J F E S
Journal of Forest and Environmental Science
Journal of Forest and Environmental Science Vol. 34, No. 4, pp. 338-351, August, 2018 https://doi.org/10.7747/JFES.2018.34.4.338
Managing Red Oak ( Quercus rubra L.) Reduces Sensitivity to Climatic Stress
Sophan Chhin*
Division of Forestry and Natural Resources, West Virginia University, Morgantown, WV 26506, USA
Abstract
This study was conducted in a long-term experimental forest in the central hardwoods region of southwestern Michigan to retrospectively examine the role of past forest management practices and climate on red oak (Quercus rubra L.) productivity. Initially, in 1971, plots within the experimental forest were treated separately with a clearcut and shelterwood regeneration harvest in an attempt to increase red oak regeneration. From 1987-1989, a new study was initiated within a portion of the clearcut and shelterwood plots to evaluate the effectiveness of additional oak crop tree release using mechanical and chemical applications. Cumulative diameter and mortality rates of 719 red oaks were monitored across the four silvicultural treatments: Clearcut-A (clearcut without additional release treatment), Clearcut-B (clearcut with additional release treatment), Shelterwood-A (shelterwood without additional release), and Shelterwood-B (shelterwood with additional release) plus an untreated control. Increment cores were obtained from red oak trees and neighboring competitor species. Tree-ring analyses (dendrochronology) were applied to examine the effect of these silvicultural treatments and climatic factors (temperature and precipitation) on red oak productivity. The results indicated that crop tree release following a clearcut or shelterwood harvest reduced mortality rates and thus increased survival of red oak. Red oak in control plots or plots only receiving the initial regeneration harvesting treatment and no additional competition release were negatively affected by climatic stress, which included summer moisture stress. In contrast, red oak in plots that received the competition release treatment from shade tolerant tree species not only had higher tree level productivity (i.e., tree basal area) and lower mortality rates, but were also relatively more resilient to climatic stress by showing limited or no associations between climate and growth.
Key Words: clearcut, climate, dendrochronology, release treatment, shelterwood
Received: December 8, 2017. Revised: March 17, 2018. Accepted: April 26, 2018.
Corresponding author: Sophan Chhin
Division of Forestry and Natural Resources, West Virginia University, 322 Percival Hall, PO Box 6125, Morgantown, WV, 26506, USA Tel: 1-304-293-5313, Fax: 1-304-293-2441, E-mail: [email protected]
Introduction
Future climate change may exacerbate existing chal- lenges with regenerating oak species and maintaining their dominance over shade tolerant competitors (Johnson et al.
2002). There are multiple avenues toward increasing adap- tive capacity of forests to climate change which includes fos- tering forest resilience or promoting forest resistance (Millar et al. 2007). Ecosystem resilience is defined as the
capacity of an ecosystem to absorb some disturbance (e.g., drought), which in turn may induce some minor degree of ecosystem change, but still maintain its essential structure and function (Folke et al. 2004; DeRose and Long 2014).
Resilient forests can undergo gradual structural and func- tional changes due to climatic stress but also have the ca- pacity to eventually recover to the predisturbance condition (Millar et al. 2007; D’Amato et al. 2013). Forest resistance is defined as the capacity of forests to resist the influence of
climate change while generally maintaining structural and functional integrity during the climatic disturbance event (Millar et al. 2007). Forest management practices (e.g., in- termediate stand tending practices such as thinning) can moderate the degree of competition for environmental re- sources (i.e., soil moisture, light) which in turn has the po- tential to buffer residual trees from stressful climatic con- ditions (Laurent et al. 2003, Millar et al. 2007, Chmura et al. 2011, D’Amato et al. 2013, Magruder et al. 2013; Dolos et al. 2016). This resource buffering effect in turn may de- couple growth from the prevailing regional climate. For in- stance, planting trees at low initial stand densities (Magruder et. al. 2012) and thinning the smaller crown classes to a moderate residual density (Magruder et al.
2013) can increase resiliency of red pine (Pinus resinosa Ait.) to climatic stress. In addition to the general tolerance of oak to drier sites, silvicultural preference of oak in some regions such as Europe is expected to lead to replacement of oak over less drought-adapted species (Dolos et al. 2016).
The instrumental climatic record has indicated that global average surface temperatures have increased by 0.78°C for the 2003-2012 period relative to 1850-1900 pe- riod (IPCC 2013). Projections of future climate change based on general circulation models and different emission scenarios of greenhouse gases indicate a further warming within the range of 0.3°C–4.8°C by the end of the 21st cen- tury (2081-2100) relative to 1986-2005 (IPCC 2013).
While little change is expected in annual average precip- itation, higher temperatures are expected to lead to in- creased rates of evapotranspiration in plants. Future climate change (i.e., global warming and increased summer dry- ness) is expected to generally reduce forest productivity and increase rates of tree mortality (Allen et al. 2010). Regional climate forecasts for the state of Michigan and the Great Lakes Region also indicate that average temperatures will rise 3-11°C in the summer and 3-7°C in the winter by the end of the 21st century (Kling et al. 2003; Handler et al.
2014). The projected regional increase in temperature is ex- pected to shorten winters and decrease the duration of lake ice cover (Kling et al. 2003). Furthermore, climatic stress can lower tree productivity and weaken tree health and thereby increase the vulnerability of tree hosts to dis- turbance agents such as insect pests (Finley et al. 2016) and fungal pathogens (Handler et al. 2014; Chhin and O’Brien
2015).
In dry-mesic closed-canopy forest communities of the central hardwood forest regions, oak species such as red oak are difficult to regenerate and maintain in the co-dominant and dominant crown classes (Johnson et al. 2002). In the absence of disturbances such as fire, oak dominated forests succeed to shade tolerant competitors including sugar ma- ple (Acer saccharum Marsh.), red maple (Acer rubrum L.), and other mesophytic hardwoods (e.g., black cherry (Prunus serotina Enrh.) especially on higher quality mesic sites (Brose et al. 2013; Dey 2014). The poor competitive ability of oak on mesic sites has been attributed to the adap- tation of oak species to concentrate growth in the juvenile years towards root production at the expense of above- ground shoot production (Johnson et al. 2002). This growth allocation pattern is an adaptation towards having the resources for resprouting after damage to the above- ground shoot from disturbances such as fire and herbivore grazing. Regeneration of oak in more mesic closed canopy forests can be promoted with regeneration harvest treat- ments (i.e., clearcut and shelterwood) (Hill and Dickmann 1988; Crunkilton et al. 1992; Buckley et al. 1998; Dech et al. 2008; Parker and Dey 2008; Dey et al. 2009; Spetich et al. 2009; Dey 2014) and prescribed fires fires (Brose et al.
