Kor.
J
Appl. Microbiol. Bioeηig.Vol. 15, No. 4, 267-272 (198η
Purificationand Properties of Extracellular
β-Xylosidasefrom Thermophilic Alkalophilic Bacillus Sp. K-17
Sung
,
Nack Kie,
l* In SOO Kang,
l Hyo Kon Chun,
lTeruhiko Akiba3 and Koki Horikoshi3
J Department 01 Food Science and Technology, Gyeong Sang National University, Chinju, Korea
2 Department 01 Food and Nutrition, Chinju Technical Co//ege, Chi,ηju 62α Korea JThe Institute 01 Physical and Chemical Research, Wako, Japan
高週, 알칼리성 Bacillus Sp. K-17 이 生成하는 홉體外 β-Xylosidase의 精製 및 特性
成洛쫓 1
* •훌仁秀2
•全孝坤 1
•秋훌빠훌3
•提越弘혔3
l
경상대학교 식품공학과 Z
진주실업전
문대학 식품영앙학과 3일본 이화학연구소 미샘물샘태학연구실
Aß-xylosidase was purified 22-fold from culture filtrate of
thermoph패c
a1kalophilic Bacillus sp.K-17 by ammonium sulfate precipitation
,
DEAE-Sephadex A-50 chromatography and Sephadex G-100 gel filtration. The molecular weight was calculated to be 51,
000 by SDS gel electrophoresis.The enzyme had a pH optimum for activity at 7.0 and retained appreciable activity at pH 10.5
,
and its stability range was pH 6.0-10.0. The optimal temperature of enzyme was 450C,
and its ac띠vity was completely inactivated by incubation at 60 oC for 10 min. The Michaelis constant for p-nitrophenyl-ß-o-xylopyranoside was calculated to be 0.45 mM. ß-Xylosidase could degrade xylo-oligosaccharide
,
produced from xylan by the action of xylanase,
to xylose.Xylan can be degraded to xylose by the sequen- losidase into the medium. There was so far few tial reaction of xylanase (1,4-o-xylan xylanohydro- report on the
extracell띠ar
production of ß-xylosi-l없e;
EC 3.2.1.8) and ß-xylosidase (1,4-o-xylan dase by bacteria.xylohydrol ase; EC 3.2.1.37). ß-Xylosidase degrade ln the present report, a ß-xylosidase from ther- not only xylobiose but also xylo-oligo-saccharides mophilic alkalophilic Baci//us sp. K-17 has been from their reducing end to release xylose unit (1). purified from the culture supernatant and enzyma- Severalß-xylosidase from different rnicroorganisms tic properties have been characterized.
(1-8) have been purified and characterized.
Thermophilic alkalophilic Baci//us sp. K-17
,
Materials and Methods which we isolated previously (9), produced twotypes of xylanase (10) and a ß-xylosidase. Although Microorganism and cultivation
ß-xylosidases from Baci//us subti/is (11) and Baci//us sp. K-17
str밍n , is이ated
in. the previous Baci//uspumi/;μs
(7.8) were reported to be located in study (9),
was used. The strain was grown aerobi- intracellularcomp따tment ,
our strain secreted ß-xy- cally for 48hr at 45 oC in때
alkaline medium consis-I{ey words: ß-Xylosidase, thermophilic aJka10philic Bacillus sp.,
• Corresponding author
267
ting of: wheat bran, 1O.Og; yeast extract, 5.0g;
polypeptone
,
5.0g; K2HP04,
1.0g; MgS04.7Hp,
0.2g; and Na2C03
,
1O.Og per 1 liter of water.Sodium carbonate was sterilized separately and ad- ded to the medium.
ß-Xylosidase assay
The activity of ß-xylosidase was measured with p-nitrophenyl-ß-D -xylopyranoside (PNPX) in 50 mM phosphate buffer (pH 7.0). The reaction mix- ture
,
composed of 1 ml substrate solution (1 mg/ml) and 1 ml properly diluted enzyme,
was in- cubated at 40 C for 10 min. The reaction was stop- ped by addition of 2 ml 0.5M Na2C03 and the op- tica1 density of p-nitrophenol released Was mea- sured at 405 nm. One unit of enzyme was defined as the amount of enzyme capable of releasing 1 mole p-nitrophenol in 1 min.Enzyme purification
All purification steps were carried out at 40C.
The buffer used throughout the purification was 50 mM phosphate buffer (pH 6.5).
