Kor. J. Appl. Microbiol. Bieng Vol. 11, No. 4, 297-301 (1983)
297
Trypsín Inhibitor from Streptomyces Sp. (III)
- Cultural conditions for the inhibitor production-
Dong heu ì Yì, and Jung hwn Seu *
Department of Microbiological Technology Kon-Kuk University Seoul, Korea Department of Microbiology, Kyung-Pook National University Taegu, Korea
(Received December 1, 1983)
Streptomyces
屆園**가 生成 하는
Trypsin Inhibitor (III)-펌害物質의 生屋條件-
이동희 · 서정훈*
건국대학교 미생물공학과
*경북대 학교 마생물학과 (1983 년 12 월 1 일 수리 )
Streptomyces sp. AS-707 was aerobically cultured with reciprocal shaker (5 cm, 90 strokeslmin) at 350C and initial pH 7. The production 01 the trypsin inhibitor reached at maximum after 3 days in the medium containing 2.0%
glucose, 0.3 % peptone, 0.05 % sodium nitrate, 0.05 % K2 HP04, 0.05 % MgS04. 7바 o and 0.05% NaCL Peptone, yeast extract, casein, aspartic acid, lysine and sodium nitrate as nitrogen sources were favorable, but all the sugars were
얻qually utilized as carbon sources. The production of the inhibitor was depressed by silver, cupric, lead and mercuric salts.
lnhibitors against proieolytic enzymes have been found to be produced in animals and plants{l) and also in micro- organisms(2, 3). And most of microbial protease inhibitors have been reported to be produced by actinomycetes(4, 5) and fungi(6)‘ The inhibitors are useωin the diagnosis and therapy of inflammation, pancreatitis and tumor caused by proteases(7, 8), and are capable of the potential application in biochemistry and physiochemistry(9, 1 이 Moreover, certain inhibitors can bind onto the cellular suriaces and modify the lunctions 01 immune response-participating cells{lI.13)‘
A trypsin inhibitor produced by Streptomyces sp. AS-707 was obtained and the purification, stabilities and biological activities of the inhibitor were reported in the previous papers(14, 15). In this present paper, c비tural conditions lor production of the trypsin inhibitor are described.
Materials and Methods
Microorganism
The microorganism used in this work was Streptomyces
sp. AS-707, which was isolated from s이I and was maintained on J)otato dextrose agar (pH 7) containing sodium nitrate.
Cultivation methods
A 100 mL-Erlenmeyer flask containing 20 mL 01 basal medium was inoculated with 0.1 mL 01 spore suspension 01 Streplomyces sp. AS-707 and cultivated at 350C lor 3 days on a reciprocal shaker (5 cm, 90 strokes/min). The basal medium (pH 7) was composed 012.0% glucose, 0.3% pep- tone, 0.05% MgS040 7H20, 0.05% K2HP04 and 0.05% NaCL The effect 01 carbon and nitrogen sources and metal salts were studied by substituting various carbon compounds lor glucose, nitrogen compounds for peptone and metal salts lor MgS04.7H20 in the basal medium. The role of amino acids in the inhibitor production was determined by adding these in the concentration of 0.05 % into the medium containing 2.0% glucose, 0.2% NaN03, 0.05% CaCI202H20, 0.05%
K2HP04 and 0.05% NaCL The effect olvitamins was studied by adding these in the concentration 01 2.5 μglmL into the basa! medium.
298
Determination of the inhibitor activity
Determination 01 inhibitory activity was carried out by measuring residual tryptic activity after solution 01 trypsin was preincubated with cultured liltrate by the method described previously(l4l
Estimation of cell growth
Cultured microorganisms were gathered by filtration and then washed with distilled water 3 times. Thereafter they were dried at 100-11OoC and weighed.
Results
Kor. J. Appl. MiC1'obiol. Bieng.
·---(
100
*lL
%
。
¢ ·ζ닙애 ”‘~r;i써{@며
20 ..J E
Z .-<
、、、
bIJ 를 } .