2013). Since natural regeneration may be insufficient, un- derplanting has been promoted to increase oak stocking levels (Cogliastro and Paquette 2012; Parrott et al. 2012;
Lhotka and Loewenstein 2013; Craig et al. 2014). To in- crease oak survival and growth and attaining a stable posi- tion in the co-dominant and dominant crown classes, crop tree release around selected oak trees has been advocated and generally involves timely midstory removal of compet- ing tree species (Gauthier and Jacobs 2010; Cogliastro and Paquette 2012; Parrott et al. 2012; Lhotka 2013; Lhotka and Loewenstein 2013; Craig et al. 2014; Dey 2014).
Red oak (Quercus rubra L.) is an intermediate shade tol- erant tree species (Sander 1990), which in the absence of disturbances (i.e., fire) is succeeded by more shade tolerant competitors (Kabrick et al. 2008). Red oak xylem has a ring porous structure containing large early wood conducting vessels produced using energy reserves generated in the previous growing season (LeBlanc and Stahle 2015). In southern Michigan, site quality for red oak is affected by moisture availability and dependent on the depth to ground
Fig. 1. (a) Location of Fred Russ Experimental Forest (●) managed by Michigan State University in Cass County and regional context of the state of Michigan in the midwestern United States. (b) Physical layout of silvi- cultural experimental design showing 4 experimental blocks.
CC-A=Clearcut A (clearcut without additional release treatment);
CC-B=Clearcut-B (clearcut with additional release treatment);
SW-A=Shelterwood-A (shelterwood without additional release);
SW-B=Shelterwood-B (shelterwood with additional release).
water table (Sander 1990). Oak acorns provide an im- portant resource for many seed predators, which in eastern deciduous forests include mice, squirrels, and deer (Sander 1990; Johnson et al. 2002). Previous dendrochronological studies of red oak have shown that drought negatively im- pacts growth (Pan et al. 1997; Abrams et al. 1998; LeBlanc 1998; Fekedulegn et al. 2003; Speer et al. 2009; LeBlanc and Terrell 2011), and that drought can increase suscepti- bility to other disturbances such as acid rain and insects (LeBlanc 1998). Red oak studies have also indicated that drought primarily controls distributional range limits (Tardif and Conciatori 2006; Tardif et al. 2006). Exotic non-native populations of red oak planted in Europe have further underscored the importance of summer moisture stress as a limiting factor to radial growth (Matisons et al.
2015). Few studies have examined how silvicultural treat- ments can modify sensitivity of red oak to climatic stress (Rentch et al. 2002).
The objective of this study is to examine the impact of sil- vicultural management practices (including shelterwood and clearcut regeneration harvests followed by oak crop tree release treatments) on: a) red oak diameter growth and mortality; and b) sensitivity to climatic stress. It is hypothe- sized that red oak receiving multiple silvicultural inputs (i.e., regeneration harvests plus competition release treat- ments) will show higher diameter growth and reduced mor- tality rates since residual trees will have greater growing space and access to environmental resources (Laurent et al.
2003; Millar et al. 2007; Chmura et al. 2011; D’Amato et al. 2013; Magruder et al. 2013). Furthermore, it is ex- pected silvicultural investments will likely decouple the re- lationship between radial growth of red oak and the region- al climate since the increased growing space of residual trees will likely increase resource availability and moderate the effects of climatic stress (Magruder et al. 2013).
Materials and Methods
Study Site and Field Sampling Design
This study was conducted in the Michigan State University (MSU) Fred Russ Experimental Forest which is located in Cass County in southwest Michigan (Fig. 1a).
This experimental forest is located in the central hardwoods region of southern Michigan (Parker and Merritt 1994)
and in the unmanaged areas, including designated control plots, is dominated by an overstory of red oak, white oak (Quercus alba L.), and black oak (Quercus velutina Lam.).
According to the nearest meteorological station (Three Rivers, Station ID# 208184, 28 km southeast of Fred Russ) and for the 30-yr reference period of 1980–2009, mean annual temperature was 9.4°C with warmest mean monthly temperatures in July (22.2°C) and coldest mean monthly temperatures occurring in January (-4.3°C). Total annual precipitation was 986 mm with most precipitation
occurring in July (109 mm). The soils are classified as deep, well-drained sandy loams belonging to the Oshtemo (Coarse-loamy, mixed, active, mesic Typic Hapludalfs) and Kalamazoo (Fine-loamy, mixed, active, mesic Typic Hapludalfs) series originating from outwash plains (Hill and Dickmann 1988). The 50-year site index for red oak on this site is 20.1 m (Hill and Dickmann 1988).
The stand management history consisted of a control and three types of regeneration harvests (shelterwood, clearcut, and group selection) applied in the fall of 1971 (Hill and Dickmann 1988). The original 18.2 ha stand had uniform stand characteristics and was divided into 4 blocks, and each block consisted of 4 plots to which the 4 silvicultural treatment types were randomly assigned. Each treatment plot had a size of 1.11 ha with plot interiors (0.71 ha) used for forest inventory and measurements. The control treat- ment had no cutting. All trees were removed in the clearcut treatment. The group selection cut involved removing a group of trees occupying an area of 0.08 ha. The shelter- wood treatment consisted of an establishment cut that left behind a residual basal area range of 4.82 m2/ha to 8.03 m2/ha. The residual overstory in the shelterwood treatment was removed during the winter of 1974-1975.
In 1987, a second set of treatments (including a control and red oak crop tree release treatment) was applied in the shelterwood and clearcut plots (Fig. 1b). Each shelterwood and clearcut plot (0.71 ha) was equally divided into two sub-plots (0.35 ha). Each sub-plot was randomly assigned either a control (designated Clearcut-A [CC-A] or Shelterwood-A [SW-A]) treatment or a red oak crop tree release treatment (designated Clearcut-B [CC-B] or Shelterwood-B [SW-B]). Unfortunately, one of the treat- ment plots (in experimental block #1) for the shelter- wood treatment was damaged, reducing the number of replicates down from 4 to 3 for this treatment type. This led to an incomplete randomized split-plot experimental design. The control sub-plots were left untreated and ap- proximately 20-30 trees/ha were selected and tagged for future monitoring. In the sub-plots receiving the release treatment, approximately 20-30 trees/ha were selected for release and tagged for future monitoring. For either con- trol trees or crop trees released from competition, the cri- teria for selection included form (i.e., selecting trees with straight stems with few forks), crown class (i.e., selecting
mainly dominants, codominants and few intermediate trees), and vigor (i.e., selecting trees with healthy live crown with no dead tops). All trees surrounding a crop tree that were touching or had overlapping crowns with a crop tree were removed with either chainsaw or herbicide application. The herbicide application involved low-vol- ume basal bark spraying with a mixture of 75% basil oil mixed with 25% garlon-4 (obtained from ArborChem, Mechanicsburg, PA). Since the chemical release in 1987 was not uniformly effective, in 1989, surviving com- petitor trees in the competitive zone of red oak crop trees were removed with a chainsaw to complete the release treatment.
