Conditioned Media of RAW 264.7 Cells Stimulated with Phellinus linteus Extract Regulates the Epithelial-mesenchymal Transition in Prostate Cancer Cells
Taewoo Kang
1,2,3†, Hyun-Hee An
4†, Sul-Gi Park
4, Sun-Nyoung Yu
4, You-Lim Hwang
4, Ji-Won Kim
4and Soon-Cheol Ahn
4*
1Department of Surgery (Busan Cancer Center), Pusan National University Hospital, Busan 49241, Korea
2Biomedical Research Institute, Pusan National University Hospital, Busan 49241, Korea
3Pusan National University Hospital, Busan 49241, Korea
4Department of Microbiology and Immunology, Pusan National University School of Medicine, Yangsan 50612, Korea
Received July 22, 2019 /Revised August 26, 2019 /Accepted August 27, 2019
Prostate cancer (PCa) is one of the most metastatic tumor. Although hormone therapy or surgical cas- tration is mostly conducted to treat PCa, it has a lot of side effects. Recently, many researchers have been exploring the tumor microenvironment to remedy these circumstances. Immune cells, especially macrophages, are an important composition of the tumor microenvironment. Under normal con- ditions, macrophages exhibit mild tumoricidal activity against tumors. However, once activated by in- terferon gamma or lipopolysaccharides, macrophages can kill cancer cells directly or indirectly by se- creting cytokines and chemokines. In this study, murine macrophage RAW 264.7 cells were treated with Phellinus linteus extract. To analyze their pro-inflammatory phenotype, we were used several as- says such as a real-time polymerase chain reaction, an enzyme-linked immunosorbent and nitric oxide assay. Prostate cancer cells were treated with the RAW 264.7-conditioned media, which was identified as a pro-inflammatory nature, for 48 h, and the expression of epithelial-mesenchymal transition (EMT)-related genes was determined. Not only N-cadherin, Snail, Twist, Slug, and Cadherin 11, which are mechenchymal-related proteins, were decrease, but epithelial marker of E-cadherin was increased.
In addition, the mRNA level of vimentin, ccl2, and vegfa were decreased, as the EMT is closely related to the migration and invasion of cancer cells. In conclusion, the RAW 264.7-conditioned media stimu- lated with P. linteus extract inhibited migration and invasion and regulated the EMT pathway in hu- man prostate cancer cells.
Key words : Conditioned media of murine macrophage RAW 264.7 cells, EMT pathway, human
prostate cancer PC-3 cells, Phellinus linteus
†Authors contributed equally.
*Corresponding author
*Tel : +82-51-510-8092, Fax : +82-55-382-8090
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2019 Vol. 29. No. 8. 904~915 DOI : https://doi.org/10.5352/JLS.2019.29.8.904
Introduction
Cancer is a malignant tumor formed by abnormal growth of deformed cells when the normal cell undergoes a variety of genetic mutation caused by external stimulators. Cancer is the leading cause of death in the world. It is regarded as one of the most important factors threatening human life worldwide [4]. In Korea, the mortality rate by cancer continues to increase [7]. Prostate cancer is the most com- mon male cancer in the United States and the second lead-
ing cause of cancer-related deaths [9]. In Korea, the fre- quency and mortality rate are increasing due to changes in lifestyle such as westernized eating habits, and aging [12].
The incidence of prostate cancer varies according to genetic factors, race, and age [38, 41]. According to US statistics, prostate cancer is rare in males less than 45 years old, but the incidence is higher after age 50. It is reported that the probability of developing prostate cancer in men over 50 years old is about 30% and that in men over 80 years old is about 80%[45].
Macrophages are known to be involved in host defense
and homeostasis by participating in various host responses
such as acquired immunity as well as natural immunity. In
the inflammatory response, it produces cytokines such as
interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), and
IL-6 in the early stage of infection. In cancer patients, macro-
phages infiltrate cancer cells and appear as tumor-associated
macrophages (TAMs). TAMs play an important role in the
relationship between inflammation and cancer, which is ba- sically an important influence on disease progression. Stu- dies have shown that macrophages are likely to kill tumor cells if properly activated, by interferon-γ (IFN-γ) and lip- opolysaccharides (LPS) therapy [1].
The epithelial-mesenchymal transition (EMT) is a key de- velopmental program that is often activated during cancer invasion and metastasis. EMT is a biologic process that ap- proves a polarized epithelial cell, which normally interacts with basement membrane via its basal surface, to undergo multiple biochemical changes that enable it to assume a mes- enchymal cell phenotype, which causes enhanced migratory capacity, invasiveness, elevated resistance to apoptosis and greatly increased production of extracellular matrix compo- nents [2]. A number of biomarkers of EMT have been found in recent studies, such as E-cadherin, N-cadherin, vimentin, Snail, Zeb1, and Twist [3]. E-cadherin is located on the cell surface of epithelial tissue and mediates cell-cell adhesion to bind cells in normal epithelial cells. E-cadherin expression is negatively correlated with the occurrence of EMT and tu- mor invasion [5]. In contrast, vimentin (cytoskeleton marker) and N-cadherin (cell surface markers) are associated with the onset of EMT and progression to invasive carcinomas.
Snail, Slug, Zeb1, Zeb2, and Twist can also lower E-cadherin level and consequently trigger EMT [6]. E-cadherin, N-cad- herin, Zeb1, Twist, Slug, Snail, and other EMT markers play an important role in the invasion of prostate cancer cells and the regulation of their metastatic potential. Therefore, a therapeutic strategy that intervenes the EMT process or reverses the EMT phenotype may be an alternative to cancer treatment in the future [5].
Conventional anti-cancer drugs cause side effects such as immune function deterioration, hematopoietic disorder, hair loss, and genetic damage. Recently, researches are being con- ducted to develop anti-cancer agents based on natural prod- ucts that act specifically on cancer cells with lower side ef- fects and reducing adverse effects [8]. In modern society, the function of food is classified as a primary function of nutrition, a secondary function of taste and palatability, and a tertiary function of biological control which helps to pre- vent and cure disease [10]. Since our food ingredients are not genotoxic and cytotoxic. Efforts to develop excellent therapeutic agents from everyday natural products have been made worldwide [11]. Phellinus linteus is a white rot fungus that belongs to the Hymenochaetaceae Phellinus. The similar species are P. gilvus, P. isniarius, P. nigricans, P. pini,
P. conchaius, P. densus, and P. hatigii. Among them, P. linteus
has become a target of mushrooms with novel anti-cancer activity [13]. Many studies have been carried out on the hot water extract of fruit body of P. linteus as it has a blocking effect on gastrointestinal cancer [14]. Also it has an enhance- ment effect on the immune function when the chemotherapy is treated onto the cancer of the gastrointestinal system in- cluding gastric cancer, esophageal cancer, colon cancer, and rectal cancer, as well as liver cancer surgery [14]. It has been reported that the polysaccharide extracted from fruiting body of artificially cultured P. linteus or natural P. linteus has anti-complement acticity, which increased the useful val- ue of P. linteus [15].