‘ t t
10 ξ
"t:i
5
Concentration of glucose (%)
l @
Q
Sources of carbon
The effect 01 different carbon sources on the trypsin in- hibitor production was studied. From the results shown in Table 1, it was lound that among the carbon sources used ex-
cept salicin none was specilic as carbon sources. So we chose tion 01 the inhibitor. The conceniration 01 glucose in the glucose and examined the optimal concentration lor produc- medium atfected either cell growth or the inhibitor produc-
Table 1. Effect of carbon sources on the inhibitor production by Streptomyces AS-707
carbon source growth final pH relative activity
D-glucose ++++ 5.6 100
D-galactose +++ 6.2 95
다 mannose ++ 6.0 92
다Iructose ++++ 5.4 103
D-sorbose ++ 6.0 98
다xylose +++ 6.2 100
D-arabinose ++ 6.0 82
D-erythrose ++ 6.0 97
D-rhamnose ++ 6.0 94
D-sorbitol +++ 6.0 99
D-arabitol +++ 6.0 100
mannitol ++ 6.0 100
Î-inositol ++ 6.0 82
xylitol ++ 5.8 100
glycerol +++ 5.4 97
sucrose +++ 6.0 100
lactose +++ 6.4 99
maltose ++ 6.4 94
raffinose ++ 6.0 85
inulin ++++ 6.0 96
dextrin +++ 5.6 102
starch ++++ 5.4 94
salicin + 6.0 64
Fig. 1. Effect of concentration of glucose on the trypsin inhibitor production by Streptomy- ces AS-707.
tion (Fig. 1). The optimal concentration of glucose lor produc- tion 01 the inhibitor was abnut 2%, but lor cell growth 3%
glucose was suitable.
Sources of nitrogen
The effect 01 nitrogen sources was tested by adding several kinds 01 organic and inorganic nitrogen sources. As shown in Table 2, peptone, yeast extract and sodium nitrate were favorable sources for the inhibitor production‘ The op- timal concentration 01 peptone was 0.3%, but the best cell growth was occurred in 0.2 % (Fig. 2)‘ And addition 01 sodium nitrate to the medium increased the inhibitor pìOduc- Table 2_ E한'ect of nitrogen sources on the trypsin in.
~ibitor production by Streptomyces. AS.707
nitrogen source growth linal pH relative activity
peptone ++++ 5.4 100
yeast extract ++++ 5.8 93
casem ++++ 5.4 86
malt extract ++++ 5.6 69
beel extract ++++ 5.6 63
corn steep liquor +++ 5.4 58
asparagme ++++ 5.8 43
(NH2hCO + 5.4 3'7
(NH4hHP04 +++ 5.0 14
(NH4hS04 + 5.。 0
NH4CI ++ 5.0 0
NaN03 ++ 5.6 93
NaN02 ++ 5.4 22
m
m비
mm
m
mm
m m m
mm
%
m
때
m
%
mm
m
mm
빼
m영mm
%
때 뻐
m
mm mm
m
299
....l.
o E 20:-< •
나)
E
*‘ 승
10 음
..c ...
능
a 。
25 30 35 40
Cultured temperature ("C) ð
Fig.4. Effect of temperature on the production of the trypsin inhibitor by Streptomyces AS .. 707.
none
Table 3. Effect of amino acids on the inhibitor pro- duction by Streptomyc양 AS-707
relative actIvlty
++++
growth
+++
+++
+++
+++
+++
+++
peptone glycine alanine valine leucine isoleucine senne threonine cysteme cystine methionine phenylalanine tyrosme tryptophan proline hydroxyproline asparti<; acid glutamic acid asparagme glutamine Iysine
++
++
++ ++ ++
~ / ..