The current study is focused on these 5 treatment types: unmanaged control that has had no silvicultural practices applied, Clearcut-A (control), Clearcut-B (release), Shelterwood-A (control), and Shelterwood-B (release) (Fig. 1b). Mortality and diameter at breast height (DBH) were recorded for all red oak trees that were tagged in these 4 treatment types and the surveys were conducted in 1992, 1998, 2003, and 2015 and com- pared to a baseline pretreatment survey conducted in 1986. Each treatment plot was divided into 4 equal sized quadrants, followed by random selection of 3 of these quadrants, and then sampling the red oak tree nearest to the center of a selected quadrant with an increment borer by taking 2 cores per oak tree at breast height (1.3 m).
Relative to each selected oak tree, the nearest non-oak competitor species was also cored (2 cores per tree).
Nearest neighbors were all of sufficient size to have some degree of crown interaction with focal red oak trees such that a minimum DBH threshold was not required. For the unmanaged control, Clearcut-A and Clearcut-B, 12 red oak and 12 competitor trees were sampled in each of these treatment categories. For Shelterwood-A and Shelterwood-B, 9 red oak and 9 competitor trees were sampled in these treatment categories due to the loss of one replicate block. Stand basal area was assessed in summer of 2011 using a wedge prism (i.e., 10 BAF) at the center of each of the 3 randomly chosen quadrants of each treatment type.
Sample Processing, Cross-Dating and Tree-Ring Measurement
All wood increment cores were processed according to standard dendrochronological techniques and sanded with progressively finer grades of sandpaper to highlight annual rings (Stokes and Smiley 1996; Speer 2010). All samples within each treatment type were visually crossdated under a binocular microscope to identify any missing and/or false double rings (Yamaguchi 1991). All samples were digitally scanned at an optical resolution of 1,200 dpi. Annual ring width was measured using an image analysis soft- ware-based system (CooRecorder and CDendro: Cybis Elektronik and Data AB, Sweden). Difficult sections of some increment cores containing extremely narrow rings were measured with a stage micrometer coupled with a ster- eo microscope to an accuracy of 0.001 mm (Velmex:
Bloomfield, New York). Additional statistical quality con- trol was provided through the use of the program COFECHA (Holmes 1983; Grissino-Mayer 2001).
Annual ring measurements were converted to annual basal area increment values (cm2 year-1). At the end of each growth year, annual ring width measurements from each in- crement core of each tree were converted to cumulative measures of diameter inside bark (DIB). If coring missed the pith, the missing radius was estimated from the differ- ence of half of the diameter outside bark (DOB) taken in the field and total cumulative radial measurements plus bark retained in each core (Metsaranta and Lieffers 2009).
Cumulative basal area values were derived from cumulative DIB values and the formula for the area of a circle. A per- fect cylindrical shape with a central pith is assumed al- though trees can have off-center piths and elliptical shapes.
Annual basal area increment values (cm2 year-1) were ob- tained by subtracting cumulative basal area in the previous year (t-1) from the current year (t).
Analysis of Stand Monitoring Data
The mortality census data that were collected in 1992, 1998, 2003, 2015 were expressed in cumulative percent val- ues relative to the number of initial red oak individuals sur- veyed in the baseline pretreatment survey conducted in 1986. Annual mortality rate (%) between each measure- ment period was calculated according to Sheil and May
(1996): m=(1-(Nt/N0)1/t)100, where No and Nt are the population numbers at the beginning and end of the meas- urement interval (t). Cumulative diameter at breast height (DBH) and mortality rates for each inventory year were compared between the silvicultural treatments using a one-way ANOVA followed by multiple comparison tests using Fisher’s Honest Significant Difference (HSD) test in Systat (version 10.2). Except for the control treatment, the other treatment types represented a combination of both a regeneration harvest technique and whether competition release was carried out. Consequently, two-way ANOVA analyses were conducted in Systat (version 10.2) for each of the four inventory periods for cumulative diameter growth, cumulative mortality, and annual mortality to verify if inter- action occurred between regeneration harvest technique (2 levels) and whether competition release was performed (2 levels). The inventory of stand basal area was also com- pared across silvicultural treatments using a one-way ANOVA followed by Fisher’s HSD test to conduct the multiple comparisons. The significance threshold for all statistical tests was =0.05.
Growth-climate analyses
Monthly climate data was obtained from the nearest me- teorological station in Three Rivers (Station ID# 208184).
The primary climate variables provided included mean monthly temperature and total monthly precipitation.
These primary variables were used as the basis to produce a synthetic monthly climatic moisture index (CMI) calcu- lated as precipitation minus potential evapotranspiration (PET) (Hogg 1997). Monthly values of PET are primar- ily a function of mean vapor pressure deficit which in turn is estimated from monthly temperature. Consequently, CMI combines the effects of both precipitation and temperature on soil moisture regimes. Since tree growth may respond more strongly with climatic variables at the seasonal scale, monthly climatic variables were converted to seasonal 2-month and 3-month periods (temperature variables aver- aged during the 2-month or 3-month seasonal periods, and moisture index variables were summed during the seasonal periods) over two growing seasons from April of the prior year (t-1) to October of the current year (t) or radial growth (Chhin et al. 2008).
Interannual basal area increment time series for each in-
Table 1. Species composition (percentage followed by number of trees) of the nearest competitor species of red oak across different silivicul- tural treatments
Species Control (%) Clearcut-A (%) Clearcut-B (%) Shelterwood-A (%)
Shelterwood-B
(%) Overall (%)
Acer rubrum 20.0 (2) 0 (0) 0 (0) 0 (0) 0 (0) 3.8 (19)
Acer saccharum 40.0 (4) 0 (0) 8.3 (1) 0 (0) 22.2 (2) 13.5 (8)
Carya glabra 30.0 (3) 8.3 (1) 8.3 (1) 0 (0) 0 (0) 9.6 (8)
Fraxinus americana 0 (0) 0 (0) 0 (0) 0 (0) 11.1 (1) 1.9 (7)
Fagus grandifolia 10.0 (1) 0 (0) 0 (0) 0 (0) 0 (0) 1.9 (5)
Juglans nigra 0 (0) 0 (0) 0 (0) 0 (0) 11.1 (1) 1.9 (2)
Populus grandidenta 0 (0) 25.0 (3) 16.7 (2) 22.2 (2) 11.1 (1) 15.4 (1)
Prunus serotina 0 (0) 25.0 (3) 58.3 (7) 66.7 (6) 33.3 (3) 36.5 (1)
Ulmus americana 0 (0) 41.7 (5) 8.3 (1) 11.1 (1) 11.1 (1) 15.4 (1)
crement core were standardized to remove age-related trends (Chhin et al. 2008). Standardization was based on applying a linear, locally weighted regression (Loess) with a neighborhood span equivalent to 10 years using the func- tion ‘loess’ (package STATS) in the program R (version 3.1.1) (Venables and Ripley 2002; R Core Team 2016).
This standardization approach ensured that decadal scale trends related to the silvicultural treatments were still preserved. Basal area growth index (BAGI) was calculated as the ratio of their observed versus predicted values from the Loess model. BAGI values derived from the two incre- ment cores of each tree were averaged together then sum- marized further by determining averages for red oak and competitors for each of the 5 silvicultural treatments.
