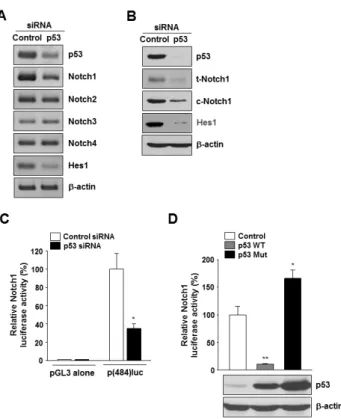
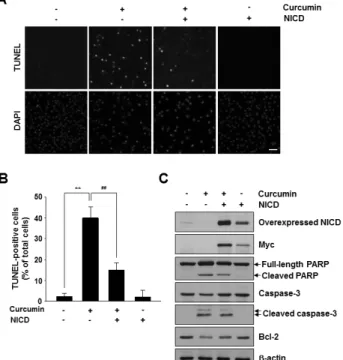
291 http://dx.doi.org/10.4196/kjpp.2013.17.4.291
ABBREVIATIONS: ChIP, chromatin immunoprecipitation; EMSA, electrophoretic mobility shift assay; NICD, Notch1 intracellular domain; siRNA, small interfering RNA; TUNEL, terminal deoxy- nucleotidyl transferase dUTP nick end labeling; RT-PCR, reverse transcription polymerase chain reaction.
Received February 26, 2013, Revised May 15, 2013, Accepted May 28, 2013
Corresponding to: Soo-Kyung Bae, Department of Dental Pharma- cology, School of Dentistry, Yangsan Campus of Pusan National University, 49 Busandaehak-ro, Moolgeum-eup, Yangsan 626-870, Korea. (Tel) 82-51-510-8253, (Fax) 82-51-510-8233, (E-mail) skbae@
pusan.ac.kr
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://
creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.