3T3-L1세포에서 AMPK 활성이 TNF-α에 의한 지방세포 대사 이상 및 Wnt와 sFRP5에 미치는 영향
The Effect of AMPK Activation on Wnt and sFRP5 in TNF-α Induced Adipocyte Metabolic Dysfunction in 3T3-L1 Cell
홍석우․이진미․최지훈
(1)․박세은
(1)․이은정
(1)․박철영
(1)․오기원
(1)․박성우
(1)․김선우
(1)․이원영
(1)*강북삼성병원 의학연구소, 성균관대학교 의과대학 강북삼성병원 내분비내과(1)
Seok-Woo Hong, Jin-Mi Lee, Ji-Hun Choi(1), Se-Eun Park(1), Eun-Jung Rhee(1), Cheol-Young Park(1), Ki-Won Oh(1), Sung-Woo Park(1), Sun-Woo Kim(1), Won-Young Lee(1)*
Institute of Medical Research; and Department of Endocrinology and Metabolism
(1), Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine
요 약
연구목적: 지방세포에서 만성적인 염증은 비만 및 대사질환 과 매우 밀접하게 관련되어 있다. 본 연구는 지방세포의 염증 유발 및 회복 과정에서 Wnt 신호전달체계 및 지방대사의 변화 를 관찰하였다.
방법: 3T3-L1 지방 세포주에 Tumor-necrosis factor (TNF)-α를 단독, 혹은 Aminoimidazole carboxamide (AICAR)와 함께 처리하여 염증의 유발 및 회복을 유도한 지 방세포 모델을 제작하였다. Wnt 신호전달체계 및 지방대사에 관련된 인자의 변화를 관찰하였다.
결과: 지방세포에 TNF-α의 처리는 염증 인자의 발현 증가 및 non-canonical Wnt 신호전달체계의 활성화를 유도하면서, PPAR-γ의 발현 감소와 지방분해가 증가를 유도하였다.
AICAR의 처리는 TNF-α에 의한 염증반응을 억제하고, non-canonical Wnt 신호전달체계 및 지방대사의 변화를 회복 시켰다. 또한 항염증 아디포카인인 sFRP5의 분비가 유의하게 증가하였다.
결론: 본 연구는 염증이 유발된 지방세포에서 AMPK 활성 화가 sFRP5 분비를 촉진하여 non-canonical Wnt 신호전달체 계를 억제하고, 염증억제 및 지방대사를 회복할 수 있음을 제 시한다.
ABSTRACT
Background: Chronic inflammation in adipose tissue is
closely associated with metabolic diseases, such as, type 2 diabetes and obesity. In the present study, we investigated the Wnt signaling pathway and fat metabolism in the course of TNF-α-induced inflammation and recovery in 3T3-L1 adipocyte.Methods: We stimulated the fully differentiated 3T3-L1
adipocyte with either TNF-α only or with both TNF-α and AICAR for 24 hrs to induce inflammation. We assessed the alteration of Wnt signaling pathway and the factors associated with fat metabolism with western blot assay and real-time PCR.Results: The expression of inflammatory cytokines, IL-6
and MCP-1, was increased by TNF-α treatment in fully differentiated 3T3-L1 adipocyte, and the marked activation of Wnt5a, a noncanonical Wnt ligand, was observed. The expression of PPAR-γ was reduced, and the lipolysis measured by glycerol release, was markedly increased. The activation of AMPK by AICAR inhibited the TNF-α-induced inflammation, reversed the alteration in Wnt signaling pathway, and reversed fat metabolism induced by TNF-α.AMPK activation stimulated the secretion of sFRP5, an anti-inflammatory and anti-Wnt signaling adipokine.
Conclusion: The activation of AMPK suppressed
noncanonical Wnt signaling pathway and protected the adipoctye from inflammation and lipolysis, induced by TNF- α treatment, by sFRP5 stimulation. Based on these results, we suggest that noncanonical Wnt signaling pathway and sFRP may have important roles in metabolic diseases such as, obesity and diabetes.ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
접수일자: 2011년 11월 15일, 심사일자: 2012년 1월 16일, 게재승인일자: 2012년 2월 22일 교신저자: 이원영, (110-746) 서울시 종로구 평동 108 성균관대학교 의과대학 강북삼성병원 내분비내과 Tel: 02-2001-2579, Fax: 02-2001-1588, E-mail: [email protected]
* 본 논문은 대한비만학회 2010년 문석연구비 지원에 의하여 작성되었습니다.