Relationships between annual BAGI of red oak and competitors in each of the silvicultural treatments (i.e., re- sponse variable) and past climate records (both monthly and seasonal variables: i.e., independent variables) were as- sessed using a step-wise multiple linear regression approach (Chhin et al. 2008). The multiple regression analysis, with forward selection, was conducted using the function stepAIC (package MASS) in the program R (Venables and Ripley 2002; R Core Team 2016). The multiple regression model developed for each climate variable set (mean tem- perature and CMI) and at each scale (monthly and season- al) was based on minimizing the value of Akaike’s in- formation criterion (AIC) as long as adding each successive predictor variable lowered the AIC value by at least 2 (Burnham and Anderson 2002). The regression analyses were conducted for monthly and seasonal climate variables
over two growing seasons for climate variables in April of the prior year (t-1) to October of the current year (t) of ring formation. The regression analyses were also conducted over a 30 year period (1980-2009). There were a total of 19 predictor variables for the regression models based on monthly climate variables. There were a total of 17 pre- dictor variables for the regression models based on seasonal climate variables. The regression analysis for either temper- ature or CMI was initially applied separately at each scale (monthly, 2-month seasonal scale, and 3-month season scale). The significant variables identified at each scale then served as the pool of potential predictor variables for devel- opment of the final regression model for either temperature or CMI. Standardized () partial regression coefficients were also calculated to help assess the relative importance of the predictor variables in each regression model (Zar 1999).
Results
Growth and Mortality Responses to Silvicultural Treatments
The species composition of nearest competitor species in the unmanaged control treatment was dominated by sugar maple (A. saccharum) which represented 40.0% of all of the competitors (Table 1). American elm (Ulmus Americana L.) was the most common competitor species in the Clearcut-A treatment (41.7%). Black cherry (P. seroti- na) was the most common competitor species in the remain- ing treatment types: i.e., Shelterwood-A (66.7%), Clearcut-B (58.3%), and Shelterwood-B (33.3%). Overall,
Table 2. Basal area of red oak stands receiving different silvicultural treatments Species
group
Stand basal area (m2/ha)
df SS F p
Control Clearcut-A Shelterwood-A Clearcut-B Shelterwood-B
Red oak 5.36 (2.58) 4.78 (1.7) 3.32 (2.46) 5.36 (3.06) 7.27 (2.54) 4, 13 28.4, 78.7 1.17 0.367 Competitors 15.69 (6.17) 24.49 (4.19) 22.96 (3.06) 20.15 (3.09) 19.9 (5.38) 4, 13 176.2, 291.5 1.97 0.160 All species 21.05 (6.45) 29.27 (3.02) 26.28 (5.43) 25.51 (3.45) 27.17 (4.44) 4, 13 147.0, 294.1 1.63 0.227 Stand basal area based on stand inventory conducted in 2011.
Clearcut-A (clearcut without additional release treatment); Clearcut-B (clearcut with additional release treatment); Shelterwood-A (shelterwood without additional release); Shelterwood-B (shelterwood with additional release). Standard deviation in parentheses.
df, degrees of freedom (between groups, within groups); SS, sum of squares (between groups, within groups); F, f ratio.
Table 3. Mean cumulative diameter growth, mean cumulative mortatily rate, and mean annual mortality rateof red oak stands receiving differ- ent silvicultural treatments
Growth and mortality Clearcut-A Clearcut-B Shelterwood-A Shelterwood-B df SS F p
Initial tree sample size (n) 185 210 165 159
Cumulative diameter growth (cm)
1992 (21 yrs,3 yrs) 7.5 (2) 7.9 (0.7) 6.4 (0.3) 8.6 (0.1) 3, 10 7.8, 13.6 1.91 0.192 1998 (27 yrs, 9 yrs) 9.6 (2.7) a 11.9 (1) ab 9.1 (1.3) a 13.5 (0.6) b 3, 10 40.0, 28.4 4.70 0.027 2003 (32 yrs, 14 yrs) 11.1 (3.4) 14.2 (1.7) 11.3 (3) 15.9 (0.6) 3, 10 54.4, 61.5 2.95 0.085 2015 (44 yrs, 26 yrs) 15.9 (6.2) 21.7 (4.1) 14.3 (3.4) 23.2 (0.1) 3, 10 184.7, 188.0 3.27 0.067 Cumulative mortality (%)
1992 (21 yrs, 3 yrs) 39.1 (30.7) 13.7 (3.6) 33.1 (16.0) 15.1 (6.3) 3, 10 1,794.6, 3,458.7 1.73 0.223 1998 (27 yrs, 9 yrs) 66.5 (18.7) a 20.3 (7.1) b 67.4 (13.4) a 25.8 (7.9) b 3, 10 6,903.8, 1,685.0 13.66 <0.001 2003 (32 yrs, 14 yrs) 71.9 (15.1) a 29.6 (2.7) b 77.7 (12.7) a 31.6 (13.6) b 3, 10 6,822.2, 1,397.8 16.27 <0.001 2015 (44 yrs, 26 yrs) 83.9 (10.9) a 48.7 (10.1) b 84.7 (10.2) a 47.5 (11.8) b 3, 10 4,565.0, 1,142.6 13.32 <0.001 Annual mortality (%)
1992 (21 yrs, 3 yrs) 9.2 (8.1) 2.4 (0.7) 6.7 (4) 2.7 (1.2) 3, 10 121.4, 232.1 1.74 0.221 1998 (27 yrs, 9 yrs) 10.0 (1.6) a 1.4 (1.0) b 11.9 (3.1) a 2.3 (1.2) b 3, 10 295.1, 33.1 29.70 <0.001 2003 (32 yrs, 14 yrs) 3.2 (0.8) 2.4 (1.2) 8.9 (7.0) 2.0 (2.3) 3, 10 96.5, 115.7 2.78 0.096 2015 (44 yrs, 26 yrs) 5.1 (1.8) 2.7 (1.6) 3.8 (2.2) 2.1 (0.4) 3, 10 18.5, 27.9 2.22 0.149 Mean diameter growth and mortality rate are reported during 4 inventory time periods (1992, 1998, 2003, 2015); and the number of years since original reproduction harvests in 1971 and final release treatment conducted in 1989 are also provided in parentheses. Cumulative mor- tality rate was expressed relative to a baseline inventory conducted in 1986. Mean annual mortality rate was determined according to Sheil and May (1996).
Clearcut-A (clearcut without additional release treatment); Clearcut-B (clearcut with additional release treatment); Shelterwood-A (shelterwood without additional release); Shelterwood-B (shelterwood with additional release). Standard deviation in parentheses. For each variable and time period, if the main treatment effect is significant in the ANOVA, multiple comparison tests are provided and silvicultural treatments with different letters are statistically significant (p<0.05).
df, degrees of freedom (between groups, within groups); SS, sum of squares (between groups, within groups); F, f ratio.
combined across all treatment types, P. serotina (36.5% of all competitors) was the most prevalent competitor of red oak.