Step 1. Ammonium sulfate precipitatiòn
The culture broth was centrifuged at 8 ,αm
rpm for 15min to remove cell mass and wheat bran debris. To the supernatant
,
arnmoniuni sulfate was added to 8011,10 satu- ration. After standing for one day
,
the resulting precipitate was collected by centrifugation at 10,α)() rpm,
dissolved in the buffer and dia1yzed for 48hr against the sarne buffer. The insoluble solid was re- moved by centrifugation.Step 2. OEAE-Sephadex A-50 chromatography.
The dia1yzed enzyme solution was charged onto a OEAE-Sephadex A-50 column (2.2 x 8Ocm) equilibrated with the buffer.
After the column was fully wåshed with
Protein measurement
Protein concentration was measured by. the method of Lowry et al. (2) with bovine serum albumin as the standard. The protein concentration in the column chromatographys was followed by optical density at 280nm.
Gel electrophoresis
Polyacrylarnide gel electrophoresis was carried out in a 7.7% acrylarnide gel by the method of Oavis (13). Webber and Osborn’s method (14) of sodium dodecyl sulfate (SOS) polyacrylamide disc gel electrophoresis was used for the molecular weight estimation of the enzyme protein. Bovine a1bumin .(molecular weight 66,아)()), egg a1bumin
(45 ,α)()) , glyceraldehyde-3-phosphate dehydro- genase (36,α)()) , trypsin inhibitor (20
,
100) and a-lactoalbumin (14,200) were used as the molec비arweight marker proteins. Protein staining was performed with Amidoblack lOB.
Hydrolysis of xylan
To 1 m/of 1 % xylan solution was added O.lml (5 units) of xylanase 1 and 11, r~spectively , and the mixture was incubated at 60 oC for 5hr. To each of theresultant reaction mixture was added 0.1 ml (2.5 units) ofβ-xylosidase , and the reaction was further continued at 40 oC for 19 hr.
Chemicals
Xylooligosaccharides were donated by Or. T.
Akiba of the Institute .of Physica1 and Chemica1 Re- search
,
Japan,
and molecular weight marker pro- teins, p-nitrophenyl-,β-D-xylopyranoside and larch- wood xylan were purchased from Sigma ChemiCa1 Co. (St. Louis. USA). OEAE-Sephadex A-50,
Sep- hadex 0-100 were purchased from Parmacia Fine Co. (Sweden). Other chemic외s used were of an외ytica1 grade and obtained commercia11y.
the buffer
,
ß-xylosidase was eluted with a Results linear gradient of from 0 to 0.5M NaCl inthe buffer. Purification of β-xylosidase
Step 3. Oel filtration. The result of enzyme purification is summarized ß-Xylosidase was purified by ge1 filtration in Table 1. β-xylosiqase was purified 22-fold in spe- on Sephadex 0-100 column (2.0 x 5Ocm) cific activity with yi~ld of 7.4%.
equilibrated with the buffer. The homogeneity of the purified β-xylosidase
Vol. 15, No. 4 269
Table 1. Summary of purification of the β-xylosidase from Bacillus sp. K-17
Purification Total Total
volume Activity
steps
(ml) (U)
Culture f!uid 1,800 2,160
(NH4)2S04
precipitation 200 640
DEAE-Sephadex 150 375
Sephadex G-I00 80 160
Total protem
(mg) 2,114 176 24 7.3
Specific actlVlty (U/mg)
1.0 3.6 15.6 21.9
Yield
(%)
100.0 29.6 17.4
7.4
was examined by disc and SDS polyacrylamide gel 5.0 electrophoresis. Fig. 1 shows the polyacrylamide
disc gel electrophoretic patterns of ß-xylosidase.
Molecular weight estimation
Relative electrophoretic mobilities of purified ß-xylosidase and of the reference proteins on SDS polyacrylamide gel electrophoresis were plotted ver- sus their molecular weights. The mobility of the purified ß-xylosidase corresponded to a molecular
Fig. 1. Polyacrylamide disc gel electrophoresis of the purifìed ß-xylosidase.
s bIl
.,
4.8
$
4·6g
잉
.,
-
E g @」
4.4
A
β -xylosidase B
C
、R
、f
I
0.2 0.4 0.6 0.8
Relative mobility (Rm)
Fig. 2. Estimation of the molecular weight of the puri- liedß.xylosidase by SDS polyacrylamide gel electropho- resls.