서 론
비만은 지방의 축적과 에너지 저장을 위한 조절과정에 주로 발생하는데1), 지방세포 내에 과도한 지방의 축적은 세 포 내 염증반응과 동반하여 당뇨병, 고혈압, 심혈관 질환을 포함한 만성질병을 유발하게 되는 원인으로 알려져 있다.2,3) 따라서 세포 내 과도한 지방의 축적이나 염증반응을 예방하 거나 축적된 지방을 분해하도록 자극하는 방안에 대한 연구 가 최근 많은 관심을 끌고 있다. 지방세포 내의 지방방울 (lipid droplet)은 지질의 대사 및 조절에 중요한 역할을 하 는 것으로 알려져 있으며 지방방울 내에 주로 존재하고 있 는 중성지방의 분해와 이로 인한 glycerol의 유출은 세포 내 지방의 축적을 조절하는 중요한 기전으로 여겨져 왔다.4) 특 히 lipase, Tumor-necrosis factor (TNF)-α, perilipin A 등은 중성지방 분해 과정의 조절에 중요한 역할을 담당하고 있 다.5-7)
Wnts는 세포로부터 분비되는 cystein이 많은 당 단백질 로서, 주변세포의 수용체(receptor)에 결합하여 활성화된 수 용체를 통해 여러 가지 단계를 거쳐 많은 유전자의 발현을 조절함으로써, 다양한 생명 현상을 조절한다고 알려져 있
다.8-10) 최근에 Wnt 신호전달체계가 지방세포 형성의 억제
효과가 발표되어, 지방세포 형성의 조절 target으로서 Wnt signaling에 대한 연구가 활발히 이루어지고 있다.11)
지방세포 분화(adipocyte differentiation) 과정에서는 Wnt 신호전달체계 중, canonical 신호전달체계의 기능이 매우 강 조되어 왔다. 지방전구세포에서 발현되는 Wnt10b는 세포 내 조절인자로서 지방세포 분화에 중요 전사인자인 CCAAT enhancer binding protein (C/EBP)α와 peroxisome proliferation activator receptor (PPAR)γ의 발현을 억제함 으로써 지방세포 특이 유전자가 발현되지 못해 지방세포 형성을 막는다.12) 또한 Wnt10b가 증가시키는 세포 내 β-catenin양과 PPARγ 간의 균형 조절이 지방 세포 분화에 중요하다고 알려져 있으며 이 두 단백질 간의 균형은 Glycogen synthesis kinase (GSK)3β에 의한 β-catenin의 인 산화를 통한 단백질 분해에 의존적으로 조절되고 있음이 확 인되었다.13) 지방조직을 비롯한 다양한 조직에서 유전학적, 생화학적인 방법을 통하여 많은 연구가 canonical Wnt 신 호전달체계의 규명에 집중이 되어 왔지만, 최근에는
“noncanonical Wnt 신호전달체계”의 생물학적 중요성이 인 식되면서 이 분야의 연구가 점차 활성화되는 추세이다.
AMP-activated protein kinase (AMPK)는 지방산의 합성 과 분해를 매개함으로써 간 지방 대사의 조절에 중요한 역 할을 담당한다. AMPK의 활성화는 ATP를 소비하는 과정 에 필요한 효소들을 인산화 및 불활성화시키고, 지방산 산 화를 증가시킨다.14) AICAR 또는 metformin의 처리를 통한 AMPK의 활성화는, 간조직 내 지질 산화를 증가시키고 혈 중 중성지방 수치를 감소시킨다.15-17) 또한, 근육 내 AMPK 의 활성화는 PGC-1α 발현을 통해 미토콘드리아 관련 유전 자들의 발현을 증가시키는 것으로 보고되었다.18) 지방세포 에서 AMPK의 활성화는 ACC와 HSL (Hormone Sensitive Lipase)의 억제를 통하여 지방생성과 지방분해 모두를 억제 하는 기능이 보고된 바 있으나,19) 그 기전은 잘 알려져 있지 않다.
비만에 의한 염증반응은 지방세포 분비물의 변화에 기인 한다. 지방세포의 분비물 중에는 염증을 유발하는 인자가 포함되어 있는데, TNF-α와 interleukin (IL)-6가 대표적인 염증유발 아디포카인으로서, 대식세포를 유도하거나, 직접 적인 염증반응을 유발시킨다. 본 연구진은 염증반응을 유도 한 지방세포에서 Wnt signaling pathway 및 지방대사의 변 화를 관찰하였다. 또한 AMPK 활성화에 의한 지방세포 모 델에서 noncanonical Wnt 신호전달체계 및 지방대사의 변 화를 관찰하고자 본 연구를 시행하였다.
