to detect released cytochrome c. As shown in Fig. 6B, cytosolic cytochrome c levels increased in the presence of A1E, but total levels in cell lysate were unaltered. Actin was used as a cell fractionation control (Fig.
6B). Fig. 6C shows expression of Bcl-2 and Bcl-xL, anti-apoptotic proteins showing reduced levels following A1E treatment.
Discussion
The main objective of this study is to investigate the anti-cancer effects of A1E in SiHa cervical cancer cells. A1E is formulated from extract of 11 plants with anti-cancer activities. We identified the A1E extract using GC-MS as recently reported (Bak et al., 2013). The A1E is consisted of coumarin and 2H-Furo[2,3-h]-1- benzopyran-2-one, and so on (Bak et al., 2013). Mainly, 2H- Furo[2,3-h]-1-benzopyran-2-one was known as angelicin and this compound has anti-cancer effects (Rahman et al., 2012). Coumarins are natural compounds found in many plants that possess medical value by itself and its modified derivatives (Sun et al., 2011).
HPV positive cervical cancer cells such as SiHa and CaSki, HPV-negative cervical cancer cell C33A, and HaCaT normal keratinocyte cell line were exposed to A1E. CaSki cells harbor several times more HPV genomes than SiHa cells (Snijders et al.,
1990). High-risk E6 and E7 oncogenes drive cell proliferation.
A1E significantly inhibited proliferation of SiHa cells, whereas it did not CaSki, C33A, and HaCaT cells. A1E targets E6 and E7 oncogenes and CaSki cells harboring more HPV genomes are more resistant to A1E compared to SiHa cells. Because nuclear condensation, DNA fragmentation, and exposure of phosphatidylserine at the cell surface are characteristics of the apoptotic cell death (Torchinsky et al. 1999; Vermes et al., 1995; Bak et al., 2011). We investigated whether A1E-induced cell death was apoptotic or necrotic. The results from Hoechst and Annexin-V/PI staining in our study indicated that A1E-treated SiHa cells were apoptotic (Fig. 1D).
Cell cycle analysis indicated an increased number of cells in sub-G1 and S phase arrest (Fig. 2A). p53 can induce G1 arrest through activation of transcription of the cyclin-dependent kinase inhibitors (CDKIs) such as p21 and p27 (Suzuki and Matsubara, 2011). p53, p21, and p27 were increased in A1E treated SiHa cells (Fig. 2B), reminded that these CDKIs can cause cell cycle arrest in cancer cells (Funk et al., 1997; Kim et al., 2000). Previous studies have shown that, when active Rb bound to E2F, transcriptional enhancers were inactivated and G1/S cell cycle progression was arrested (Ji and Zhu, 2005). Our results indicated that p53, p21, and p27 can works normally because A1E reduced HPV E6/E7 induced signaling. Tumor suppressor protein Rb was also enhanced by A1E. These effects could be due to the targeting E6 and E7 oncogenes by A1E.
SiHa cells express HPV 16 viral proteins E6 and E7, which are functionally targeting to p53 and pRb, respectively (Yamada et al., Fig. 5 Effects of A1E on expression of TRAIL, Fas, and FADD. (A) RT-PCR for Fas, FADD, and TRAIL mRNA exposed to increasing concentrations of A1E in SiHa cells. Levels of all 3 transcripts were decreased in a dose-dependent manner. (B) qPCR analysis of FADD and Fas mRNA expression levels. High concentration of A1E treatment significantly suppressed Fas mRNA level. The data presented 3 independent qPCR experiments.
**p <0.01 compared to the untreated control cells.
1995; Shaikh et al., 2012). We therefore analyzed E6 and E7 RNA levels in A1E-treated SiHa cells. As shown in Fig. 3, A1E treatment significantly decreased E6 and E7 expression levels in SiHa cells. Therefore, cell cycle related protein p53, p21, p27, and pRb works normally. Consequently, this process occur cell cycle arrest. As shown in the Fig. 2A, apoptosis (sub-G1 phase) was induced by A1E in a dose response manner and cell cycle arrest only occurred at low doses (~up to 0.25 mg/mL). Apoptosis progressed through sub-G1 level and thus cells were arrested in G1/S phase in SiHa cells treated with high concentration of A1E (Fig. 2A, 0.5 mg/mL).
