Lab Anim Res 2017: 33(3), 202-208 https://doi.org/10.5625/lar.2017.33.3.202
ISSN 2233-7660 (Online)
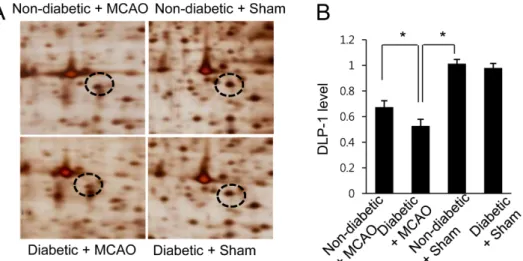
Hyperglycemia exacerbates downregulation of dynamin-like protein 1 in ischemic cerebral injury
Dong-Ju Park
1, Myeong-Ok Kim
2, Phil-Ok Koh
1,*
1
Department of Anatomy, College of Veterinary Medicine, Research Institute of Life Science,
2