방 법 1. 재료 및 시약
Wnt5α, Wnt3α, Dvl3, JNK, phospho-JNK, β-catenin, GSK3-β, phospho- GSK3-β, TCF4, IRS1, phosphor-IRS1, Akt, phosphor-Akt, PPAR-γ, β-actin에 대한 항체는 Cell Signaling (Beverly, MA, USA)에서 구입하였고, sFRP5의 항체는 Abnova (Jhonggli, Taoyuan, Taiwan)에서 구입하여 실험하였다. AICAR는 Calbiochem (San Diego, CA)에서 구매하였고, 이외의 모든 시약은 Sigma사 (St. Louis. MO) 의 제품을 구입하여 사용하였다.
2. 3T3-L1 세포배양과 분화
3T3-L1 지방전구세포의 배양과 유지는 10% fetal bovine calf serum (FCS)을 넣은 Dulbecco’s modified Eagle’s medium (DMEM) 배지로 5% CO2, 37℃에서 50% 세포가 자랐을 때 계대배양을 하였다. 세포의 분화는 6-well plate 중심단어: 지방세포, 염증, Noncanonical Wnt 신호전달체
계, sFRP5, AMPK, TNF-α
Key words: Adipocyte, Inflammation, Noncanonical Wnt
signaling pathway, sFRP5, AMPK, TNF-α에 well 당 1 × 106 세포를 분주하여 세포가 100% 밀집되 게 배양하였다. 2일 후 10% fetal bovine serum (FBS)와 MDI solution (0.5 mM IBMX, 0.5 uM dexamethasone, 10 ug/mL insulin)을 포함한 DMEM 배지를 3일 동안 처리하 였고 다시 10% FBS와 10 ug/mL insulin을 포함한 DMEM 으로 배양하며 세포 내 지방방울의 형성을 근거하여 지방세 포로 분화시켰다. 모두 12일 동안의 분화를 통해 80% 이상 의 세포가 분화되었음을 확인하였다. 분화가 이루어진 3T3-L1 세포는 염증의 유발 및 회복을 유도하기 위하여 10 ng/mL의 TNF-α를 처리하거나, 10 ng/mL의 TNF-α와 1 mM 의 AICAR를 동시에 처리하였다.
3. 분비된 Glycerol의 측정
분화된 3T3-L1 세포에 시료를 처리한 후 48시간 배양한 배지를 eppendorf tube에 넣고 70℃에서 10분간 가열하여 세포로부터 유리된 효소들을 불활성화 시켰다. 50 uL의 배 지를 glycerol reagent에 첨가하여 1분간 반응시킨 후 microplate에 옮겨 넣고 흡광도 570 nm에서 측정하였다. 이 때 단백질량을 Bradford (Bio-rad, Hercules, CA)방법으로 정량하여 세포수를 보정하였다.
4. 실시간 역전사 중합효소연쇄반응법 (Real-Time Reverse Transcription Polymerase Chain Reaction (real-time RT-PCR))
시료의 투여에 따른 면역관련 인자 (inflammatory cytokines) 들의 발현 변화를 보기 위하여 분화된 3T3-L1 세포에 아무 것도 처리하지 않은 대조군과, TNF-α 혹은 TNF-α와 AICAR를 동시에 처리한 실험군에서, 24시간 후 RNA를 추출하여 RT-PCR를 시행하였다. RNA는 TRIZOL 용액 (Gibco-Brl, Grand Island, NY)을 사용하여 추출하였고, Molomey murine leukemia virus reverse transcriptase (MMLV-RT, Invitrogen, Carlsbad, CA)를 이용하여 cDNA 를 제작하였다. Real-Time PCR은 SYBR (Invitrogen, Carlsbad, CA)과 특이적인 primers (Bioneer Co. Daejon, Republic of Korea)를 사용하여 시행하였다. 사용된 primers는 다음과 같다.IL-6 (Interleukin-6)
Sense 5’-AGTTGCCTTCTTGGGACTGA-3’
Antisense 5’-TCCACGATTTCCCAGAGAAC-3’
MCP-1 (Monocyte chemoattractant protein-1) Sense 5’-ATGCAGGTCTCTGTCACG-3’
Antisense 5’-CTTCTGGACCCATTCCTTATTGG-3’
β-actin
Sense 5’-AGGTCATCACTATTGGCAAC-3’
Antisense 5’-ACTCATCGTACTCCTGCTTG-3’
각각의 cDNA 1uL, forward primer (10 pmol/uL) 1 uL, reward primer (10 pmol/uL) 1uL, SYBR Green q PCR kit Premix (×2) 용액 10 uL, 증류수 7 uL를 혼합한 후 Light-Cycler 480 (Roche, Lewis, UK)을 사용하여 각각의 유전자 조건에 맞게 PCR을 시행하였다.