There are 2 major apoptotic pathways: the intrinsic and extrinsic pathways. PCR analyses revealed that Fas, FADD, or TRAIL expressions were inhibited, suggesting that extrinsic pathway was not involved by A1E. These data may indicate a resistance to death receptor-mediated apoptosis in the SiHa cancer cell line, because A1E down-regulated the expression levels of Fas, FADD, and TRAIL (Fig. 5). In contrast, markers of the intrinsic apoptotic pathway, including PARP, caspase-9, and the downstream caspase- 3 pro-form were decreased in the presence of A1E and cleaved forms of PARP and caspase-3 were increased at high concentration of A1E. These results reminded that angelicin down-regulated the expression of anti-apoptotic proteins through intrinsic mitochondria mediated apoptotic pathway which does not participate in Fas/
FasL-induced caspase-8-mediated extrinsic pathway (Rahman et al., 2012). The caspase 9 inhibitor, z-IETD-fmk, suppressed A1E induced caspase-3 activity, which is downstream of caspase-9 (Fig. 4B). The caspase-3 inhibitor, z-DEVD-fmk, induced a
reduction in angelicin-induced cytotoxicity which confirmed that the intrinsic caspase-dependent pathway (Rahman MA et al., 2012). These results supported that A1E-induced cytotoxicity is mediated by the intrinsic caspase-dependent pathway during this apoptosis. Akt is one of the primary downstream targets of activated PI3K. We revealed that both PI3K/Akt and cyclin D expression levels were decreased by A1E. The PI3K/Akt pathway can induce cell cycle progression by modulating the protein stability of cyclin D. Akt phosphorylate pro-apoptotic Bad, and allows the excressive Bcl-2 or Bcl-XL to out-compete Bax (Chang et al., 2003). The reductions PI3K/Akt levels by A1E treatment caused the decrease of Bcl-2 and Bcl-xL, which are involved in mitochondria-mediated apoptosis proteins, the important components of the intrinsic pathway. We detected disruption of the mitochondrial membrane potential using JC-1, which aggregates into healthy mitochondria and fluoresces red.
Upon mitochondrial collapse in apoptotic cells, JC-1 no longer accumulates and is distributed throughout the cell, resulting in decreased red fluorescence (Koppikar et al., 2010). As shown in Fig. 6A, A1E-treated cells were shifted in this respect, and cytochrome c was released into the cytosol compared to control cells as expected. These data corroborated that A1E induced collapse of mitochondrial membrane potential.Bcl-2 and Bcl-xl expressions were also inhibited by A1E. Bcl-2 and Bcl-xL are frequently over-expressed in cancer cell lines and human cancer tissues (Chen et al., 2012).
In conclusion, A1E consists of several compounds which are targeting on several pathways such as E6/E7 pathway, mitochondrial Fig. 6 Effects of A1E on mitochondrial membrane potential and the intrinsic apoptotic pathway. (A) Mitochondrial membrane potential determined by JC-1 staining and flow cytometry. (B) Cytosolic cytochrome c levels, determined by western blotting, are shown in the presence of increasing concentrations of A1E. (C) Levels of Bcl-2 and Bcl-xl in the presence of increasing concentrations of A1E, showing decreased levels of these anti- apoptotic proteins in SiHa cells.
intrinsic pathway and PI3K/Akt pathway. A1E inhibited E6/E7 oncogenes and then induced intrinsic apoptosis via p53/pRb dependent pathways, and a mitochondria-mediated pathway. A1E also inhibited PI3K/AKT survival proteins. Therefore, our data suggested that A1E extracts were a promising resource as an agent of chemotherapeutic activity on SiHa cervical cells. However, it is necessary to elaborate further investigation of its bioactive compounds.
Acknowledgments This study was supported by a grant (B110053) from the Korean Health Technology R&D Project, Ministry of Health & Welfare, Republic of Korea. D.Y. was partially supported by a program (2012-0006686) from the National Research Foundation of Korea (NRF).