A: bovine albumin (M.W., 66,000), B: egg albumin (45,000), C: glyceraldehyde-3-phosphate dehydrogenase (36,000), D: trypsin inhibitor (20,100), B: lactoalbumin
(14,200)
weight of 51 ,α)() (Fig. 2).
Effect of pH
Optimum pH. The activities of β -xylosidase were measured at various pHs in McIlvaine buffer
φH 3.0-8
,
0) and McIlvaine-NaOH buffer (pH 8.0- pH 10.5). The result is shown in Fig. 3. ß -Xylo- sidase had optimal activity at pH 7.0, and broad pH curves with relatively high activities in the alkaline reglOn.pH stability. ß-Xylosidase was incubated at various pHs in McIlvaine and McIlvaine-NaOH buffer for 30 min at 45 oC and the remaining acti꺼-
1.0
100
~ 80
;
형
> 60여
-
잉즙 ~O
훌 20
0
5 6 7 8 9 10 11 pH
Fig. 3. Optimal pH and pH stability of the purified ß-xylosidase.
。-0 ; OptimaJ temperature
• - .; pH stability
ties were assayed. Fig. 3 shows ß -xylosidase was stable at pH 6.0 to 10.0. The remaining activity at pH 10.5 of ß-xylosidase was 65070.
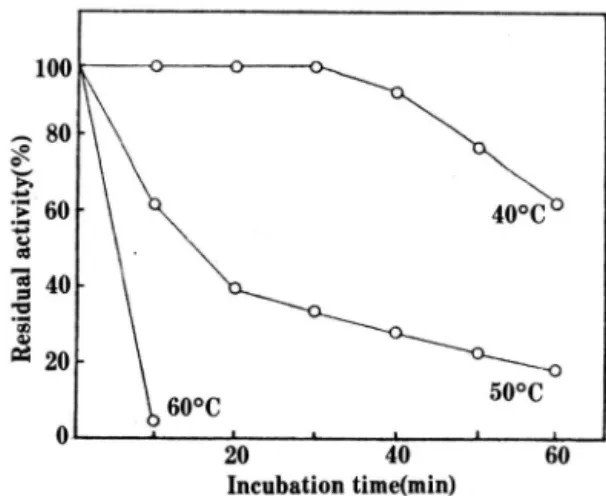
Effect of temperature.
Optimum temperature. The effect of temper- ature on the enzyme activities is shown in Fig. 4.
The optimal temperature ofß-xylosidase was 45 o
c.
Thermal stability. Thermal stability of ß-xylo- sidase was exaniined by incubation in McIlvaine buffer (pH 6.5) at various temperatures. The resi- dual activity was measured at the interval of 10 min. The thermal stability of
β
-xylosidase was100
‘
ε 。 80
.형F--혀ii
60공여a-aua
40120
0 30 40 50 60
Temperature (OC)
Fig. 4. Optimal temperattJre of the purified β-xylo
sidase.
~ 80
s
ζ.;: 60
g 빼
열
40 -= ∞월
200
400C
600C 500C
20 40 60
Incubation time(min)
Fig. 5. Thermal stability of the purified ß-xylosidase.