5. Western Blot Analysis
Western blot을 위하여 세포를 Phosphate Buffered Saline으로 잘 세척한 후 SDS sample buffer (62.5 mM Tris-HCl (pH 6.8), 6% SDS, 30% glycerol, 125 mM DTT, 0.03% bromophenol blue)로 용해시키고, 세포를 긁 은 후 21 gauge needle을 통과시켜 geneomic DNA를 파괴 하고 원침하여 cell lysate를 만들었다. 단백질은 sodiumdodesyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE)에 의하여 분리되고, nitrocellulose membrane 으로 이동되었다. Membrane은 blocking buffer (1 × TBS, 0.1% Tween-20 with blocking reagent, 5% milk)에 담가서 일차항체들과 4℃에서 밤새 반응시켰다. Membrane을 HRP 가 중합된 이차항체에 120분간 반응시킨 후 세척한 다음 chemiluminescent detection system (New England Biolab.
Beverly, MA)을 이용하여 현상하였다
6. 자료처리 및 분석
단백질 정량은 Bradford법(Bio-rad, Hercules, CA)을 이 용하였고, 표준은 bovine serum albumin을 통행 측정하였 으며, glycerol release는 3회 반복하여 평균을 측정하였다.
TNF-α 및 AICAR에 처리에 의한 염증관련 인자의 실시간 역전사 중합효소 연쇄반응은 4회 반복하여 평균 ± 표준오차 로 표시하였고, 통계처리는 sigma plot program의 students t-test를 이용하여 P < 0.05 에서 유의성을 검정하였다.
결 과
1. 3T3-L1 지방세포에서 염증의 유발
지방세포에서 염증반응의 유발을 위하여 분화된 3T3-L1 지방세포에 염증 유발제인 TNF-α를 10 ng/mL의 농도로 24 시간 동안 처리하였다. 또한 염증반응에 대한 AMPK의 효 능을 관찰하기 위하여 AMPK 활성 유도제인 AICAR를 1uM의 농도로 TNF-α와 동시에 처리하였다. 대조군에는 TNF-α 및 AICAR의 처리용매인 dimethyl sulfoxide (DMSO) 를 동일한 양으로 처리하였다. 지방세포 내 염증 관련 인자 의 발현을 실시간 역전사 중합 효소법을 사용하여 측정한 결과, TNF-α 처리군에서 IL-6의 유전자 발현이 대조군에 비하여 약 19.11배 (P < 0.01) 증가하였고, MCP-1은 약 2.80 (P < 0.05)배 유의하게 증가하였다(Fig. 1A, B). 그러 나 TNF-α와 AICAR를 동시에 처리한 지방세포에서는 IL-6
가 대조군의 약 8.28배(P < 0.05), MCP-1은 약 0.57 배로 TNF-α (P < 0.05)를 처리한 세포보다 적은 양의 발현이 관 찰되었다. 이는 TNF-α에 의한 지방세포의 염증반응이 AMPK 활성화에 의하여 억제되었음을 시사한다.
2. 지방세포의 염증반응에 따른 Wnt 신호전달체 계의 활성 변화
본 연구진은 지방세포의 염증반응에 관련하여 Wnt 신호
전달체계의 활성변화를 관찰하였다. 몇몇의 Wnt 리간드 단 백질의 발현 변화를 확인하였는데, Noncanonical Wnt 신호 전달 리간드 단백질인 Wnt5a의 발현이 TNF-α 처리 시 약 1.4배 증가하였다(P < 0.05). Noncanonical Wnt 신호전달 관련 인자들 역시 현저히 증가되었다. Dvl3 (2.61 ± 0.25배;
P < 0.01)을 비롯하여 Noncanonical Wnt 신호전달의 필수 인자인 phospho-PKC (3.24 ± 0.30배; P < 0.01)와 phospho-JNK (2.14 ± 0.35배; P < 0.01) 역시 유의하게 증
Fig. 1. AMPK suppressed TNF-
α induced inflammatory gene expression. 3T3-L1 adipocyte was incubated with TNF-α (10 ng/mL) for 24 hrs either in the presence or absence of AICAR (1 mM). The gene transcript of IL-6 (A) and MCP-1 (B) was measured by real-time RT-PCR.* P < 0.05 and ** P < 0.01 compared with control group.
†P < 0.05 compared with TNF-α group.
Fig. 2. Noncanonical wnt signaling is specifically involved in inflammation and restoration in 3T3-L1 adipocyte. Expression of ligand
proteins and signaling molecules of noncanonical (A) and canonical (B) Wnt signaling pathway was evaluated by western blot assay.* P < 0.05 and ** P < 0.01 compared with control group.