References
Bak Y, Ham S, Baatartsogt O, Jung SH, Choi KD, Han TY et al. (2013) A1E inhibits proliferation and induces apoptosis in NCI-H460 lung cancer cells via extrinsic and intrinsic pathways. Mol Biol Rep 40, 4507–19.
Bak YS, Kim HJ, Kang JW, Lee DH, Kim MS, Park YS et al. (2011) A synthetic naringenin derivative, 5-hydroxy-7,4'-diacetyloxyflavanone-N- phenyl hydrazone (N101-43), induces apoptosis through up-regulation of Fas/FasL expression and inhibition of PI3K/Akt signaling pathways in non-small-cell lung cancer cells. J Agric Food Chem 59, 10286–97.
Chang F, Lee JT, Navolanic PM, Steelman LS, Shelton JG, Blalock WL et al.
(2003) Involvement of PI3K/Akt pathway in cell cycle progression, apoptosis, and neoplastic transformation: a target for cancer chemotherapy.
Leukemia 17, 590–603.
Chen J, Zhou H, Aguilar A, Liu L, Bai L, McEachern D et al. (2012) Structure-Based Discovery of BM-957 as a Potent Small-Molecule Inhibitor of Bcl-2 and Bcl-xL Capable of Achieving Complete Tumor Regression. J Med Chem 55, 8502–14.
Doorbar J (2006) Molecular biology of human papillomavirus infection and cervical cancer. Clin Sci (Lond) 110, 525–41.
Eckardt NA (2010) Myo-inositol biosynthesis genes in Arabidopsis:
differential patterns of gene expression and role in cell death. Plant Cell 22, 537.
Franklin JL (2011) Redox regulation of the intrinsic pathway in neuronal apoptosis. Antioxid Redox Signal 14, 1437–48.
Funk JO, Waga S, Harry JB, Espling E, Stillman B, and Galloway DA (1997) Inhibition of CDK activity and PCNA-dependent DNA replication by p21 is blocked by interaction with the HPV-16 E7 oncoprotein. Genes Dev 11, 2090–100.
Furumoto H and Irahara M (2002) Human papilloma virus (HPV) and cervical cancer. J Med Invest 49, 124–33.
Ham SS, Kim SH, Moon SY, Chung MJ, Cui CB, Han EK et al. (2009) Antimutagenic effects of subfractions of Chaga mushroom (Inonotus obliquus) extract. Mutat Res 672, 55–9.
Ji P and Zhu L (2005) Using kinetic studies to uncover new Rb functions in inhibiting cell cycle progression. Cell Cycle 4, 373–5.
Kiefer D and Pantuso T (2003). Panax ginseng. Am Fam Physician 68, 1539–
42.
Kim TY, Choi KE, Cho HN, Ko HJ, Yang IW, Kim WJ et al. (2000) Expression of cyclin E and p27(KIP1) in cervical carcinoma. Cancer Lett 153, 41–50.
Kishida Y, Miki H, Nishii T, Inoue T, Nishida S, Yoshikawa H et al. (2007) Therapeutic effects of Saireito (TJ-114), a traditional Japanese herbal medicine, on postoperative edema and inflammation after total hip arthroplasty. Phytomedicine 14, 581–6.
Koppikar SJ, Choudhari AS, Suryavanshi SA, Kumari S, Chattopadhyay S, and Kaul-Ghanekar R (2010) Aqueous cinnamon extract (ACE-c) from the bark of Cinnamomum cassia causes apoptosis in human cervical
cancer cell line (SiHa) through loss of mitochondrial membrane potential. BMC Cancer 10, 210.
Latt SA and Stetten G (1976) Spectral studies on 33258 Hoechst and related bisbenzimidazole dyes useful for fluorescent detection of deoxyribonucleic acid synthesis. J Histochem Cytochem 24, 24–33.
Lee JE, Seo IW, Jeong SJ, Koh W, Jung JH, Kwon TR et al. (2011) Herbal cocktail ka-mi-kae-kyuk-tang stimulates mouse bone marrow stem cell hematopoiesis and janus-activated kinase 2/signal transducer and activator of transcription 5 pathway. Am J Chin Med 39, 1235–52.