stable at 450 for 30 rnin
,
but was not stable up to 500C. ß-xylosidase was completely inactivated by incubation at 600C for 10 min (Fig. 5).Effect of various compounds
After the enzyme solutions were mixed with 50 mM phosphate buffer (pH 6.5) containing the test reagents at the final concentration of 5 mM and preincubated at 300C for 30 min. The residual activities were measured. As summarized in Table 2
,
pCMB,
Co ++,
Cu ++,
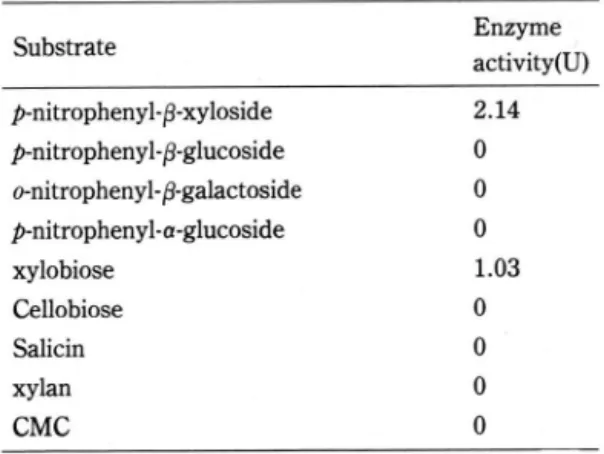
Ag + and Hg + + showed inhibitory effect on the -xylosidase.Substrate specificity
Sever외
a andβ ~glycosides
were tésted to ex- Table 2. Effect of various compounds on the activity of purified ß -xylosidaseCompounds Activity Relative activity
(5mM) (U) (%)
Control 0.41 100
MnS04.4H20 0.41 100
ICH2COOH 0.37 90
EDTA 0.37 90
pCMB* 0.05 12
CoCl2 0.17 42
CuCl2 0.15 37
AgN03 0.16 39
FeS04'7H20 0.36 88
HgCI2 0 0
‘
Sodiump-
chloromercuric benzonate.Vol. 15, No. 4
Table 3. Substrate specificity of the purified
a웅ylosidase
Substrate Enzyrne
activity(U)
p-nitrophenylβ-xyloside
2.14p-nitrophenylβ-glucoside
0o-nitrophenyl-βgalactoside
0 p-nitrophenyl-a-glucoside 0xylobiose 1.03
Cellobiose 0
Salicin 0
xylan 0
CMC 0
amine the Substrate specificityof the enzyme (see Table 3)The enzyme was active on PNPX and xylo-
biose
,
with little activity on cellobiose,
p-nitro- phenyl-ß-glucoside,
o-nitrophenyl-ß-galactoside and p-nitophenyl-a-glucoside. Enzyme had no acti- vity on xylan o.r carboxmethyl cel1ulose.Kinetic constants
lnitial velocities in the hydrolysis of PNPX at various concentration of from 0.2 mM to 0.9 mM were measured
,
and the Michaelis constant (Km) was estimated from Lineweaver-Burk plot(l5) of the data. The Km caculated was 0.45 mM PNPX.Hydrolysis of xylan with xylanase and ß:xylosidase As previously reported(lO), xylanase 1 from our strain yie1ded xylobiose
,
xylotriose and xylotetraose from xylan,
and xylanase 11 did yield xylose,
xylo- biose and xylotriose. We studied the effect of the action of ß-xylosidase on these products in the xylan hydroly~tes. In the early stage of hydrolysis,
xylosidase (2.5 unifs) was added to the reaction mixtures ofxylanase 1 and 11. It was recognized that addition of ß -xylosidase increased the degree of xylan degradation
,
from 22070 to 28% and 30% to 33%,
as shown in Fig. 6,
and the final product were xylose only both in the two reactions. This indi- cates that xylo-oligosaccharides produced from xylan via xylanase 1 and 11 were hydrolyzed to xylose by β-xylosidase.-
s、
효
혀‘
40
30
∞
20∞ ;.,
-
~ ;.,
=
271
5 6 7 ’ 10’ 15 ’ 24 TiRle (hr)
Fig. 6. Hydrolysis of xylan with xylanases 1, 11 and ß -xylosidase.
Arrow indicate addition of the ß -xyjosidase.
() - () ; xylanase 1
• - • ; xylanase II
。
-
0; 8-xylosidaseDiscussion
The bacterial strains so far reported to produce
β-xylosidase produced intracell벼ar or cell bounded enzyme
,
but our strain secreted ß-xylosidase into medium as Bacillus circulans WL-12(l6),
which was solely reported to produceextracel1ular ß-xylo- sidase. But ß-xylosidase from Bacillus circulans was so labile that the remaining activity could not be detected after 72 hr at refrigerator temperature (40C), thus neither purified nor characterized.An extracel1ular ß-xylosidase from thermophilic alkalophilic Bacillus sp. K-17 was purified and characterized. The purified enzyme was homogene- ous on polyacrylamide disc gel electrophoresis. The enzyme had an optimal pH for activity at 7.0
,
but appreciable acitivity was detected at pH 10.5. The enzyme was stable in the range of pH 6.0-10.0. The optimum pHofβ-xylosidase
from Chaetomium tri- laterale (4) and Aspergillus fumigatsu (5) was 4.5,
which was lower value than that of ours. The op- timal temperature (450C) of the enzyme was lower than 550C of Chaetomium trilaterale and 750C of Asp. fumigatus (5). The Km value of the purified ß-xylosidase against PNPX was 0.45 mM. This
value was lower than 2.4 mM for ß-xylosidase from B. pumilus (8) and 2.0 mM from Asp. jumigatus (5), and higher than 0.2 mM from Asp. niger (1).