/ / / / / i
/ / / /
i
100
。
O
〉、
흥 :50 +‘ 딩
ol
'"
... >애 잉
여
amino acid 0
+++
+++
o. 1 0.3 0.5
Concentration of peptone (%)
Fig.2. Effect of peptone as a nitrogen source on the trypsin inhibitor production.
tion and cell growth up to about 10% (Fig. 3) Effect 01 temperature and pH
The temperature and pH requirements lor production 01 the inhibitor in the basal medium are illustrated in Fig. 4 and 5, respectively. As shown in Fig. 4, the maximal inhibitor ac- tivity was attained at 350C, and also cell growth was best at the temperature‘ Fig. 5 shows the ellect 01 the initial pH 01 the medium on growth and production 01 the inhibitor, and the initial pH 7 was most lavorable for growth and the in- hibitor' production
Effect of amino acid and vitamin
Among the amino acid used, aspartic acid and Iysine played an important role on the inhibitor production and in- creased yield up to 30-40%, but the level didn’t exceed that of peptone addition (Table 3). And all vitamins used did not affect on the production 01 the inhibitor and cell growth (Table 4)
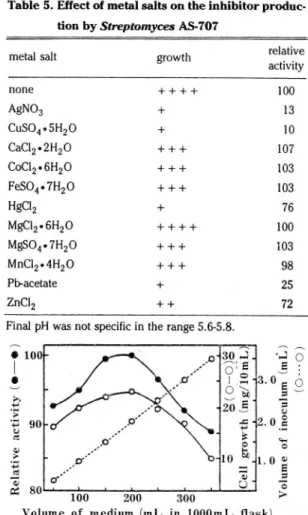
Effect of metal salts
In the production 01 the inhibitor, AgN0 3, CuS04' HgCI2,
--
20::::-
」
E o
、、~‘
E bl)
10 ‘‘ 승
;., i
"0
s ...
*
@
&
@
Q Vol. 11. No.4
100
Õ 90
O
γ 80
>‘
+‘
>
*‘ 엉 70
'"
... >
!:! 60
。」
0::
+++
++++
+++
+++
+++
+++
+++
+++
+++
+++
Final pH was not specific in the range 5.4-5.8 +++
+++
+++
arglnme histidine ornithine
β-alanine
GABA
0.1 0.3 0.5
Concentration of NaN03 (%) Fig.3. Effect of sodium nitrate as a accessory
nitrogen source on the inhibitor production.
(」E-【\벼E
ia
~〔』}〕)-ig、rFPF야
-- -}]
정 (---·)때 πω
%
%
% I (。|1O){“;;〕얘 @〉-§i@여
0
Kor. J. Appl. Microbiol. Bieng
Table 5, Effect 01 metal salts on the inhibitor produc.
tion by Streptomyces AS.707
relative activity growth
+
nu qu nU
?’
qJ
”J
”o
nU qa ng
q니 이ι
m
l I
m m m
7
m m
9 2 T ++++
+++
+++
+++
+ +
++++
+++
+++
+ metal salt
none AgNû3 CuSû405H2ü CaC12.2H2ü CoC12.6H2ü FeSü4. 7H2Û HgCI2 MgC12.6H2ü MgSÜ4.7H2ü MnC12.4H2ü Pb-acetate ZnCI2 :1iEξ융E ·gν* ‘{』딩) 공〉〉g빼
:@Q (---·)
때 애
300
~ 100 O
O
>‘
...
윌 80
‘J
'"
Q)
>
*‘ 어
@
Q;: 60
5 7 ' 9
Initial pH
Fig. 5, Effect of the initial pH on the inhjbitor production by Streptomyces AS-707.
3
Table 4. E야'ect of vitamins on the inhibitor produc.
tion by Streptomyces AS.707
++
,,;30 니 J '..J n
./.-1'0: 트 I ~튿 i
cf , l ;::; 13. 0 c 。
’ ., , 시
/、 I V bIi I =
&\... ~2ó흥 1 응
g 、잉 \ l £ 12. o §
.• 0" \-1
허 강
。.' 'OilO ... ~L 0 연
σ 펴 j 흐
! ] 1 Õ
80' 100 200 300
Volume of medium (mL in 1000mL flask) Fig.6. Effect of aeration on the inhibitor product
ion by Streptomyces AS-707.