In this study, we observed that P. linteus induces RAW 264.7 cells into pro-inflammatory macrophages and the con- ditioned media regulate the expression of migration, in- vasion, and EMT related genes in human prostate cancer, PC-3 cells.
Materials and Methods
Chemicals and Materials
Phellinus linteus was purchased from Kumwhangbio Co.
(Jinju, Korea). Diaion HP-20 was purchased from the Mitsu- bishi Kasei Co. (Tokyo, Japan). Lipopolysaccharide (LPS) and polymyxin B sulfate salt (PB) were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). Antibodies of E- cadherin, vimentin, and snail-1 were purchased from Cell Signaling Technology (Victoria, BC, Canada), CCL2 from Bio X Cell (BxCell, West Lebanon, NH, USA), β-actin from Santa Cruze Biotechnology (Santa Cruz, CA, USA). And goat-an- ti-rabbit IgG secondary antibody was purchased from Enzo Life Science (Farmingdale, NY, USA). Western blot detection was performed using an enhanced chemiluminescence re- agent (WesternBright ECL, Advansta, Menlo Park, CA, USA). N-(1-Naphthyl) ethylenediamine dihydrochloride and sulphanilamide were purchased from Sigma-Aldrich Co.
Extraction of Phellinus linteus
The dried fruit bodies of P. linteus were crushed by a
grinder. Grinded fruit bodies of P. linteus were extracted
with hot water (PLE) at 100℃ for 20 mins. PLE solution was
centrifuged at 13,000 rpm for 10 mins and its supernatant
was used for the experiments. Diaion HP-20 is one of the
adsorption resins, which binds the unwanted lipophilic com-
pounds and concentrates the target substances from a large
Table 1. Schematic process for purification of bioactive com- pounds from Phellinus linteus
amount of aqueous solution. Diaion HP-20 column was used to separate the active portion of the PLE supernatant. The PLE was passed from Diaion HP-20 resin to remove the ad- sorbed material, yielding two fractions of Phellinus linteus hot water extract/passing (PLEP), which was not adsorbed on resin and Phellinus linteus hot water extract/elution (PLEE), which was adsorbed on resin and eluted with meth- anol (MeOH) (Table 1). All fractions were dried in Modul 4080C speed vacuum (Hanil Science Industrial Co., Gimpo, Korea) and solvent-free residue was quantitatively re-dis- solved in water or MeOH, respectively.
Cell lines and culture
Mouse macrophage cell line, RAW 264.7 and human pros- tate cancer cell lines, PC-3 and DU145, were cultured in Dulbecco’s modified Eagle’s minimal medium (DMEM, WelGENE Inc. Daejon, Korea.) supplemented with 10% fetal bovine serum (FBS, WelGENE Inc.), and 100 unit/ml of pen- icillin, and streptomycin (WelGENE Inc.). Cells were cul- tured in a humidified atmosphere with 5% CO₂ at 37℃.
For the activation of macrophages, RAW 264.7 cells were treated with 100 μg/ml of PLEP or 1 μg/ml of LPS as a positive control for 24 hr. The prostate cancer cells were treated with the conditioned media of RAW 264.7 cells stimulated with or without LPS or PLEP for 48 hr.
Measurement of nitric oxide
Nitrite accumulation was used as an indicator of nitric oxide (NO) production. Quantification of NO was examined in murine macrophage RAW 264.7 cells by the Griess reaction. Griess reagent contains 0.2% naphthylethylenedi- amine dihydrochloride in DW and 2% sulfanilamide in 5%
phosphoric acid. RAW 264.7 cells were dispensed into 96- well plates and stimulated with 10, 30 or 100 μg/ml of PLE, PLEP and PLEE for 24 hr. The supernatants were collected
and mixed with 0.5 volumes of Griess reagent and incubated at room temperature for 10 mins. The plates were measured at 540 nm in a VERSA
maxmicroplate reader (Molecular Devices, Sunnyvale, CA, USA).
Cell viability analysis
The cell viability was determined by the colorimetric MTT assay. Prostate cancer PC-3 and DU145 cells were seeded in 48-well plates at a density of 1×10
4/well and treated with the conditioned media of RAW 264.7 cells for 24 or 48 hr.
0.5 mg/ml MTT solution was treated and incubated at 37℃
for 3 hr. The supernatant was removed by suction and the dyed precipitates were dissolved in dimethyl sulfoxide. The optical density was measured at 570 nm in a microplate.
Wound healing assay
This method mimics cell migration during wound healing
in vivo. Wound healing assay were carried out as describedpreviously [16]. Briefly, PC-3 cells were seeded at high con- centration in a 6-well plate and grown overnight until the density reaches 80%. The monolayer was scratched using pipette tips. And the floating cells were removed and the conditioned media of RAW 264.7 cells was treated along with new media. Migration of PC-3 cells into scratched areas was observed by an optical microscope CKX41 (Olympus, Tokyo, Japan).
