346
†
전체 글
346
†
수치
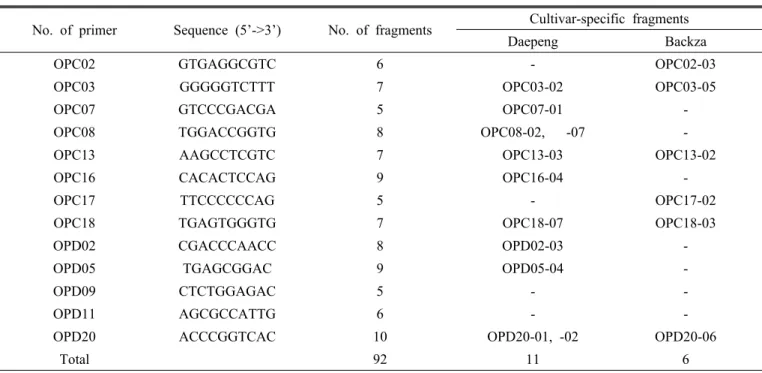
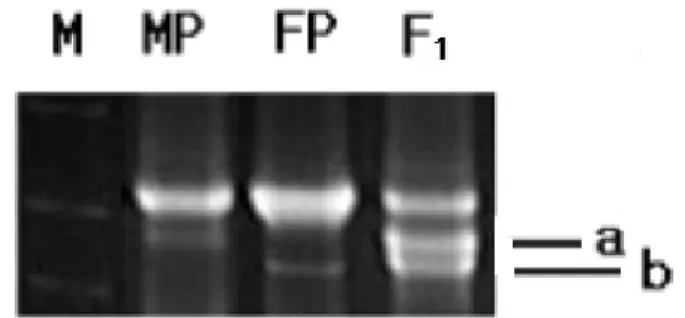
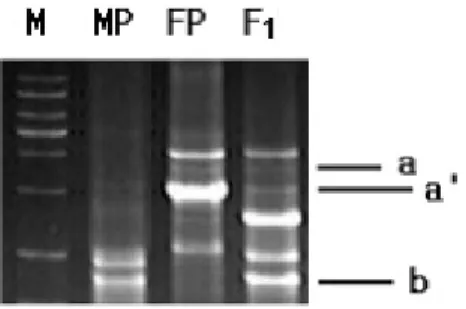
관련 문서
Therefore, head and neck massage is an exercise program that is effective for brain function activation and stress factors in middle-aged women, and it is judged that it can be
The results show that the green seed extracts are only shown as antioxidants, but the others are shown as a prooxidants against DNA damage and Escherichia coli
Seed germination, cutting propagation and early growth characteristics were investigated in order to develop the method of cutting propagation and to find
replicate designs obtained using the classical method of confounding are shown to be particularly useful for deriving suitable balanced designs for
Biochemical markers of bone turnover can be classified according to the process that underlie in markers of bone formation, products of the osteoblast
Third, this study developed a music education program using block coding that can be used in music class, explained its content and functions and
This study, therefore, suggests that cooperative art activity based on collage can be used in the child education field for the improvement of
In this study, it can be seen that sex and metabolic syndrome are important for biological factors, and occupation, smoking, and exercise are important