Lee MY, Shin IS, Jeon WY, Seo CS, Ha H, Huh JI et al. (2012) Protective effect of Bojungikki-tang, a traditional herbal formula, against alcohol- induced gastric injury in rats. J Ethnopharmacol 142, 346–53.
Lucken-Ardjomande S and Martinou JC (2005) Newcomers in the process of mitochondrial permeabilization. J Cell Sci 118, 473–83.
Muise-Helmericks RC, Grimes HL, Bellacosa A, Malstrom SE, Tsichlis PN, and Rosen N (1998) Cyclin D expression is controlled post-transcriptionally via a phosphatidylinositol 3-kinase/Akt-dependent pathway. J Biol Chem 273, 29864–72.
Munagala R, Kausar H, Munjal C, and Gupta RC (2011) Withaferin A induces p53-dependent apoptosis by repression of HPV oncogenes and upregulation of tumor suppressor proteins in human cervical cancer cells.
Carcinogenesis 32, 1697–705.
Nakajima Y, Nishida H, Matsugo S, and Konishi T (2009) Cancer cell cytotoxicity of extracts and small phenolic compounds from Chaga [Inonotus obliquus (persoon) Pilat]. J Med Food 12, 501–7.
Rahman MA, Kim NH, Yang H, and Huh SO (2012) Angelicin induces apoptosis through intrinsic caspase-dependent pathway in human SH- SY5Y neuroblastoma cells. Mol Cell Biochem 369, 95–104.
Shaikh F, Sanehi P, and Rawal R (2012) Molecular screening of compounds to the predicted Protein-Protein Interaction site of Rb1-E7 with p53- E6 in HPV. Bioinformation 8, 607–12.
Sima N, Wang S, Wang W, Kong D, Xu Q, Tian X et al. (2007) Antisense targeting human papillomavirus type 16 E6 and E7 genes contributes to apoptosis and senescence in SiHa cervical carcinoma cells. Gynecol Oncol 106, 299–304.
Snijders PJ, van den Brule AJ, Schrijnemakers HF, Snow G, Meijer CJ, and Walboomers JM (1990) The use of general primers in the polymerase chain reaction permits the detection of a broad spectrum of human papillomavirus genotypes. J Gen Virol 71, 173–81.
Sun JG, Chen CY, Luo KW, Yeung CL, Tsang TY, Huang ZZ et al. (2011) 3,5-Dimethyl-H-furo[3,2-g]chromen-7-one as a potential anticancer drug by inducing p53-dependent apoptosis in human hepatoma HepG2 cells.
Chemotherapy 57, 162–72.
Suzuki K and Matsubara H (2011) Recent advances in p53 research and cancer treatment. J Biomed Biotechnol 2011, 978312.
Tan S, de Vries EG, van der Zee AG, and de Jong S (2012) Anticancer drugs aimed at E6 and E7 activity in HPV-positive cervical cancer. Curr Cancer Drug Targets 12, 170–84.
Torchinsky A, Ivnitsky I, Savion S, Shepshelovich J, Gorivodsky M, Fein A et al. (1999) Cellular events and the pattern of p53 protein expression following cyclophosphamide-initiated cell death in various organs of developing embryo. Teratog Carcinog Mutagen 19, 353–67.
Vermes I, Haanen C, Steffens-Nakken H, and Reutelingsperger C (1995) A novel assay for apoptosis. Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin V.
J Immunol Methods 184, 39–51.
Walker NI, Harmon BV, Gobe GC, and Kerr JF (1988) Patterns of cell death.
Methods Achiev Exp Pathol 13, 18–54.
Yamada T, Yamashita T, Nishikawa T, Fujimoto S, and Fujinaga K (1995) Biologic activity of human papillomavirus type 16 E6/E7 cDNA clones isolated from SiHa cervical carcinoma cell line. Virus Genes 10, 15–25.
Yim NH, Jung YP, Kim A, Ma CJ, Cho WK, and Ma JY (2013) Oyaksungisan, a Traditional Herbal Formula, Inhibits Cell Proliferation by Induction of Autophagy via JNK Activation in Human Colon Cancer Cells. Evid Based Complement Alternat Med 2013, 231874.