Reverse transcriptase (RT)-PCR and quantitative PCR (qPCR) analysis
Using a GeneAll
RRiboEx RNA extraction kit (GeneAll
Biotechnology, Seoul, Korea), total RNA from RAW 264.7
cells and PC-3 cells was isolated in accordance with the man-
ufacturer's specifications. The purity and concentration of
total RNA were measured by a NanoDrop spectropho-
tometer ND-8000 (NanoDrop Technologies, Oxfordshire,
UK). The Cdna was synthesized using by TOPscript ™ RT
DryMIX (Enzynomics, Daejeon, Korea) according to the
manufacturer's instructions. qPCR was performed using
TOPreal ™ qPCR 2X PreMIX (Enzynomics). All primer se-
quences for qPCR analysis were described in Table 2. The
PCR reactions consisted of pre-incubation process, 3 steps
of amplification processes, and melting process. The pre-in-
cubation process was carried out at 95℃ for 600 secs and
the 3 steps of amplification processes were performed at 9
5℃ for 10 secs, at 60℃ for 10 secs, at 72℃ for 10 secs and
finally a total of 45 cycles. The melting process was carried
Table 2. List of human- and mouse-originated primers and antibodies used in this study Mouse origin primers
Symbol Forward primer (5'→3') Reverse primer (5'→3')
iNOS IL-1β IL-6 COX2 TNFα GAPDH
CAG CTG GGC TGT ACA AAC CTT GCC GGG AAG ACA ATA ACT GC GAG GAT ACC ACT CCC AAC AGA CC
GGG TGT CCC TTC ACT TCT TTC A CAT CTT CTC AAA ATT CGA GTG ACA A
AAG GTC ATC CCA GAG CTG
CAT TGG AAG TGA AGC GTT TCG TTC AGC TTC TCA CCC AGG GA AAG TGC ATC ATC GTT GTT CAT ACA
TGG GAG GCA CTT GCA TTG A TGG GAG TAG ACA AGG TAC AAC CC
GCC ATG AGG TCC ACC ACC CT Human origin primers
Symbol Forward primer (5'→3') Reverse primer (5'→3')
E-cadherin N-cadherin
Twist Slug Cadherin 11
Snail Vimentin
CCL2 VEGFA GAPDH
ACA GCC CCG CCT TAT GAT T CAT CCC TCC AAT CAA CTT GC TCT CAA GAG GTC GTG CCA AT AAG CCA AAC TAC AGC GAA CTG
CTC CCA GGG AAG ACA TGA GA CCC CAA TCG GAA GCC TAA CT AGT CCA CTG AGT ACC GGA GAC GCT CAT AGC AGC CACA CTT CAT TC
AAG GAG GAG GGC AGA ATC AT CCA CCC ATG GCA AAT TCC ATG GCA
TCG GAA CGC TTC CTT CA ATG TGC CCT CAA ATG AAA CC
ATG GTT TTG CAG GCC AGT TT CTG AGC TGA GGA TCT CTG GTT
ACT CTC TGT AGC CAC CAC AT GCT GGA AGG TAA ACT CTG GAT TAG A
CAT TTC ACG CAT CTG GCG TTC GGA CAC TTG CTG CTG CTG ATT C
CCT TTC CCT TTC CTC GAA CT TCT AGA CGG CAG GTC AGG TCC ACC
out at 95℃ for 10 secs, 65℃ for 60 secs, and 95℃ for 30 secs for melting curve. All RT-PCR reactions were performed in triplicate and mRNA expression was normalized to glyc- eraldehyde 3-phosphate dehydrogenase (GAPDH).
Western blotting
PC-3 cells were cultured in a 6-well plate at a density of 2×10
5/well and incubated with the conditioned media of RAW 264.7 cells stimulated with 100 μg/ml of PLEP for 48 hr. The cells were detached from the plate surface with tryp- sin-EDTA. The cells were lysed in PRO-PREP protein ex- traction solution (iNtRON Biotechnology, Sungnam, Korea).
The cell lysates were quantified by Bradford method and separated on sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The gels were transferred to polyvinylidene fluoride membranes. The membranes were blocked with 5%
skim milk in TBS-T buffer [0.1% Tween 20, 150 mM NaCl, 20 mM Tris (pH 7.4)] at room temperature for 1 hr. The membranes were probed with 5% skim milk containing spe- cific primary anti-body at 4℃ overnight. Finally, the mem- branes were detected with ECL western blotting detection reagent. Each band was determined by using a fluorescence scanner (LAS 3000, Fuji Film, Tokyo, Japan) and analyzed with Multi Gauge V3.0 software (Fuji Film).
Enzyme-linked immunosorbent (ELISA) assay ELISA was performed using ELISA Ready-SET-Go!
(eBioscience, Seoul, Korea) according to the manufacturer's specifications. In brief, the capture antibodies were coated to 96-well immune-plates and incubated at 4℃ overnight.
The plates were washed with 200 μl of PBS containing 0.1%
Tween 20 (PBST). Then plates were blocked with 1% BSA for 1 hr at 37℃. Standard and culture supernatants were added into wells and incubated for 3 hr at 37℃. After 3 hr, plates were washed five times with PBST. Then, 100 μl of biotin-conjugated anti-mouse detection antibodies were added into each wells and incubated for 1 hr at room temperature. After 1 hr, the plates were washed five times with PBST. Then avidin-conjugated hourseradish peroxidase were added and incubated for 30 mins. Tetra-methyl-benzi- dine was used for a substrate. The reaction was stopped by H
2SO
4solution.
Annexin-Ⅴ/propidium iodide (PI) staining
Analysis of apoptotic cells was measured by an FITC
Annexin-Ⅴ/PI detection kit. PC-3 cells were treated with
the conditioned media of RAW 264.7 cells stimulated with
PLEP for 48 hr and harvested cells were washed with 1x
PBS. Then collected cells were resuspended in 1x binding
buffer (100 μl) and stained with Annexin-Ⅴ/PI staining sol-
A B
C
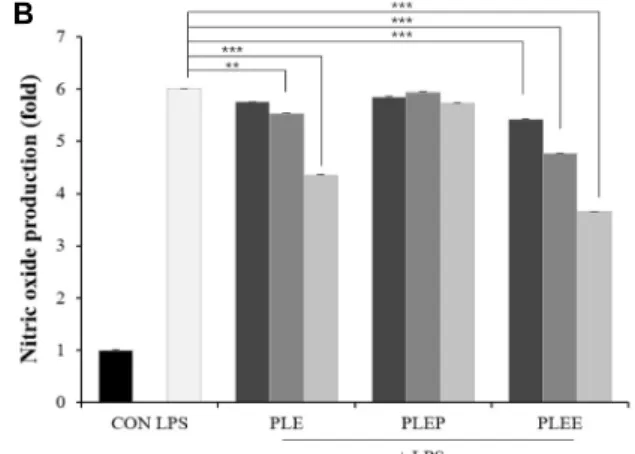
Fig. 1. Effects of PLE and its fractions on nitric oxide production in RAW 264.7 cells. (A) Induction of NO productions; (B) Inhibition of NO production; (C) Effect of polymyxin B.