†P < 0.05 and ††P < 0.01 compared with TNF-α group.
가하였다. 그러나 TNF-α에 의한 noncanonical Wnt 신호전 달체계의 활성 증가는 AICAR 처리에 의하여 회복되는 양 상을 보였다. TNF-α에 의하여 증가하였던 Wnt5a, Dvl3, pPKC, pJNK의 발현은 AICAR의 동시 처리에 의해 대조군 수준의 발현량으로 회복되었다(Fig. 2A). Canonical Wnt 신 호전달 리간드인 Wnt3a, Wnt10a의 발현은 Wnt5a와 유사 한 변화를 보였지만, 유의성 있는 변화는 관찰되지 않았다.
Phospho-GSK3β 및 β-cantenin을 포함한 canonical Wnt 신 호전달 인자들 역시 TNF-α, AICAR에 의한 염증, 회복과정에 서 변화하지 않았다(Fig 2B). 이는 지방세포에서 TNF-α 및 AICAR에 의한 염증 및 회복과정에 noncanonical Wnt 신 호전달체계가 관여함을 제시한다.
3. 지방세포의 염증반응에 의한 인슐린 신호전달의 변화
인슐린 신호전달체계의 활성화 정도는 인슐린-인슐린 수 용체 간의 결합에 의하여 직접 활성화되는 insulin receptor substrate (IRS)-1과 하위 신호전달체계 인자인 Akt의 인산 화 정도를 측정하여 나타내었다. 인슐린 저항성의 잠재적인 매개자로 알려진 IRS-1의 serine 302의 인산화 정도를 측정 하였는데, TNF-α에 의하여 1.7배(P < 0.05) 증가하였다 (Fig. 3A). Akt의 활성화를 유도하는 threonine 308의 인산 화 정도 역시 감소하는 경향을 보였다(Fig. 3B). 그러나 AMPK 활성 유도제인 AICAR와 TNF-α를 동시에 처리한 경우, IRS-1 serine 302의 인산화는 현저히 감소하는 양상 을 보였고, Akt의 활성화 역시 증가하였다.
4. 염증반응에 의한 지방대사의 변화
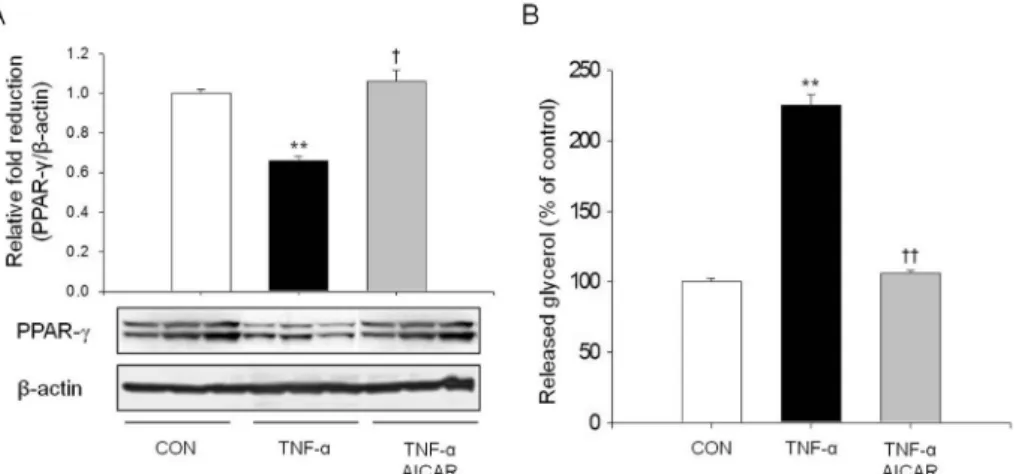
지방세포에서 TNF-α에 의한 염증반응은 다양한 신호전
달 체계를 활성화 시키거나, 저해시킨다. 따라서 본 연구진 은 지방세포 내 지방대사의 변화를 확인하고자, 지방합성을 포함한 지방대사에서 가장 중요한 인자인 PPAR-γ의 양적 변화를 관찰하였다. 또한 지방분해의 변화를 관찰하기 위하 여 세포에서 분비된 glycerol의 양을 측정하였다. TNF-α의 처리는 PPAR-γ의 발현을 현저히 억제하였다(Fig. 4A). 반 면에 세포 외로 분비된 glycerol의 양은 대조군의 비하여 약 2배(P < 0.01) 증가하였다(Fig. 4B). 이는 TNF-α에 의한 염 증의 유발이 지방대사에 영향을 미치고, 특히 지방분해를 촉진할 수 있음을 시사한다. TNF-α에 의한 PPAR-γ의 발현 감소 및 분비된 glycerol의 증가는 AICAR 처리에 의하여 회복되었다.