Based on the stand inventory conducted in 2011 (40 years since regeneration harvest, 22 years since last release treatment), stand basal area of red oak was not significantly
different across the treatment types (F (4, 13)=1.17, p=0.367) (Table 2). Stand basal area of competitors was not significantly different across the treatment types (F (4, 13)=1.97, p=0.160). Total stand basal area (red oak and competitor species) was not significantly different across
Fig. 3. Interannual basal area growth index (BAGI) growth chronology of (a) red oak and (b) competitor tree species across the different silvicultural treatments. Clearcut-A (clearcut without additional release treatment), Clearcut-B (clearcut with additional release treatment), Shelterwood-A (shelterwood without additional release), Shelterwood-B (shelterwood with additional release).
Fig. 2. Cumulative basal area growth trajectories of (a) red oak and (b) com- petitor tree species across the different silvicultural treatments. Clearcut-A (clearcut without additional release treatment), Clearcut-B (clearcut with additional release treatment), Shelterwood-A (shelterwood without addi- tional release), Shelterwood-B (shelterwood with additional release).
the treatment types (F (4, 13)=1.63, p=0.227).
This study involved the monitoring of DBH and mortal- ity of a total of 719 red oak trees across the 4 treatment types: Clearcut-A, Shelterwood-A, Clearcut-B, and Shelterwood-B (Table 3). Cumulative DBH of red oak in the 1998 inventory year was significantly greater in Shelterwood-B compared to Shelterwood-A and Clearcut-A (all p<0.05 for Fisher’s HSD tests) (Table 3).
In the 1992 inventory year, there was no significant differ- ence in mortality rates between the treatment types (F (3, 10)=1.73, p=0.223) (Table 3). In the subsequent in- ventory years (1998, 2003, 2015), the cumulative mortality rate of red oak trees in Clearcut-B and Shelterwood-B were significantly lower than that found in Clearcut-A and Shelterwood-A (all p<0.05 for Fisher’s HSD tests).
Annual rates of mortality were significantly lower in both Clearcut-B and Shelterwood-B compared to both Clearcut-A and Shelterwood-A for the mortality census conducted in 1998 (F (3, 10)=29.70, p<0.001) (Table 3).
Two-way ANOVA analyses indicated that there was no sig-
nificant interaction between regeneration harvest technique (clearcut or shelterwood) and whether or not competition release was performed for any of the survey periods (results not presented).
After the second release treatment was conducted in 1989, mean cumulative tree basal area in Clearcut-B and Shelterwood-B increased past Clearcut-A and Shelter- wood-A (Fig. 2a). By 2009 (38 years since regeneration harvest, 20 years since last release treatment), mean cumu- lative tree basal area of Clearcut-B was 0.038 m2 and Shelterwood-B was 0.053 m2. For competitors, there was limited separation between each of the treatment groups (Fig. 2b). After the second release treatment, cumulative average tree basal area of competitors was greater than oak for the same treatment type especially for oaks in Clearcut-A and Shelterwood-A treatment categories. By 2009, competitors had a tree basal area range of 0.065-0.080 m2.
Interannual basal area growth index (BAGI) patterns are shown in Fig. 3. During the prominent drought year of
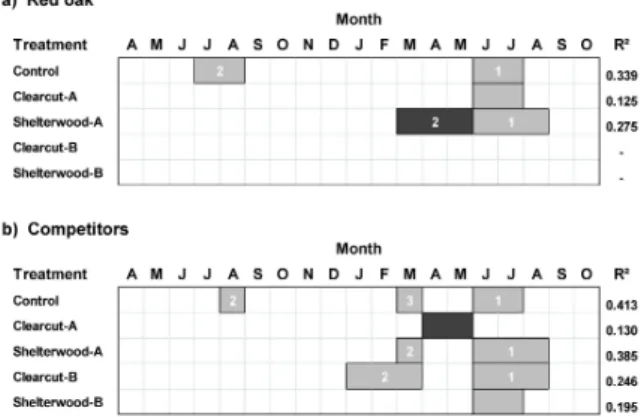
Fig. 4. Regression models between basal area growth index (BAGI) of (a) red oak and (b) competitors with temperature variables. Climate variables spanned April of the prior year (t-1) to October of the current year(t)). For each regression model, predictor climate variables having a positive relation- ship with growth are denoted by light gray boxes, and predictor climate vari- ables having a negative relationship with growth are denoted by darker gray.
The numbers within the boxes denote the ranking of the climate variables according to their standardized regression () coefficient (highest=rank 1).
Fig. 5. Regression models between basal area growth index (BAGI) of (a) red oak and (b) competitors with moisture index variables. Climate variables spanned April of the prior year (t-1) to October of the current year(t)). For each regression model, predictor climate variables having a positive relation- ship with growth are denoted by light gray boxes, and predictor climate vari- ables having a negative relationship with growth are denoted by darker gray.
The numbers within the boxes denote the ranking of the climate variables according to their standardized regression () coefficient (highest=rank 1).
1988, interannual BAGI showed a decline in red oak (Fig.
3a), but this was not as pronounced as the growth decline found in competitors (Fig. 3b).
Radial Growth-Climate Relationships
Radial growth of red oak in the control treatment and Clearcut-B was negatively related to summer temperature of the previous year (t-1) (Fig. 4a). Furthermore, Clearcut-B radial growth was also negatively associated with fall temperature (i.e., September and October 2-month seasonal period) of the current year. Red oak growth in Clearcut-A, Shelterwood-A, and Shelterwood-B were not sensitive to temperature.
Growth of competitors in Clearcut-A, Shelterwood-A, and Clearcut-B were negatively related to summer temper- ature (t-1) (Fig. 4b). Shelterwood-A and Clearcut-B also were negatively associated with July (t) temperature. For three of the silvicultural treatments (Clearcut-A, Shelterwood-A, and Clearcut-B), radial growth of com- petitor tree species was positively related to spring temper- ature (t) of the current year. BAGI of competitor tree spe- cies in Shelterwood-A was also positively associated with fall (October) (t) temperature.
Red oak in Control, Clearcut-A, and Shelterwood-A were sensitive to moisture stress in the summer (t) (Fig.
5a). Furthermore, in Control and Shelterwood-A treat- ments, red oak responded primarily to summer moisture index since this variable had the highest standardized () regression coefficient (Zar 1999). Red oak in Clearcut-B and Shelterwood-B were not sensitive to moisture stress.
Competitors in all treatment types were affected by the moisture index, mainly in the form of moisture stress in the summer (t) (Fig. 5b).
Discussion
Growth and Mortality Responses to Silvicultural Treatments
The comprehensive survey of red oak conducted in this study indicated that the initial regeneration harvest treat- ments in combination with the additional competition re- lease (i.e., Clearcut-B and Shelterwood-B) significantly re- duced cumulative mortality rates of red oak 27-44 years af- ter the initial regeneration harvesting treatments (9-26 years after the crop tree release treatment). Red oak re- quires adequate light availability to drive aboveground shoot growth and reduce mortality risk. While the light compensation point for red oak is approximately 5% of full sunlight, 20% of full sunlight is needed for a positive car- bon balance to drive shoot growth (Dech et al. 2008).