CON, untreated negative control; LPS, 1 μg/ml of lip- opolysaccharide; PLE, P. linteus hot water extract; PLEP, P.
linteus hot water extract/passing; PLEE, P. linteus hot water extract/MeOH elution. Data are presented as mean ± SD (n=3 in each group). **p<0.01, ***p<0.001 vs. CON group.
ution at room temperature for 15 mins. Finally, the stained cells were analyzed by a flow cytometry (FACS Calibur, Becton Dickinson Co. CA, USA) and quantitative analysis of apoptotic cells was performed by the Cell Quest software (Becton Dickinson Co.)
Statistical analysis
All experiments were performed at least three times.
Unless otherwise stated, the data are expressed as means
± SD. Differences between the three groups were performed using by ANOVA, followed by Tukey-Kramer multiple com- parison tests. Results were statistically significant at *p<0.05.
Results
Various fraction of Phellinus linteus induces NO production in RAW 264.7 cells
PLE is the extract derived from natural product by hot water extraction. Therefore, it may contain both compounds to increase and decrease NO production. Diaion HP-20 col- umn was used to separate some compounds, which adsorbs substances with molecular weight of 1,500 or more and hy- drophobic character. By fractionation with Diaion HP-20, the passing fraction, PLEP, increased NO production and, on the
contrary, the elution fraction with methanol, PLEE, reduced NO production induced by LPS (Fig. 1A, Fig. 1B). NO pro- duction in PLEP was higher than that in PLE, because of the molecules that reduce NO production were collected in the elution fraction. Therefore, PLEP was used for the sub- sequent experiments. In addition, endotoxin inhibitor poly- myxin B (PB) was used to confirm whether the PLE and PLEP exhibits LPS-like mode action or is contained with LPS. PB was pre-treated at a concentration of 5 μg/ml in RAW 264.7 cells for 1 hr before treatment with samples (Fig.
1C). As a result, NO production was not changed after treat- ment with PB, while NO induction by LPS was reduced up to base line. This result indicates that PLE and PLEP increase NO production through different pathways from LPS.
PLEP activates macrophage RAW 264.7 cells into pro-inflammatory phenotype
Previously we found that PLEP promotes the production
of NO from RAW 264.7 cells. Production of NO is one of
the important evidence that macrophages have nature of
pro-inflammatory. Macrophages exposed to activating agents
such as LPS release a various of inflammatory cytokines and
other substances including interleukin-1β (IL-1β), IL-6, IL-8,
and TNF-α, all of which directly induce tumoricidal activity
A
B
Fig. 2. The production of mRNA and cytokine in RAW 264.7 cells stimulated with PLEP. (A) mRNA expression; (B) Cytokine production. Data are presented as mean ± SD (n=3 in each group). *p<0.05, **p<0.01, ***p<0.001 vs. CON group.
of macrophages [17]. Therefore, the effect of PLEP was inves- tigated on the release of inflammatory cytokines from RAW 264.7 cells stimulated with 10-100 μg/ml of PLEP by mon- itoring the gene expression of pro-inflammatory markers.
The mRNA expression of iNOS, which converts arginine to NO, was increased in pattern similar to results of the NO production. The gene expression of Il-1β, Il-6 and COX2 also showed the increase pattern (Fig. 2A). Cyclooxygenase (COX), known as prostaglandin-endoperoxide synthase (PTGS), is an enzyme that is responsible for the formation of instrumental biological mediators called prostanoids, including prosta- glandins, prostacyclin and thromboxane [18]. In particularly,
COX2, Il-1β, and TNF-α expressions were higher after treat-ment of 100 μg/ml of PLEP than that of 1 μg/ml of LPS as a positive control. ELISA was performed to observe the pro-inflammatory cytokines production from RAW 264.7 cells. As a result, the levels of IL-6 and TNF-α were similar to mRNA expression which was observed in Fig. 2A. The
production of IL-6 and TNF-α in the supernatant of RAW 264.7 cells stimulated with PLEP was increased in a dose-de- pendent manner (Fig. 2B). It seemed that PLEP not only reg- ulates the mRNA expression levels but also actually controls to secrete cytokines. Based on these results, it was suggested that PLEP can lead RAW 264.7 cells to tumoricidal pheno- type by producing NO and pro-inflammatory cytokines.
Also biomarkers of M2 type macrophages, Il-4, and
Arginase-1 were examined by RT-PCR. However, the ex-pression levels were too low to be detected (data not shown).
Conditioned media of RAW 264.7 cells does not affect the cell viability of two different types of pro- state cancer cells
The most generally used cell lines for prostate xenograft
models are LNCaP, PC-3, and DU145 [19, 20]. The PC-3 cell
line was originally derived from a bone metastasis of human
prostatic adenocarcinoma origin [21]. Intravenous injection
A
B
Fig. 3. Conditioned media of RAW 264.7 cells stimulated with PLEP does not affect apoptosis in prostate cancer cells. (A) Cell viability of prostate cancer cells; (B) Rate of apoptotic cell death of PC-3 cells. Various volumes of conditioned media of RAW 264.7 cells were treated into PC-3 cells and incubated for 48 hr. CON, un-treated negative control; CM, conditioned media of RAW 264.7 cells; LPS-CM, conditioned media of RAW 264.7 cells stimulated with LPS; PLEP-CM, conditioned media of RAW 264.7 cells stimulated with PLEP. Data are presented as mean ± SD (n=3 in each group). *p<0.05, ***p<0.001 vs. CON group.
of PC-3 cell line has led to the establishment of lymph node metastases [22]. The PC-3 cells have a high metastatic poten- tial and are negative for AR expression [20]. The DU145 cell line, which has weaker metastatic potential compared with the PC-3 cell line, was derived from a brain metastasis of human prostatic adenocarcinoma origin [23]. Two types of prostate cancer cells were used to observe the anti-cancer activity of conditioned media of RAW 264.7 cells stimulated with PLEP. To investigate the anti-cancer activities of PLEP itself and conditioned media of RAW 264.7 cells treated with PLEP, MTT assay was performed (Fig. 3A). As a result, PLEP itself and conditioned media of RAW 264.7 cells with LPS or PLEP did not affect the cell viability of prostate cancer cell lines. In addition, apoptosis analysis exhibited that con- ditional media of RAW 264.7 cells with stimulated with PLEP also did not affect the phenotype (Fig. 3B).