5. AMPK에 의한 항염증인자의 분비증가
본 연구진은 AMPK의 인위적인 활성화가 TNF-α에 의한 noncanonical Wnt 신호전달을 억제함에 따라, AMPK에 따 른 항 Wnt 신호전달 물질(anti-Wnt signaling molecule) 의 양적 변화를 관찰하였다. 따라서 세포 외 공간에서 Wnt 단 백질과 수용체의 결합을 방해하는 secreted frizzled receptor protein 5 (sFRP5)에 주목하였다. 지방세포 내에 sFRP5의 발현은 TNF-α에 의하여 오히려 증가하였고, AICAR 동시 처리에 의하여 대조군 수준으로 감소하였다. 그러나 배양액 으로 분비된 sFRP5의 양을 측정해본 결과, TNF-α와 AICAR를 동시에 처리한 세포의 배양액에서 TNF-α를 단독 처리한 세포보다 현저히 증가하였다(Fig. 5A). 분비된 sFRP5 단백질의 비율(분비된 sFRP5의 양/세포 내 sFRP5의 양)은 TNF-α에 의하여 감소하였으나, AICAR에 약 3배(P< 0.05) 증가하였다. 이상의 결과는 지방세포에서 TNF-α에
Fig. 3. AMPK prevented TNF-
α induced insulin signaling impairment in 3T3-L1 adipocyte. The key molecules of insulin signaling, pIRS-1 (A), pAkt (B) were detected by a specific antibody. Total protein and β-actin were used for quantitative analysis.* P < 0.05 compared with control group.
††P < 0.01 compared with TNF-α group.
의한 염증반응에 의한 Wnt5a의 증가에 따라 noncanonical Wnt 신호전달체계가 증가되고, AICAR에 의한 AMPK의 활성화가 sFRP5의 분비를 촉진시켜 Wnt 신호전달체계를 억제함을 제시하고 있다.
고 찰
지방조직은 과거에는 잉여에너지의 단순한 저장고로 여 겨졌지만 최근에는 식욕, 체온조절, 면역 및 신경호르몬 기 능을 조절하는 adipokine을 분비하여 에너지 대사의 항상성 에 기여하고 있는 내분비 기관으로 인식되고 있다. 그 중에 는 adiponectin, leptin과 같은 항대사질환 기능의 아디포카 인도 있지만 TNF-α, IL-6와 같이 염증을 일으키거나 인슐 린 저항성을 증가시키는 싸이토카인도 있다. 특히 TNF-α의
발현은 비만과 같은 제 2형 당뇨와 같은 대사질환을 가진 개체의 지방세포에서 증가하며, 분비 시 지방조직을 포함한 다양한 조직에서 염증 반응을 일으킨다.
Wnt 신호전달체계는 최초에 암 유발에 관련이 연구되었 으나, 최근 들어서 Wnt 신호전달체계가 다양한 질병과의 연 관성이 밝혀지고 있다. 그 중 지방세포의 형성(adipogenesis) 와 관련하여 Wnt 신호전달에 의한 지방세포의 형성 억제에 관한 연구가 많이 진행되어 왔는데, 지방전구세포에서 발현 되는 Wnt10b는 세포 내 조절인자로서 지방세포 분화에 중 요 전사인자인 C/EBPα와 PPARγ의 발현을 억제함으로써 지방세포 특이 유전자가 발현되지 못해 지방세포 형성을 막 는다.11,12)
Noncanonical Wnt 신호전달체계는 GSK3β와 β-catenin 의 신호전달 체계를 매개로 하지 않으며, 특정 Wnt/Fz의 결
Fig. 4. TNF-
α and AMPK regulated PPAR-γ expression and lipolysis. The amount of PPAR-γ protein (A) and released glycerol (B) to medium was estimated by western blot and free glycerol reagent.** P < 0.01 compared with control group.
†P < 0.05 and ††P < 0.01 compared with TNF-α group.
Fig. 5. AMPK stimulated sFRP5 secretion in 3T3-L1 adipocyte. The amount of sFRP5 in medium and cell lysate was measured by
western blot (A). The secretion rate was described as a ratio of secreted sFRP5 / non-secreted sFRP5. (B)* P < 0.05 compared with control group.
†P < 0.05 compared with TNF-α group.
