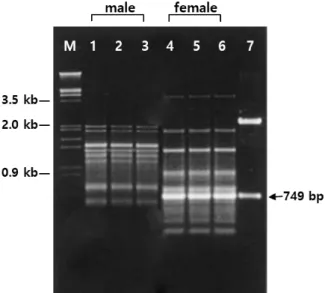
Development of a Female-associated SCAR Marker in Schisandra nigra Max.
Hyo Shim Han
1and Jae Sung Jung
2*
1
Liberal Arts Education Center, Sunchon National University, Suncheon 57922, Korea
2
전체 글
1
2
수치

관련 문서
§ To develop new mesh-type reference Korean phantoms (MRKPs) for adult male and female to address limitations of current voxel-type reference Korean adult phantoms...
• Theory can extent to molten polymers and concentrated solutions.. The single-molecule bead spring models.. a)
Among the sub-factors of self-esteem, positive, male was higher than female, and on the contrary, negative In this study, females were higher than males,
caryatid: sculpted female figure used in place of a column.. Components of Greek Orders.. Doric Temple of Athene Aphaia, Aegina Ionic Temple, Metaponte.. Between
The results of analysis on the difference of psychological happiness according to frequency of participation of external physical activity of female middle school
The study photgraphs of male dentist dressed in formal attire (A) white coat1. The study photgraphs of female dentist dressed in formal attire (A)
In this study, it is showed that female and blood type O are the significant factors associated with prolonged C/Epi CT value despite of aspirin discontinuation more
Third, the analysis of the body-centered vertical speed between male.. and female students showed that male students appeared faster than female students at