Conditioned media of RAW 264.7 cells represses the epithelial-mesenchymal transition (EMT)
To identify whether PLEP also plays a role similar to LPS, EMT-related genes such as E-cadherin, N-cadherin, snail, twist,
slug, and cadherin 11 were observed by using RT- PCR (Fig.4A). The EMT is important developmental program that is
activated during cancer metastasis and invasion. Cellular
contacts greatly limit the ability of epithelial cells to move
or migrate. It has been proposed that tumor cells lose this
restriction during the last steps of the process of tumori-
genesis, concomitantly with the loss of epithelial character-
istics. One of the proteins that is down-regulated at this stage
of tumorigenesis is E-cadherin, a molecule integral in the
establishment of homotypic adhesion junctions [24-26]. The
strong evidence that down-regulation of E-cadherin during
the process of epithelial differentiation occurs at the tran-
scriptional level [29-31]. Expression of E-cadherin was in-
A
B
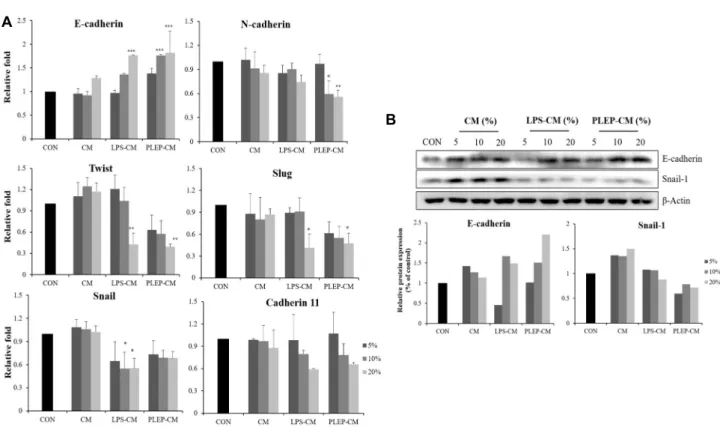
Fig. 4. Conditioned media of RAW 264.7 cells activated with PLEP suppress EMT process in PC-3 cells. (A) mRNA expression;
(B) Protein expression. The various volumes (5, 10, and 20%) of the conditioned media were treated in PC-3 cells for 48 hr. The mRNA expression levels were determined by real-time PCR and normalized to GAPDH. Data are presented as mean ± SD (n=3 in each group). *p<0.05, **p<0.01, ***p<0.001 vs. CON group.
creased in a dose-dependent manner after treatment with conditioned media of RAW 264.7 cells stimulated with PLEP or LPS which increased to 1.81-fold in PC-3 cells treated with conditioned media of RAW 264.7 cells stimulated with PLEP and 1.71-fold with LPS. On the other hand, the level of mRNA expression of N-cadherin was decreased. When the conditioned media of RAW 264.7 cells stimulated with PLEP showed 0.59-fold decrease compare to control. Loss of E-cad- herin or switch of E-cadherin to N-cadherin leads to destruc- tion of cell-cell adhesion, which drives adenoma to become carcinoma [32]. Mesenchymal markers, Snail and Twist, are able to down-regulated the level of E-cadherin and drive EMT to occur [34]. Expression of snail was decreased to 0.68-fold with conditioned media stimulated with PLEP and 0.55-fold with LPS. Expression of twist was decreased to 0.39-fold with conditioned media stimulated with PLEP and 0.42-fold with LPS. Expression of Slug was decreased to 0.47-fold with conditioned media stimulated with PLEP and 0.41-fold with LPS. Expression of cadherin 11 was decreased to 0.65-fold with conditioned media stimulated with PLEP and 0.59-fold with LPS. In the same manner, the expression of EMT-related proteins showed similar results with mRNA
expression. The protein expression of E-cadherin was in- creased in a dose-dependent manner in PC-3 cells after treat- ment of conditioned media of RAW 264.7 cells and on the other hand, snail-1 protein was decreased (Fig. 4B). Thus, conditioned media of RAW 264.7 cells activated by PLEP inhibit EMT by a mechanism that reduces N-cadherin and increases E-cadherin in PC-3 cells.
Conditioned media of RAW 264.7 cells inhibits migration, invasion, and angiogenesis ability of PC-3 cells
Cell migration and invasion are important steps in many
physiological events such as implantation of embryo, em-
bryogenesis, morphogenesis, neurogenesis, angiogenesis,
wound healing, and inflammation [35, 39]. Cell migration
and invasion are also involved in the pathophysiology of
many diseases, such as cancer. Furthermore, the capacity to
produce metastases in very different cancers is the main fea-
tures of malignant tumors, which is one of the main causes
of death for cancer [42]. There is a report that decreased
expression of migration-related genes, vimentin and CCL2,
inhibited the migration ability [43]. As a result of mRNA
A
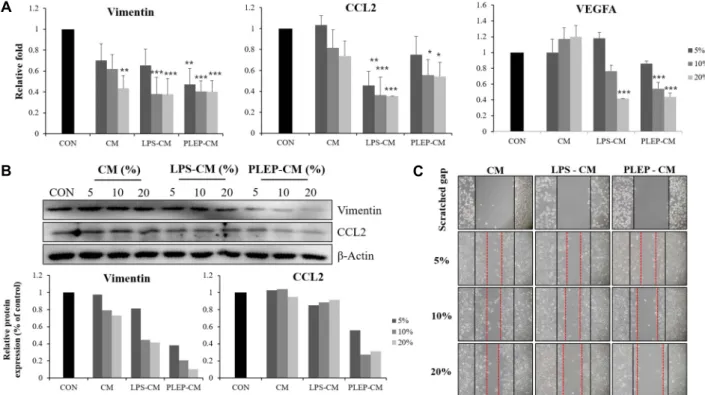
B C
Fig. 5. Conditioned media of RAW 264.7 cells represses migration, invasion and angiogenesis ability in PC-3 cells. (A) mRNA ex- pression; (B) Protein expression; (C) Wound healing. Data are presented as mean ± SD (n=3 in each group). *p<0.05, **p<0.01,
***p<0.001 vs.CON group.
expression of vimentin and ccl2 was decreased to 0.40 and 0.54-fold with conditioned media with PLPE and 0.37 and 0.36-fold with LPS, respectively (Fig. 5A). On the other hand, the protein expression of vimentin and CCL2 protein was further reduced after treatment with the conditioned media of RAW 264.7 cells stimulated with PLEP (Fig. 5B). Wound healing assay was performed to confirm the migration effi- ciency of PC-3 cells. PC-3 cells were scratched with 200 μl tips and treated with conditioned media of RAW 264.7 cells for 24 hr. Images of the wound were taken at x100 magnifi- cations using an optical microscope. Cell migration ability was more inhibited in PC-3 treated with conditioned media of RAW 264.7 activated with PLEP than LPS (Fig. 5C). These results indicate that the conditioned media of RAW 264.7 cells stimulated with PLEP inhibits the migration of PC-3 cells through regulation of vimentin and CCL2. Additional- ly, the expression of angiogenesis-related gene, vegfa, was down-regulated to 0.43-fold with conditioned media with PLEP and 0.41-fold with LPS (Fig. 5A). Angiogenesis is phys- iological process through which new blood vessels shape from pre-existing vessels [44]. Angiogenesis is an advanced field of cancer research and has been proposed as a new treatment. These results implied that conditioned media of
RAW 264.7 cells stimulated with PLEP inhibits the capability of invasion and angiogenesis through reducing the mRNA expression of vegfa.