합에 의한 세포 내 칼슘의 증가로 PKC나 calmodulin dependent kinase (CamKⅡ)와 같은 Ca2+-sensitive enzyme 이 활성화되어 신호가 전달되거나, 혹은 small GTPase의 일종인 rho나 cdc42를 통해서 JNK의 활성화가 이루어지는 경로이다.20,21) 본 연구에서 지방세포주인 3T3-L1에 TNF-α 의 처리는 IL-6와 MCP-1 유전자의 발현을 증가시키며, 염 증을 유도하였다. 지방세포 내 염증의 유발은 noncanonical Wnt 신호전달체계를 활성화 시키면서, Wnt5a 단백질의 발 현 증가와 PKC 및 JNK의 활성화를 유도하였다. Wnt5α는 대표적인 noncanonical Wnt 리간드 단백질로서, Takada23) 등에 의하여 지방세포 내 지방대사의 중요 인자인 PPAR-γ 의 전위 활성화(transactivation)를 억제하는 기능으로 보고 된 바 있다. 이러한 결과는 noncanonical Wnt 신호전달체계 가 지방세포 내 지방대사의 변화를 매개하는 신호전달 체계 임을 시사하고 있다. 본 연구의 결과에서도, noncanonical Wnt 신호전달의 활성화가 PPAR-γ의 억제 및 지방분해의 활성화를 동반하며 지방대사의 이상을 초래함을 증명하고 있다. 그러나 TNF-α에 의한 염증반응에서 canonical Wnt 신호전달은 변화가 관찰되지 않아, 지방세포 형성과정과는 다르게 성숙지방세포의 지방대사에서 noncanonical Wnt 신 호전달체계의 중요성을 제시하고 있다.
Wnt 신호전달체계는 Wnt 리간드-수용체 간의 결합에 의 하여 활성화되는 신호전달체계로 몇몇의 세포 외 인자들 (extracellular molecules)에 의하여 조절된다. sFRPs는 대표 적인 Wnt 신호전달체계의 조절 인자로서, 조직 및 세포에 따라 sFRP1, 2, 3, 4, 5의 단백질이 동정되어 있고, 현재까 지는 단순한 Wnt 신호전달체계의 억제제로서 보고되었 다.27) 그러나 최근 비만 및 제2형 당뇨병 생쥐모델에서 sFRP5의 발현이 현저히 감소됨과 동시에, 지방세포에서 염 증 관련 아디포카인의 분비증가 및 JNK의 활성화를 보이면 서, sFRP5의 항염증 작용이 보고되었다.24) 본 연구에서 지 방세포 내 인위적인 AMPK의 활성화는 sFRP5의 분비를 현저히 촉진하였고, 이는 Wnt5α의 발현감소 및 전체적인 noncanonical Wnt 신호전달체계의 저해를 유도하였다. 또 한 AMPK의 활성화는 TNF-α에 의한 염증 관련 인자의 발 현을 억제하고, 인슐린 신호전달 체계 및 지방대사를 유의 하게 회복시키며, 지방세포의 보호 기능이 관찰되었다. 이는 지방세포에서 AMPK의 활성화가 sFRP5의 분비를 촉진하 여, noncanonical Wnt 신호전달체계를 억제하고, 이에 따라 지방세포의 염증반응 및 지방대사의 변화가 회복된다는 가 능성을 제시한다
본 연구를 통하여 3T3-L1 지방세포주의 염증 유발 시 AMPK 활성화를 통해 염증 억제 및 지방대사의 개선을 보여주 었고, 이러한 효과는 염증에 의하여 활성화되는 noncanonical Wnt 신호전달체계의 억제에 의하여 이루어짐을 유추할 수 있었으며, 항 Wnt 신호전달 아디포카인(anti-Wnt signaling
adipokine)인 sFRP5과 관련이 있음을 확인하였다. 현재 비 만이나 제2형 당뇨병과 같은 대사질환에서 지방조직에 대한 중 요성은 나날이 강조되고 있다. 본 연구의 결과는 noncanonical Wnt 신호전달체계가 지방세포의 지방대사에 매우 중요한 역할을 가지고 있음을 제시하고, 대사질환의 치료와 관련하 여 sFRP5가 중요한 표적단백질이 될 수 있음을 시사한다.
참 고 문 헌
1. Spiegelman BM, Flier JS. Adipogenesis and obesity;
rounding out the big picture. Cell 1996;87:377-89.
2. Kopelman PG. Obesity as a medical problem. Nature 2000;404:635-43.
3. Visscher TL, Seidell JC. The public health impact of obesity. Annu Rev Public Health 2001;22:355-75.
4. Frayn KN, Karpe F, Fielding BA, Macdonald IA, Coppack SW. Integrative physiology of human adipose tissue. Int J Obes Relat Metab Disord 2003;27:875-88.