Final pH was not specilic in the range 5.6-5.8.
뻐
%
(·--·)
kii、r;υi써 !}〉ii어i@여
relative activity 100 106 lOl 106 lO6 99 103 1lO 100 97 104 final pH
5.2 5.8 6.0 5.4 5.4 5.0 5.8 6.0 5.6 5.6 none
ascorbic acid inositol pyridoxine Ca-pantothenate lolic acid nicotinic acid PABA biotin B12 thiamine vitamin
, ~
20 같 φ o 's:.ì, 0
;; 115 E ~
701
I I 성
6~~ 1O:
::t I 'tl ..5 0.1 도 15 -.:;
* @
&
3 5 7 9 11 공
Cultured timc (day) υ
Fig.7. Time course of the inhibitor pooduction 8
5.0
.. 100
-
-、+‘
.~ ... ..
잉 여
@
... >
'"
Q;: 。“
The amount 01 cell mass was similar in all cases‘
PbAc2 and ZnCI 2 repressed severely cell growth and the in- hibitor production, but CaCI 2 showed slight increasing effect on the production 01 the inhibitor (Table 5)
Effect of aeration
As illustrated in Fig. 6, the flask containing 200 mL 01 the medium gave the highest production 01 the inhibitor and cell
by Streptomyces AS-707.
The production 01 trypsin inhibitor Irom Streptomyces sp.
AS-707 appeared very dependent on the conditions Iike availability 01 nutrients, pH, temperature and aeration rate.
Discussion mass
Cultivation course
The production 01 the inhibitor, cell growth and variation 01 pH with the elapse 01 cultivation time were tested. Cultiva- tion was carried out at 350C with 200 mL 01 inhibitor produc- tion medium in 1000 mL-Erlenmeyer f1ask on the shaker Fig. 7 shows the time course 01 cultivation. lnhibitory activity reached. maximum after 3 days cultivation and it was very unstable in the culture.
Vol.ll. No. 4
Among the tested inorganic nitrogen sources, sodium nitrate gave the highest yield 01 the trypsin inhibitor, whereas ammonium saIts gave the lowest yield. On the other hand, the celis grew and produced the inhibitor weli on ali organic nitrogen sources used. These tendencies were similar to the production 01 the trypsin inhibitor reported by Tsuchiya and Kimura(16), but iI1, the availability 01 sodium nitrate it was different. And in the. other cases lor the in- hibitor production with other Streptomyces(17-19), organic nitrogen sources were added at the concentration 01 2-5 % , but Streptomyces sp. AS-707 produced maximallevel 01 the inhibiior with 0.3% peptone only
In the production 01 various enzyme inhibitors, metal salts have been known to play important role(20), but in this study none 01 the used meta1 salts except CaCl2 stimulated the inhibitor production, moreover AgN0 3, CuS04 and PbAc2 repressed the inhibitor production severely.
The temperature optimum lor maximal yield by Strep- tomyces AS-707 was 350C, however other protease inhibitors lrom other species 01 Streptomyces were 25-290C (Kondo et al.(21), Umezawa et al.(22), Murao and Watanabe(20)), 18-200C (Jönsson and Torstensson(23)). But in the case 01 pH op- timum, it was similar to many other production 01 Strep- tomyces protease inhibitoril8, 19.24).
[n the cuitivation course, production 01 the inhibitor by the organism was reached to maximum in the early stage This was a general trend in the production 01 Streptomyces protease inhibitori3, 17.19,25.27)
要 約
301
B 本 東京 (1978) .