Discussion
Prostate cancer is a first leading cause of cancer incidence in the male population, especially, in the United State. That is also extremely increasing as a fifth incidence rate in Korean male. One of the causes of high mortality in prostate cancer is the spread of tumor cells to many tissues, including lymph nodes and bones [46]. More than 70% of patients were diagnosed with bone metastases. Many studies are under- way to develop anti-cancer drugs and preventive therapies.
However, synthetic drugs already developed are known to have side effects or toxicity, depending on their use and fre- quency of use. Now ingredients that were ingested in every- day life are considered to be safe for human body because of no cytotoxicity and genotoxicity unlike conventional an- ti-cancer drugs. Many studies have been researched on the hot water extract of fruit body of Phellinus linteus as it has been known of blocking effect on digestive system cancer.
It also has an effect on the enhancement of immune function
when the chemotherapy is combined for the cancer of the gastrointestinal system including gastric cancer, esophageal cancer, colon cancer, rectal cancer, and liver cancer [14].
It has been reported that macrophages are likely to kill tumor cells if properly activated by stimulator, such as inter- feron (IFN) -γ, and lipopolysaccharides (LPS) [10]. In this study, it was proved that the P. linteus hot water extract (PLE) produces nitric oxide (NO) from macrophages RAW 264.7 cells (Fig. 1A). Since PLE is a natural product-derived extract, it contains biological compounds showing both ac- tivities to increase NO production and decrease NO (Fig.
1B). To solve this problem, a simple purification was carried out using Diaion HP-20. As a result, molecules (PLEP) capa- ble of activating NO production, and molecules (PLEE) in- hibiting NO production could be separated (Fig. 1B). PLEP that activates NO production was confirmed by using re- al-time PCR and ELISA (Fig. 2). To determine the anti-cancer effect of conditioned media of RAW 264.7 cells activated by PLEP, we performed MTT and Annexin V-PI double staining assay. As a result, anti-cancer activity of conditioned media of RAW 264.7 cells stimulated with PLEP was not observed (Fig. 3).
EMT is a very important step in metastasis, during which non-motile, polarized epithelial cells separate their cell junc- tions and change into individual, motile mesenchymal cells [27, 28, 33]. Conditioned media of RAW 264.7 cells stimu- lated with PLEP was suppressed EMT by regulating EMT biomarkers in human prostate cancer PC-3 cells. It was also investigated the effect of conditioned media on migration and invasion by RT-PCR, western blotting, and wound heal- ing analysis. As a result, vimentin, CCL2, and VEGFA, were down-regulated and the wound healing was also inhibited by the conditioned media of RAW 264.7 cells stimulated with PLEP (Fig. 5). However, the mRNA level of vimentin and ccl2 was not matched with protein levels. This incon- sistency of mRNA and protein correlation has been observed with several reasons [36, 37, 40]. For one example, in the NIH3T3 cells, some genes, which is related with house- keeping genes, have stable mRNAs and proteins, thus mRNAs and proteins tended to have good correlation.
However, specific genes related with transcription factors, genes with cell cycling and coding genes with signaling are unstable and protein is also same as well [36]. Therefore, this inconsistence of mRNA and protein levels from vi- mentin and CCL2 can occur.
In conclusion, it was suggested that PLEP induces pro-in-
flammatory effect of RAW 264.7 cells and its conditioned media has an effect on migration and invasion by inhibiting EMT of PC-3 prostate cancer cells. Although the conditioned media of RAW 264.7 macrophages does not directly kill can- cer cells, its anti-cancer effect can be expected through in- hibiting migration and invasion of cancer cells. Finally, this new direction will be helpful for cancer therapy after in vivo animal experiments for more accurate treatment.
References
1. Banyard, J., Chung, I., Migliozzi, M., Phan, D. T., Wilson, A. M., Zetter, B. R. and Bielenberg, D. R. 2014. Identification of genes regulating migration and invasion using a new model of metastatic prostate cancer. BMC Cancer 14, 387- 402.
2. Berthod, F. 2013. Fibroblasts and endothelial cells: the basic angiogenic unit. Angiogenesis: insights from a systematic over- view, pp. 145-157, Nova Science Publishers, NY, USA.
3. Bolós, V., Peinado, H., Pérez-Moreno, M. A., Fraga, M. F., Esteller, M. and Cano, A. 2003. The transcription factor Slug represses E-cadherin expression and induces epithelial to mesenchymal transitions: a comparison with Snail and E47 repressors. J. Cell Sci. 116, 499-511.
4. Bowen, P. E. 2004. Effects of tomatoes and lycopene on prostate cancer prevention and treatment. J. Kor. Soc. Food Sci. Nutr. 33, 455-462.
5. Bozzuto, G., Ruggieri, P. and Molinari, A. 2010. Molecular aspects of tumor cell migration and invasion. Ann. Ist. Super.
Sanita 46, 66-80.
6. Cano, A., Pérez-Moreno, M. A., Rodrigo, I., Locascio, A., Blanco, M. J., del Barrio, M. G., Portillo, F. and Nieto, M.
A. 2000. The transcription factor Snail controls epithelial–
mesenchymal transitions by repressing E-cadherin expre- ssion. Nat. Rev. Mol. Cell Biol. 2, 76-83.
7. Catalona, W. J., Richie, J. P., Ahmann, F. R., Hudson, M.
A., Scardino, P. T., Flanigan, R. C., DeKernion, J. B., Ratliff, T. L., Kavoussi, L. R., Dalkin, B. L., Waters, W. B., MacFarlane, M. T. and Southwick, P. C. 2017. Comparison of digital rectal examination and serum prostate specific antigen in the early detection of prostate cancer: results of a multicenter clinical trial of 6,630 men. Int. J. Urol. 197, S200-S207.