5. Holm C. Molecular mechanisms regulating hormone sensitive lipase and lipolysis. Biochem Soc Trans 2003;31:1120-4.
6. Halle M, Berg A, Northoff H, Keul J. Importance of TNF-alpha and leptin in obesity and insulin resistance: a hypothesis on the impact of physical exercise. Exerc Immunol Rev 1998;4:77-94.
7. Akter MH, Yamaguchi T, Hirose F, Osumi T. Perilipin, a critical regulator of fat storage and breakdown, is a target gene of estrogen receptor-related receptor alpha.
Biochem Biophys Res Commun 2008;368:563-8.
8. Reya T, Clevers H. Wnt signalling in stem cells and cancer. Nature 2005;434:843-50.
9. Moon RT, Kohn AD, De Ferrari GV, Kaykas A.
WNT and beta-catenin signalling: diseases and therapies.
Nat Rev Genet 2004;5:691-701.
10. Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol 2004;20:781-810.
11. Ross SE, Hemati N, Longo KA, Bennett CN, Lucas PC, Erickson RL, et al. Inhibition of adipogenesis by Wnt signaling. Science 2000;289:950-3.
12. Liu J, Farmer SR. Regulating the balance between peroxisome proliferator-activated receptor gamma and beta-catenin signaling during adipogenesis. A glycogen synthase kinase 3beta phosphorylation-defective mutant of beta-catenin inhibits expression of a subset of adipogenic genes. J Biol Chem 2004;279:45020-7.
13. Moldes M, Zuo Y, Morrison RF, Silva D, Park BH, Liu J, et al. Peroxisome-proliferator-activated receptor gamma suppresses Wnt/beta-catenin signaling during adipogenesis. Biochem J 2003;376:607-13.
14. Assifi MM, Suchankova G, Constant S, Prentki M, Saha AK, Ruderman NB. AMP-activated protein kinase and coordination of hepatic fatty acid metabolism of starved/carbohydrate-refed rats. Am J Physiol Endocrinol Metab 2005;289:E794-800.
15. Bergeron R, Previs SF, Cline GW, Perret P, Russell RR 3rd, Young LH, et al. Effect of 5-aminoimidazole -4-carboxamide-1-beta-D-ribofuranoside infusion on in vivo glucose and lipid metabolism in lean and obese Zucker rats. Diabetes 2001;50:1076-82.
16. Zhou G, Myers R, Li Y, Chen Y, Shen X, Fenyk -Melody J, et al. Role of AMP-activated protein kinase in mechanism of metformin action. J Clin Invest 2001;108:1167-74.
17. Foretz M, Ancellin N, Andreelli F, Saintillan Y, Grondin P, Kahn A, et al. Short-term overexpression of a constitutively active form of AMP-activated protein kinase in the liver leads to mild hypoglycemia and fatty liver. Diabetes 2005;54:1331-9.
18. Jager S, Handschin C, St-Pierre J, Spiegelman BM.
AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylation of PGC-1alpha. Proc
Natl Acad Sci U S A 2007;104:12017-22.
19. Corton JM, Gillespie JG, Hawley SA, Hardie DG.
5-aminoimidazole-4-carboxamide ribonucleoside: a specific method for activating AMP activated protein kinase in intact cells? Eur J Biochem 1995;229:558-65.
20. Pandur P, Maurus D, Kuhl M. Increasingly complex:
new players enter the Wnt signaling network.
Bioessays 2002;24:881-4.
21. Sheldahl LC, Slusarski DC, Pandur P, Miller JR, Kuhl M, Moon RT. Dishevelled activates Ca2+ flux, PKC, and CamKII in vertebrate embryos. J Cell Biol 2003;161:769-77.
22. Takada I, Mihara M, Suzawa M, Ohtake F, Kobayashi S, Igarashi M, et al. A histone lysine methyltransferase activated by non-canonical Wnt signalling suppresses PPAR-gamma transactivation.
Nat Cell Biol. 2007;9:1273-85.
23. Bovolenta P, Esteve P, Ruiz JM, Cisneros E, Lopez-Rios J. Beyond Wnt inhibition: new functions of secreted Frizzled-related proteins in development and disease. J Cell Sci 2008;121:737-46.
24. Ouchi N, Higuchi A, Ohashi K, Oshima Y, Gokce N, Shibata R, et al. Sfrp5 is an anti-inflammatory adipokine that modulates metabolic dysfunction in obesity. Science 2010;329:454-7.