Some ß-xylosidase from other origin (4.19) were capable of hydrolyzing ß-o-glucopyranoside, but our enzyme doesn
’
t show such activity.ß-xylosidase could degrade the hydrolysis products (up to xylo- tetraose) from larchwood xylan with xylanase 1 and xylanase II to xylose.We applied gene cloning system using Escheri- chia coli as host in order to clarify the mechanism of secretion ofß7xylosidase and isolatedß-xylosidase gene from thermophilic alkalophilic Bacillus sp.
K-17.
요 약
고온, 알깔리 성
Bαcill:αs sp. K-17이 생성 하는 균체 외
β-xy
losidase를유안염 석
DEAE-Sephadex chromatography빛
gel filtration에 의해 정제하였고
SDS-polyacrylamide전기영동에 의해 분차량을
측정하였든 바
51, 000으로 추정되었다.효소의
최적 pH는 7.0이 었으며
pH 10. 5에서도상당한 활성을 나 타내였고 효소의 안정성은
pH6.0와 pH 10. 0 사이였다. 정제된 효소는
450C에서 최적 반응을 나타내었고 그 활성은
600C에서 10분만에 실활되는 것
으로보아 매우 불안정한 효소이 었다.
p-Nitrophenyl-
ß
-D-xylopyranoside에 대 한
Km치 는 0. 4
5mM이였고
ß -xylosidase는 xylanase의 xylan분해산물인
xylo-oligosaccharide를 xylose로완전분해
시켰으며 xylanase에 의 한
xylan분해시
ß
-xyl osidase의 첨가는
xylan분해율에 있어서 상 승효과가 있었다.
References
1. Takenish, S.Y. Tsujisaka and J. Fukumoto: J. Bio-
chem.
,
73,
335 (1973)2. John, M.B. Schmidt and J. Schmidt: Can. Bio- chem., S7, 125 (1979)
3. Kawaminami T. 없ld H. lizuka: J. Fermn. Tech.
,
48
,
169 (1910)4. Uziie, M.M. Matsuo and T. Yasui: Agric. Biol.
Chem.
,
49,
1159 (1985)5. Kitpreechavanich, V.M. Hayashi and S. Nagai:
Agric. Biol. Chem.
,
50,
1703 (1986)6. Matsuo
,
M.T. Yosui and T. Kobyashi: Agric. Biol.Chem.
,
41,
1601 (1977)7. Kersters-Hilderson, H.F.G. Loontiens, M. Claeys- sens and C.K. Debruyne: Eur. J. Biochem.
,
7,
434 (1969)8. Panbangred, W.O. Kawaguchi, T. Tomita, A. Shin- myo and H. Okada: Eur. J. Biochem.
,
138,
267 (1984)9. Sung, N.K., I.S. Kang, H.K. Chun, T. Akiba and K.
Hotikoshi: Kor. J. Appl. Microbiol. Bioeng., 12(2), 81-89 (1984)
10. Kang, I.S., N.K. Sung, H.K. Chun, T. Akiba and K.
Horikoshi: Kor. J. Appl. Microbiol. Bioeng., 14(6), 447-453 (1986)
11. Roncero M.I.G.: J. Bacteriol.
,
156,
257 (1983) 12. Lowry, O.H., N.J. Rosebrough, Al L. Farr and R.J.d허1: J. Biol. Chem.
,
193,
265 (1951)13. Davis B.J.: Ann. N. Y. Acad. Sci.
,
121,
404 (1964) 14. Weber K. and M. Osborn: J. Biol. Chem.,
244,
4406(1969)
15. Lineweaver H. and Burk: J. Am. Chem. Soc.
,
56,
658 (1934)
16. Esteban R., J.R. Villanueva and T.G.Villa: Can. J.
Microbiol., Z8, 733 (1982)
17. Okazaki, W.T. Akiba, K. Horikoshi and R. Aka- hoshi: Appl. Microbiol. Biotechnol.
,
19,
335 (1984) 18. Uchino F. and T. Nakane: Agric. Biol. Chem.,
45,
1121 (1981)
19. DeBruyne, C.K., G.M. Aerts and R.L. Degussem: Eur. J. Biochem.
,
102,
157 (1979)(Received June 29