8. Cho, S. Y. and Klemke, R. L. 2000. Extracellular-regulated kinase activation and Cas/Crk coupling regulate cell migra- tion and suppress apoptosis during invasion of the ex- tracellular matrix. J. Cell Biol. 149, 223-236.
9. Choi, J. H., Kim, H. I. and Lee, I. S. 2009. Effect of rosmar- inus officinalis L. on growth inhibition and apoptosis in- duction in cancer cells. J. Kor. Soc. Food Sci. Nutr. 38, 1008- 1015.
10. Dhingra, A. K., Chopra, B., Dass, R. and Sanjeev, M. S.
K. 2014. A review on COX and their inhibitors: Present and future. In. Pharm. Pharmacol. 2, 470-485.
11. Elloul, S., Elstrand, M. B., Nesland, J. M., Tropé, C. G., Kvalheim, G., Goldberg, I., Reich, R. and Davidson, B. and Davidson, B. 2005. Snail, Slug, and Smad-interacting protein 1 as novel parameters of disease aggressiveness in meta- static ovarian and breast carcinoma. Cancer 103, 1631-1643.
12. Gómez-Acebo, I., Dierssen-Sotos, T., Fernandez-Navarro, P., Palazuelos, C., Moreno, V., Aragonés, N., Castaño-Vinyals, G., Jiménez-Monleón, J. J., Ruiz-Cerdá, J. L., Pérez-Gómez, B., Ruiz-Dominguez, J. M., Molero, J. A., Pollán, M., Koge- vinas, M. and Llorca, J. 2017. Risk Model for prostate cancer using environmental and genetic factors in the spanish mul- ti-case-control (MCC) study. Sci. Rep. 7, 8994.
13. Holzapfel, B. M., Wagner, F., Loessner, D., Holzapfel, N.
P., Thibaudeau, L., Crawford, R., Ling, M. T., Clements, J. A., Russell, P. J. and Hutmacher, D. W. 2014. Species-spe- cific homing mechanisms of human prostate cancer meta- stasis in tissue engineered bone. Biomaterials 35, 4108-4115.
14. Huttenlocher, A. and Horwitz, A. R. 2011. Integrins in Cell Migration. Cold Spring Harb. Perspect Biol. 3, a005074.
15. Imtiaj, A., Sultana, S., Hossain, J., Rahman, S. A. and Ohga, S. 2016. Performance of vegetative growth and artificial fruit body formation of hypsizygus marmoreus in Bangladesh.
J. Fac. Agr. Kyushu U. 62, 258-262.
16. Ji, J. H., Kim, M. N., Chung, C. K., and Ham, S. S. 2000.
Antimutagenic and cytotoxity effects of Phellinus linteus Extracts. J. Kor. Soc. Food Sci. Nutr. 29, 322-328.
17. Ji, Z., Tang, Q., Zhang, J., Yang, Y., Jia, W. and Pan, Y.
2007. Immunomodulation of RAW264.7 macrophages by GLIS, a proteopolysaccharide from Ganoderma lucidum. J.
Ethnopharmacol. 112, 445-450.
18. Kalluri, R. and Neilson, E. G. 2003. Epithelial-mesenchymal transition and its implications for fibrosis. J. Clin. Invest.
112, 1776-1784.
19. Kang, H. I., Kim, J. Y., Cho, H. D., Park, K. W., Kang, J. S. and Seo, K. I. 2010. Resveratrol induces apoptosis in primary human prostate cancer cells. J. Kor. Soc. Food Sci.
Nutr. 39, 1119-1125.
20. Lee, J. S., Shin, D. B., Lee, S. M., Kim, S. H., Lee, T. S.
and Jung, D. C. 2013. Melanogenesis inhibitory and anti- oxidant activities of Phellinus baumii methanol extract. The Kor. J. Myc. 41, 104-111.
21. Lee, K. H., Kwon, H. J., Chun, S. S., Kim, J. H., Cho, Y.
J. and Cha, W. S. 2006. Biological activities of extracts from Phellinus linteus. Appl. Biol. Chem. 49, 298-303.
22. Li, P., Yang, R. and Gao, W. Q. 2014. Contributions of epi- thelial-mesenchymal transition and cancer stem cells to the development of castration resistance of prostate cancer. Mol.
Cancer 13, 55.
23. Müller, E., Christopoulos, P. F., Halder, S., Lunde, A., Beraki, K., Speth, M., Øynebråten, I. and Corthay, A. 2017.
Toll-like receptor ligands and interferon-γ synergize for in- duction of antitumor M1 macrophages. Front. Immunol. 8, 1383.
24. Rembold, M., Ciglar, L., Yáñez-Cuna, J. O., Zinzen, R. P., Girardot, C., Jain, A., Welte, M. A., Stark, A., Leptin, M.
and Furlong, E. E. 2014. A conserved role for Snail as a
potentiator of active transcription. Genes Dev. 28, 167-181.
25. Rodriguez, L. G., Wu, X. and Guan, J. L. Wound-healing assay. 2005. Methods Mol. Biol. 294, 23-29.
26. Rucci, N. and Angelucci, A. 2014. Prostate cancer and bone:
the elective affinities. Biomed Res. Int. 2014, 14-27.
27. Schwanhäusser, B., Busse, D., Li, N., Dittmar, G., Schuch- hardt, J., Wolf, J., Chen, W. and Selbach, M. 2011. Global quantification of mammalian gene expression control. Nature 473, 337-342.
28. Schwanhäusser, B., Busse, D., Li, N., Dittmar, G., Schuch- hardt, J., Wolf, J., Chen, W. and Selbach, M. 2013. Corrigen- dum: Global quantification of mammalian gene expression control. Nature 495, 126-127.
29. Singh, A. and Settleman, J. 2010. EMT, cancer stem cells and drug resistance: an emerging axis of evil in the war on cancer. Oncogene 29, 4741-4751.
30. Taichman, R. S., Cooper, C., Keller, E. T., Pienta, K. J., Taichman, N. S. and McCauley, L. K. 2002. Use of the stro- mal cell-derived Factor-1/CXCR4 pathway in prostate can- cer metastasis to bone. Cancer Res. 62, 1832-1837.