4. Aoyagi, T., H. Umezawa: Proteases and Biological Con- trol, Cold Spring Harbor Lab., p. 429 (1975)
5. Umezawa, H.: Methods Enzymol., 45, 678 (1976) 6. Shimada, K. and K. Matsushima: 1. Agric. Chem .. Soc.
Japan, 42, 325 (1968)
7. Vogel, R., 1. Trautshould and E. Werle: Natural Protease Inhitibor, AP, New York, p. 111 (1968)
8. Yamamoto, R.S., H. Umezawa, T. Takeuchi, T .. Mat- sushima, K. Hara and T. Sugimura: Proc. Amer. ,4ssn.
Cancer Res., 15,38 (1974)
9. Kinsella, A.R. and M. Radman: Proc. Natl. Acad. Sci USA, 77, 3544 (1980)
10 냐sch, J., R. Koelsch, G. Brezesinki, V. Herrmann, S.
Riemann, and P. Bohley: Acta. Biol. Med. Ger., 36, 1829 (1977)
11. Umezawâ, H., M. [shizuka, T. Aoyagi and T. Tàkeuchi:
1. Antibiot., 29, 857 (1976)
12. Umezawa. H.: 1. Antibiot., 30, 138 (1977)
13. Aoyagi, T., M. [shizuka, T. Takeuchi and H. Umezawa:
1. Antibiot., 30, 121 (1977)
14. Yi, D.H. and J.H. Seu: Kor. 1. Appl. Microbiol. Bioeng., 10, 275 (1982)
15. Yi, D.H. and J.H. Seu: Kor. 1. Appl. Microbiol. Bioeng., 10, 283 (1982)
16. Tsuchiya, K. and T. Kimura: Appl. Enuiron μicrobiol., 35,631 (1978)
17. Wingender, W., H. Hugo and W. Fommer: 1. Antibiot., 28,611 (1975)
18. Murao, S., and T. Watanabe: Agnc. Biol. Chem., 41, 1313 (1977)
Streptomyces 屆j 園林 AS-707 로 부터 trypsin 19. Aoyagi, T.' H. Tobe, F. Kojima, M. Hamada, T. Takeuchi inhibitor 의 生품條件을 검토한 결과 질소원으로서는 and H. Umezawa: 1. Antibiol., 31, 636 (1978) peptone 등의 대 다수의 유기 질소원과 NaNO, 가 좋 20. Murao, S. and T. Watanabe: Agric. Biol. Chem., 42,2209 은 結果플 보였으며 탄소원우로서 는 glucose를 비롯 (1978)
하여 대부분이 유사한 결과를 보였다 生훌最適 pH 21‘ Kondo, S., K. Kawamura, J. [wanaga, M. Hamada, T 와 온도는 각각 7 과 350C 였으며 , glucose 2 %, p- Aoyagi, K. Maeda, T. Takeuchi and H. Umezawa: Chem eptone 0.3%, NaNO, 0.05% 를 加한 培地에서 3 Pharm. Bull., 17, 1896 (1969)
일간 진탕배양하였을 띠1 ì:且害物質生成量이 최대에 22. Umezawa, H., T. Aoyagi, H. Morishima, M. Matsuzaki,
達하였다 M. Hamada and T. Takeuchi: 1. Antibiot., 23,259 (1970)
Literatures cited
1. Kassell, B.: Methods Enzymol., 19, 839 {197이 2. Umezawa, H.: Enzyme [nhibitors 01 Microbial Origin,
University 01 Tokyo press, 1972
3 졸柳;흑 ßI'! : 짧뚫 ;8 술物함 n.! ; - " " .:1土-、γ 소쫓
24. Andreeva, N.A. and D.N. Chermenskii: Biokhimiya, 44,
838 (1979)
25. Murao, S. and S. Sa!o: Agric. Biol. Chem., 36, 160 (1972) 26. Murao, S., S. Sato and N. Muto: Agric. Biol. Chem., 36,
1737 (1972)
27. Murao, S., K. Oda and T. Koyama: Agric. Biol. Chem., 42, 899 (1978)