Removal of competing trees thus provides adequate light regime for red oak growth and survival. Midstory competi- tion removal studies have indicated that red oak (Dech et al.
2008; Craig et al. 2014) and other oak species (Parrott et al.
2012; Lhotka and Loewenstein 2013) show increased sur- vival (and thus reduced mortality rates) in response to these silvicultural treatments. Lhotka (2013) found that density of oak species was related to the gap size of the thinning treatment with oak species showing preference for inter- mediate gap sizes. Survival of cork oak (Quercus suber L.) was negatively impacted by interspecific competition in southern Spain (Ibanez et al. 2014).
The stand-level survey of cumulative diameter growth of red oak showed that the shelterwood with additional com- petition release resulted in significantly greater cumulative diameter growth rates during the monitoring period of 27 years after the initial regeneration harvesting treatments (9 years after the crop tree release treatment) compared to the shelterwood without the release treatment. Furthermore, analyses of tree-level basal area growth trajectories from the tree-ring data showed that red oak had the higher basal area in the shelterwood with additional release treatment com- pared to the shelterwood without release. Rentsch et al.
(2002) also showed that a successively greater degree of crown release resulted in a corresponding increase in radial growth of red oak. Other studies have shown the beneficial effect of competition release mainly through thinning treat- ments towards improved diameter growth of red oak (Gauthier and Jacobs 2010; Cogliastro and Paquette 2012) or other oak species (Parrott et al. 2012). Craig et al.
(2014), in contrast, reported that competition removal had a beneficial effect on height growth while groundline diam- eter was not significantly greater. In sessile oak (Quercus petraea [Matt.] Liebl.) stands in Poland, wider initial plan- tation spacing did not improve diameter growth due to in- terspecific competition from willow and aspen in the lower density stands (Andrzejczyk et al. 2015).
The results emphasized the importance of silvicultural management inputs (i.e., regeneration harvest treatments following competition release treatments), in allowing a clear positive tree growth trajectory of red oak that trans- lated into greater overall stand-level growth. However, the high stand-level basal area presence of other competitor species and their high tree-level growth trajectories indicate
that additional thinnings may be necessary to ensure con- tinued survival and productivity of the red oak component of the stand (Parker and Merritt 1994; Johnson et al. 2002;
Dolos et al. 2016). According to the metric conversion of Gingrich’s (1967) stocking diagram, and based on the most recent 2015 stand inventories, the stocking level is ex- cessively overstocked in the Clearcut-A (>110% stocking) and Shelterwood-A (>110% stocking) treatment catego- ries, while just above the fully stocked line in the Clearcut-B (105% stocking) and Shelterwood-B (105 % stocking) treatment categories. Intensity of oak management needs to be optimized, given the propensity of oak to produce epi- cormic shoots if thinning treatments are too intense (Johnson et al. 2002).
Radial Growth-Climate Relationships
Red oak in the control and clearcut (Clearcut-A) and shelterwood (Shelterwood-A) without additional competi- tion release treatment were sensitive to summer moisture stress primarily in the summer of the current growing sea- son (t). For both control and shelterwood without addi- tional release, this response was also the primary climatic ef- fect influencing radial growth of red oak. Red oak in un- managed forests in other studies have also shown this classi- cal response to summer moisture stress (Pan et al. 1997;
Abrams et al. 1998; Speer et al. 2009; Fekedulegn et al.
2003; Tardif and Conciatori 2006; Tardif et al. 2006;
LeBlanc and Terrell 2011). Similarly, competitors across most of the treatment types were sensitive to moisture stress, primarily in the summer (t). P. serotina was the key competitor in many of the treatment groups, and other studies have also confirmed the sensitivity of this species to summer drought (Pan et al. 1997; Abrams et al. 1998).
From a global perspective, drought has also led to radial growth decline in oak species in Europe including holm oak (Quercus ilex subsp. ballota (Desf.) Sampl L.) in Spain (Camarero et al. 2015; Camarero et al. 2016).
The results indicated that with investing in the additional competition release treatment around red oak trees in clear- cut and shelterwood harvested stands, not only does this lead to increased tree- and stand-level productivity of red oak, but in addition, radial growth of red oak was no longer sensitive to the moisture index. Furthermore, red oak in shelterwood plots receiving the additional competition re-
lease treatment was also not sensitive to monthly or seasonal temperature. These results support the hypothesis that sil- vicultural management treatments (especially shelterwood followed by additional competition release) that moderate the degree of competition for resources can lead to a general decoupling of radial growth from the dry and hot regional climate (Magruder et al. 2013). Nevertheless, management intensity should be carefully considered. In plantation man- agement systems requiring repeated entries to conduct thinning operations, excessive thinning intensities to very low densities can have a negative impact on climatic resil- ience (Magruder et al. 2013) and climatic resistance (D’Amato et al. 2013). The silvicultural management in- puts so far appear to have benefited red oak in this study in the form of reduced sensitivity to climatic stress; however, future studies should carefully examine whether excessive, repeated thinning entries could offset these initial benefits.
In the control treatment, red oak growth also showed a lag effect to both summer moisture stress and high summer temperature stress in the previous year (t-1). Furthermore, red oak growth in clearcut stands with the additional release treatment was affected primarily by summer temperature (t-1). This lagged effect is associated with a number of po- tential ecophysiological processes in red oak (LeBlanc and Terrell 2011). First, carbohydrate reserves developed in the prior growing season can be used to drive radial growth in the following year (LeBlanc and Terrell 2011). High tem- peratures can increase the rate of respiration, which can in turn deplete carbohydrate reserves (Fritts 1976; Pallardy 2007). Second, foliage development in oak is a multiyear process and environmental conditions at the time of leaf pri- mordia formation in buds can influence the extent of the photosynthetic surface area the following growing season (Pallardy 2007; LeBlanc and Terrell 2011). LeBlanc and Terrell (2011) also noted lagged relationships with precip- itation and reported a lagged response to autumn precip- itation (t-1) in red oak across eastern North America. Pan et al. (1997) also reported direct relationships between red oak and precipitation in both summer and fall of the pre- vious year (t-1).
For competitor trees in the shelterwood with no release treatment and clearcut with additional release treatment, the negative relationship with June (t) temperature strongly suggests that high temperatures can potentially induce
moisture stress (Fritts 1976; Pallardy 2007). Radial growth of the competitors were positively related with spring tem- perature (t) which indicates that competitors are sensitive to the timing of the start of the growing season and radial growth benefits from warm spring temperatures. Another study of P. serotina noted a negative relationship between ra- dial growth and summer temperature (Pan et al. 1997).
From a global perspective, radial growth of oak species in Europe are also sensitive to high temperature stress includ- ing Portugese oak (Quercus faginea Lam.) (Camarero et al.
2016).