31. Tan, E. J., Kahata, K., Idås, O., Thuault, S., Heldin, C. H.
and Moustakas, A. 2015. The high mobility group A2 pro- tein epigenetically silences the Cdh1 gene during epithelial- to-mesenchymal transition. Nucleic Acids Res. 43, 162-178.
32. Thiery, J. P. 2002. Epithelial–mesenchymal transitions in tumour progression. Nat. Rev. Cancer 2, 442-452.
33. Thiery, J. P. and Sleeman, J. P. 2006. Complex networks orchestrate epithelial–mesenchymal transitions. Nat. Rev.
Mol. Cell Biol. 7, 131-142.
34. Thiery, J. P., Acloque, H., Huang, R. Y. J. and Nieto, M.
A. 2009. Epithelial-mesenchymal transitions in development and disease. Cell 139, 871-890.
35. Tian, X. M., Yu, H. Y., Zhou, L. W., Decock, C., Vlasák, J. and Dai, Y. C. 2013. Phylogeny and taxonomy of the Inonotus linteus complex. Fungal Divers. 58, 159-169.
36. Van der Pluijm, G. 2011. Epithelial plasticity, cancer stem cells and bone metastasis formation. Bone 48, 37-43.
37. Voulgari, A. and Pintzas, A. 2009. Epithelial–mesenchymal transition in cancer metastasis: Mechanisms, markers and strategies to overcome drug resistance in the clinic. Biochim.
Biophys. Acta 1796, 75-90.
38. Wang, T. T. Y., Schoene, N. W., Kim, Y. S., Mizuno, C.
S. and Rimando, A. M. 2010. Differential effects of resvera- trol and its naturally occurring methylether analogs on cell cycle and apoptosis in human androgen-responsive LNCaP cancer cells. Mol. Nutr. Food Res. 54, 335-344.
39. Watson, P. A., Arora, V. K. and Sawyers, C. L. 2015. Emerg- ing mechanisms of resistance to androgen receptor inhibi- tors in prostate cancer. Nat. Rev. Cancer 15, 701-711.
40. Wilhelm, M., Schlegl, J., Hahne, H., Gholami, A. M., Lieber- enz, M., Savitski, M. M., Ziegler, E., Butzmann, L., Gessulat, S., Marx, H., Mathieson, T., Lemeer, S., Schnatbaum, K., Reimer, U., Wenschuh, H., Mollenhauer, M., Slotta-Huspe- nina, J., Boese, J. H., Bantscheff, M., Gerstmair, A., Faerber, F. and Kuster, B. 2014. Mass-spectrometry-based draft of the human proteome. Nature 509, 582-587.
초록:상황버섯에 의해 활성화된 RAW 264.7 대식세포주 배양액의 인간 전립선암 세포주의 epithelial- mesenchymal transition 조절
강태우
1,2,3†․안현희
4†․박슬기
4․유선녕
4․황유림
4․김지원
4․안순철
4*
(1부산대학교병원, 2부산지역암센터(외과), 3의생명연구원, 4부산대학교 의학전문대학원 미생물학 및 면역학 교실)
전립선암은 전이성 종양 중의 하나로 치료를 위해 호르몬 요법이나 외과 적 거세 방법이 주로 수행되지만 많은 부작용을 나타내었다. 최근 많은 연구자들이 이러한 상황을 해결하기 위해 종양 미세 환경을 연구하고 있으며 그 중 면역 세포, 특히 대식세포는 종양 미세 환경의 중요한 구성요소이다. 정상적인 조건에서 대식세포는 여러 암세포에 대해 약한 종양 살균 활성을 갖으나 interferon-γ 또는 lipopolysaccharide에 의해 활성화되면, 염증성 사이토카인 및 케모카인을 분비함으로써 암세포를 직접 또는 간접적으로 사멸 시키게 된다. 본 연구에서는, 마우 스 대식세포인 RAW 264.7 세포에 Phellinus linteus 추출물을 처리하여 산화질소의 방출과 pro-inflammatory cyto- kine들을 real-time PCR과 ELISA 방법으로 분석하였다. RAW 264.7의 조정 배지는 48시간 동안 전립선 암세포처 리하여 상피간엽세포전이 관련 유전자의 발현을 측정 하였다. 그 때에 mesenchymal 관련 유전자들인 N-cadherin,
snail, twist, slug 및 cadherin 11이 감소했을 뿐만 아니라 epithelial 관련 유전자인 E-cadherin은 증가하였다. 또한암 전이 및 신생 혈관 형성에 관여하는 vimentin, ccl2 및 vegfa가 감소되었는데, 이는 EMT가 암세포의 이동과 침범 에 밀접한 관련이 있기 때문이다. 따라서 Phellinus linteu에 의해 자극된 RAW 264.7 세포의 조정 배지는 인간 전립 선 암세포주인 PC-3 세포의 이동과 전이를 억제하고 EMT 경로를 조절한다는 것을 나타낸다.
41. Wiseman, M. 2008. The second world cancer research fund/
american institute for cancer research expert report. food, nutrition, physical activity, and the prevention of cancer:
a global perspective: nutrition society and bapen medical symposium on ‘nutrition support in cancer therapy’. Proc.
Nutr. Soc. 67, 253-256.
42. Wu, X., Gong, S., Roy-Burman, P., Lee, P. and Culig, Z.
2013. Current mouse and cell models in prostate cancer research. Endocr. Relat. Cancer 20, R155-R170.
43. Yang, C. Y., Lin, C. K., Lin, G. J., Hsieh, C. C., Huang, S. H., Ma, K. H., Shieh, Y. S., Sytwu, H. K. and Chen, Y.
W. 2017. Triptolide represses oral cancer cell proliferation, invasion, migration, and angiogenesis in co-inoculation
with U937 cells. Clin. Oral Investig. 21, 419-427.
44. Yang, J. and Weinberg, R. A. 2008. Epithelial-mesenchymal transition: at the crossroads of development and tumor metastasis. Dev. Cell 14, 818-829.
45. Yoon, J. A., Hahm, S. W., Park, J. E. and Son, Y. S. 2009.
Total polyphenol and flavonoid of fruit extract of Opuntia humifusa and its inhibitory effect on the growth of MCF-7 human breast cancer cells. J. Kor. Soc. Food Sci. Nutr. 38, 1679-1684.
46. Zeisberg, M. and Neilson, E. G. 2009. Biomarkers for epi- thelial-mesenchymal transitions. J. Clin. Invest. 119, 1429- 1437.