Although the competitors of red oak were sensitive to cli- matic stress, they were still able to maintain a high stand-level basal area presence and have higher tree-level basal area growth rates. The results thus suggest that higher growth rates of the competitors can help offset the degree of increased sensitivity to climatic stress. In order to ensure that the competitive balance remains in favor of red oak, fu- ture thinnings are required (Parker and Merritt 1994;
Johnson et al. 2002).
Conclusion
This study underscored the importance of crop tree re- lease following a clearcut or shelterwood harvest in re- ducing mortality rates and thus increasing survival of red oak in mixed hardwood stands where the more shade toler- ant black cherry (P. serotina) was the dominant competing species. Further, crop tree release conducted after the shel- terwood harvest improved diameter growth rates at both the stand- and tree-level. In addition to reducing mortality and increasing diameter growth, crop tree release after the shelterwood regeneration harvest showed no sensitivity to either summer moisture stress or high temperature stress while the shelterwood without competition release was sen- sitive to summer moisture stress. The results indicate that silvicultural practices can moderate the effect of inter- specific competition and decouple the sensitivity of red oak to climatic stress. The results suggest that this study has im- portant implications for ensuring that oak forests remain adapted to future weather events during the regeneration to pole-sized phase of stand development of red oak.
Acknowledgements
This study was funded through Michigan State University (MSU) AgBioResearch (Project # MICL06002). I thank P. Bloese and G. Kowalewski for assistance with field logis- tics and D. Berry, E. David, and M. Magruder for field data collection and sample processing.
References
Abrams MD, Ruffner CM, Morgan TA. 1998. Tree-ring re- sponses to drought across species and contrasting sites in the ridge and valley of central Pennsylvania. For Sci 44: 550-558.
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim JH, Allard G, Running SW, Semerci A, Cobb N. 2010. A global overview of drought and heat-induced tree mortality re- veals emerging climate change risks for forests. For Ecol Manage 259: 660-684.
Andrzejczyk T, Liziniewicz M, Drozdowski S. 2015. Effect of spacing on growth and quality parameters in sessile oak (Quercus petraea) stands in central Poland: results 7 years after planting. Scand J For Res 30: 710-718.
Brose PH, Dey DC, Phillips RJ, Waldrop TA. 2013. A meta-anal- ysis of the fire-oak hypothesis: does prescribed burning promote oak reproduction in eastern North America. For Sci 59:
322-334.
Buckley DS, Sharik TL, Isebrands JG. 1998. Regeneration of northern red oak: positive and negative effects of competitor removal. Ecol 79: 65-78.
Burnham KP, Anderson DR. 2002. Model selection and multi- model inference: a practical information-theoretic approach.
2nd ed. Springer-Verlag, New York, 488 pp.
Sander IL. 1990. Quercus rubra L. or Nothern red oak. In: Silvics of North America. 1. conifers (Burns RM, Honkala BH, eds).
USDA, Washington, DC.
Camarero JJ, Franquesa M, Sanguesa-Barreda G. 2015. Timing of drought triggers distinct growth responses in Holm oak:
Implications to predict warming-induced forest defoliation and growth decline. Forests 6: 1576-1597.
Camarero JJ, Sanguesa-Barreda G, Vergarechea M. 2016. Prior height, growth, and wood anatomy differently predispose to drought-induced dieback in two Mediterranean oak species.
Annals For Sci 73: 341-351.
Chhin S, Hogg EH, Lieffers VJ, Huang S. 2008. Potential effects of climate change on the growth of lodgepole pine across diame- ter size classes and ecological regions. For Ecol Manage 256:
1692-1703.
Chhin S, O’Brien J. 2015. Dendroclimatic analysis of red pine af-
fected by Diplodia shoot blight in different latitudinal regions in Michigan. Can J For Res 45: 1757-1767.
Chmura DJ, Anderson PD, Howe GT, Harrington CA, Halofsky JE, Peterson DL, Shaw DC, St. Claire JB. 2011. Forest re- sponses to climate change in the northwestern United States:
Ecophysiological foundations for adaptive management. For Ecol Manag 261: 1121-1142.
Cogliastro A, Paquette A. 2012. Thinning effect on light regime and growth of underplanted red oak and black cherry in post-agricultural forests of south-eastern Canada. New For 43:
941-954.
Craig JM, Lhotka JM, Stringer JW. 2014. Evaluating initial re- sponses of natural and underplanted oak reproduction and a shade-tolerant competitor to midstory removal. For Sci 60:
1164-1171.
Crunkilton DD, Pallardy SG, Garrett HE. 1992. Water relations and gas exchange of northern red oak seedlings planted in a cen- tral Missouri clearcut and shelterwood. For Ecol Manag 53:
117-129.
D’Amato AW, Bradford J, Fraver S, Palik B. 2013. Effects of thin- ning on drought vulnerability and climate response in north temperate forest ecosystems. Ecol Appl 23: 1735-1742.
Dech JP, Robinson LM, Nosko P. 2008. Understory plant com- munity characteristics and natural hardwood regeneration under three partial harvest treatments applied in a northern red oak (Quercus rubra L.) stand in the Great Lakes-St. Lawrence for- est region of Canada. For Ecol Manag 256: 760-773.
DeRose RJ, Long JN. 2014. Resistance and resilience: a con- ceptual framework for silviculture. For Sci 60: 1205-1212.
Dey DC, Spetich MA, Weigel DR, Johnson PS, Graney DL, Kabrick JM. 2009. A suggested approach for design of oak (Quercus L.) regneration research considering regional differences. New For 37: 123-135.
Dey DC. 2014. Sustaining oak forests in eastern North America:
regeneration and recruitment, the pillars of sustainability. For Sci 60: 926-942.
Dolos K, Mette T, Wellstein C. 2016. Silvicultural climatic turning point for European beech and sessile oak in Western Europe from national forest inventories. For Ecol Manage 373:
128-137.
Fekedulegn D, Hicks Jr RR, Colbert JJ. 2003. Influence of topo- graphic aspect, precipitation and drought on radial growth of four major tree species in an Appalachian watershed. For Ecol Manage 177: 409-425.
Finley K, Chhin S, Nzokou P. 2016. Effects of climate on the radial growth of white ash infested with emerald ash borer. For Ecol Manage 379: 133-145.
Folke C, Carpenter S, Walker B, Scheffer M, Elmqvist T, Gunderson L, Holling CS. 2004. Regime shifts, resilience, and biodiversity in ecosystem management. Ann Rev Ecol Evo Syst 35: 557-581.
Fritts HC. 1976. Tree rings and climate. Academic Press, London,
UK.
Gauthier MM, Jacobs DF. 2010. Northern red oak, white oak, and black walnut diameter growth for the first 3 years after thinning in a mixed planting. North J Appl For 27: 110-116.
Gingrich SF. 1967. Measuring and evaluating stocking and stand density in upland hardwood forests in the central states. For Sci 13: 38-53.
Grissino-Mayer HD. 2001. Evaluating crossdating accuracy: a manual and tutorial for the computer program COFECHA.
Tree-Ring Res 57: 205-221.
Handler S, Duveneck MJ, Iverson L, Peters E, Scheller RM, Wythers KR, Brandt L, Butler P, Janowiak M, Swanston C, Clark Eagle A, Cohen JG, Corner R, Reich PB, Baker T, Chhin S, Clark E, Fehringer D, Fosgitt J, Gries J, Hall KR, Hall C, Heyd R, Hoving CL, Ibanez I, Kuhr D, Matthews S, Muladore J, Nadelhoffer K, Neumann D, Peters M, Prasad A, Sands M, Swaty R, Wonch L, Daley J, Davenport M, Emery MR, Johnson G, Johnson L, Neitzel D, Rissman A, Rittenhouse C, Ziel R. 2014. Michigan Forest Ecosystem Vulnerability Assessment and Synthesis: A report from the Northwoods Climate Change Response Framework Project.
Gen Tech Rep NRS-129. USDA, Forest Service, Northern Research Station, Newtown Sguare.
Hill JP, Dickmann DI. 1988. A comparison of three methods for naturally reproducing oak in southern Michigan. North J Appl For 5: 113-117.
Hogg EH. 1997. Temporal scaling of moisture and the for- est-grassland boundary in western Canada. Agri For Meteor 84:
115-122.
Holmes RL. 1983. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 43: 69-78.
Ibanez B, Ibanez I, Gomez-Aparicio L, Ruiz-Benito P, Garcia LV, Maranon T. 2014. Contrasting effects of climate change along life stages of a dominant tree species: the importance of soil-cli- mate interactions. Diversity Distrib 20: 872-883.
IPCC. 2013. Climate Change 2013: The Physical science basis.
Contribution of working group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; New York, NY, USA. Cambridge University Press, Cambridge.
Johnson PS, Shifley SR, Rogers R. 2002. The ecology and silvi- culture of oaks. CABI Publishing, Oxonm, UK.
Kabrick JM, Zenner EK, Dey DC, Gwaze D, Jensen RG. 2008.
Using ecological land types to examine landscape-scale oak re- generation dynamics. For Ecol Manage 255: 3051-3062.
Kling GW, Hayhoe K, Johnson LB, Magnuson JJ, Polasky S, Robinson SK, Shuter BJ, Wander MM, Wuebbles DJ, Zak DR. 2003. Climate change in the Great Lakes region: impacts on our communities and ecosystem. A report of the Union of Concerned Scientists and the Ecological Society of America, Washington, D.C.
Laurent M, Antoine N, Joel G. 2003. Effects of different thinning intensities on drought response in Norway spruce (Picea abies
(L.) Karst.). For Ecol Manage 183: 47-60.
LeBlanc DC, Stahle DW. 2015. Radial growth responses of four oak species to climate in eastern and central North America. Can J For Res 45: 793-804.
LeBlanc DC, Terrell MA. 2011. Comparison of growth-climate relationships between northern red oak and white oak across eastern North America. Can J For Res 41: 1936-1947.
LeBlanc DC. 1998. Interactive effects acidic deposition, drought, and insect attack on oak populations in the midwestern United States. Can J For Res 28: 1184-1197.
Lhotka JM, Loewenstein EF. 2013. Development of three under- planted hardwood species 7 years following midstory removal.
South J Appl For 37: 81-90.
Lhotka JM. 2013. Effect of gap size on mid-rotation stand struc- ture and species composition in a naturally regenerated mixed broadleaf forest. New For 44: 311-325.
Magruder M, Chhin S, Monks A, O’Brien J. 2012. Effects of ini- tial stand density and climate on red pine productivity within Huron National Forest, Michigan, USA. Forest 3: 1086-1103.
Magruder M, Chhin S, Palik B, Bradford JB. 2013. Thinning in- creases climatic resilience of red pine. Can J For Res 43: 878-889.
Matisons R, Jansons J, Katrevičs J, Jansons Ā. 2015. Relation of tree-ring width and earlywoodvessel size of alien Quercus rubra L. with climatic factors in Latvia. Silva Fennica 49(4): article id 1931, 14 p.
Metsaranta JM, Lieffers VJ. 2009. Using dendrochronology to obtain annual data for modelling stand development: a supple- ment to permanent sample plots. Forestry 82: 163-173.
Millar CI, Stephenson NL, Stephens SL. 2007. Climate change and forests of the future: managing in the face of uncertainty.
Ecol Appl 17: 2145-2151.
Pallardy SG. 2007. Physiology of Woody Plants. 3rd ed. Academic Press, San Diego, CA.
Pan C, Tajchman SJ, Kochenderfer JN. 1997. Dendroclimato- logical analysis of major forest species of the central Appalachians. For Ecol Manag 98: 77-87.
Parker GR, Merritt C. 1994. The central region. In: Regional silvi- culture of the United States (Barrett JW, ed). John Wiley &
Sons, New York, pp 129-172.
Parker WC, Dey DC. 2008. Influence of overstory density on eco- physiology of red oak (Quercus rubra) and sugar maple (Acer saccharum) seedlings in central Ontario shelterwoods. Tree Physiol 28: 797-804.
Parrott DL, Lhotka JM, Stringer JW, Dillaway DN. 2012.
Seven-year effects of midstory removal on natural and under- planted oak reproduction. North J Appl For 29: 182-190.
R Core Team. 2016. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/. Accessed 19 March 2016.
Rentch JS, Desta F, Miller GW. 2002. Climate, canopy dis- turbance, and radial growth averaging in a second-growth mixed-oak forest in West Virginia, U.S.A. Can J For Res 32:
915-927.
Sheil D, May RM. 1996. Mortality and recruitment rate evalua- tions in heterogeneous tropical forests. J Ecol 84: 91-100.
Speer JH, Grissino-Mayer HD, Orvis KH, Greenberg CH.
2009. Climate response of five oak species in the eastern decid- uous forest of the southern Appalachian Mountains, USA. Can J For Res 39: 507-518.
Speer JH. 2010. Fundamentals of tree-ring research. The University of Arizona Press, Tucscon, AZ.
Spetich MA, Dey D, Johnson P. 2009. Shelterwood-planted north- ern red oaks: integrated costs and options. South J Appl For 33:
182-186.
Stokes MA, Smiley TL. 1996. An Introduction to Tree-Ring Dating. The University of Arizona Press, Tucson, AZ.
Tardif JC, Conciatori F, Nantel P, Gagnon D. 2006. Radial growth and climate responses of white oak (Quercus alba) and northern red oak (Quercus rubra) at the northern distribution limit of white oak in Quebec, Canada. J Biogeogr 33: 1657-1669.
Tardif JC, Conciatori F. 2006. Influence of climate on tree rings and vessel features in red oak and white oak growing near their northern distribution limit, southwestern Quebec, Canada. Can J For Res 36: 2317-2330.
Venables WN, Ripley BD. 2002. Modern applied statistics with S.
4th ed. Springer, New York.
Yamaguchi DK. 1991. A simple method for cross dating increment cores for living trees. Can J For Res 21: 414-416.
Zar JH. 1999. Biostatistical analysis. 4th ed. Prentice Hall, Upper Saddle River, NJ.