Paenibacillus woosongensis 로부터 대장균에 Xylanase 10A의 유전자 클로닝과 정제 및 특성분석
윤기홍*
우송대학교바이오식품과학전공
Received: February 20, 2020 / Revised: May 26, 20 / Accepted: June 3, 2020
서 론
셀룰로스, 리그닌과함께리그노셀룰로스자원을구성하 는헤미셀룰로스는그주성분이 xylan, mannan, β-glucan, xyloglucan, galactan 및 arabinan으로알려져있다. 식물의 종류에따라헤미셀룰로스의성분과조성에큰차이가있지 만 xylan이각종목초와작물류에가장널리분포한다. Xylan 은 D-xylose 잔기간에β-1,4 결합의다당체이며 acetyl, L- arabinofuranosyl, 4-O-methyl-α-D-glucuronopyranosyl 또
는 D-xylosyl 등의여러종류치환체잔기가 xylose 잔기의 2번또는 3번탄소위치에측쇄로결합되어있는복합적인구 조를가지고있다. Xylan은 xylose에결합된치환체의종류 에따라 glucuronoxylan, arabinoxylan, glucuronoarabinoxylan 과 galactoglucuronoarabinoxylan으로구분된다. 이와달리 치환체잔기가없는 homoxylan은β-1,3 또는β-1,3과β-1,4
결합한 xylose 잔기만으로구성되며자연계에서드물게존
재한다. Xylan은자일리톨, 바이오연료, 바이오계면활성 제인알킬자일로사이드, 자일로올리고당의생산원료로활 용되고있으며, 효과적인활용을위해서는효소적전환과정 이필요하다[1].
Xylans의분해에는다양한종류의효소가관여하는데이
들중 D-xylosyl 잔기간 배당결합을절단하는 endo-β-1,4- Gene Cloning, Purification and Characterization of Xylanase 10A from Paenibacillus woosongensis in Escherichia coli
Ki-Hong Yoon*
Food Science and Biotechnology Major, Woosong University, Daejeon 34606, Republic of Korea
A gene coding for the xylanase was cloned from Paenibacillus woosongensis, followed by determination of its complete nucleotide sequence. This xylanase gene, designated as xyn10A, consists of 1,446 nucleotides encoding a polypeptide of 481 amino acid residues. Based on the deduced amino acid sequence, Xyn10A was identified to be a modular enzyme composed of a catalytic domain highly homologous to the glycosyl hydrolase family 10 xylanase and a putative carbohydrate-binding module (CBM) in the C-terminus. By using DEAE-sepharose and phenyl-sepharose column chromatography, Xyn10A was purified from the cell- free extract of recombinant Escherichia coli carrying a P. woosongensis xyn10A gene. The N-terminal amino acid sequence of the purified Xyn10A was identified to exactly match the sequence immediately following the signal peptide predicted by the Signal5.0 server. The purified Xyn10A was a truncated protein of 33 kDa, suggesting the deletion of CBM in the C-terminus by intracellular hydrolysis. The purified enzyme had an optimum pH and temperature of 6.0 and 55−60℃, respectively, with the kinetic parameters Vmax and Km of 298.8 U/mg and 2.47 mg/ml, respectively, for oat spelt xylan. The enzyme was more active on arabinox- ylan than on oat spelt xylan and birchood xylan with low activity for p-nitrophenyl-β-xylopyranoside.
Xylanase activity was significantly inhibited by 5 mM Cu2+, Mn2+,and SDS, and was noticeably enhanced by K+, Ni2+, and Ca2+. The enzyme could hydrolyze xylooligosaccharides larger than xylobiose. The predomi- nant products resulting from xylooligosaccharide hydrolysis were xylobiose and xylose.
Keywords: Paenibacillus woosongensis, xylanase 10A, purification, characterization
*Corresponding author
Tel: +82-42-630-9742, Fax: 82-42-630-9389 E-mail: [email protected]
© 2020, The Korean Society for Microbiology and Biotechnology
xylanase (xylanase)가가장중요한역할을한다. Xylanase 는활성영역만으로구성된단일영역효소[2−4], 또는활성
영역과별개로탄수화물결합영역 (CBM)과같은기능성영
역을동시에포함하는다영역효소로존재한다[5, 6]. 활성
영역의아미노산배열의상동성을기준으로할때 xylanase
는 glycosyl hydrolase family (GH) 2, 5, 7, 8, 10, 11, 16, 30, 35, 43과 52에 속하는 효소로 보고되었으며, 이들 중 GH10과 GH11에 속하는 xylanase가 주류를 이룬다. Xylanase는그크기가다양하여약 200−1,500 아미노산잔 기로구성되어있는데이러한크기의차이는활성영역의크 기차이가아니라주로기능성영역의존재여부에따라좌 우된다.
최근미생물의유전체정보를기반하여재조합효소를발 현하고활성을분석함으로써한종류미생물에서 2 종류이 상의다형 xylanases 생산예가다수보고되었다. 특히식물
체 xylan은측쇄종류와조성의차이가크므로자연계미생
물이이를효과적으로분해하기위해서β-xylosidase, α-L- arabinofuranosidase, α-D-glucuronidase, acetylxylan esterase, feruloyl esterase와α-galactosidase 등의보조효소도필요 하며, 동시에기질의특이성을비롯한반응특성에차이가있 는다형의 xylanases를생산하는것이유리하다고하겠다. Xylan 분해균으로 탐색된 Paenibacillus속 균주 중 P.
panacisoli SD1201, P. curdlanolyticus B-6, P. barcinonensis BP-23, P. amylolyticus KCTC 3005, Paenibacillus sp.
JDR-2, Paenibacillus sp. DG-22, Paenibacillus sp. W-61, Paenibacillus sp. HC1과 Paenibacillus sp. KCTC 8848P 로부터 2종류이상의 xylanase 유전자와효소특성이밝혀 졌다[3, 7−12]. 최근에는분해가잘되지않는것으로알려
진 옥수수 섬유소의 xylan을 효율적으로 분해하는
Paenibacillus sp. H2C가분리되었으며, 배양상등액으로부 터 4종류의 xylanases (Xyn5A, Xyn10B, Xyn11A, and Xyn30A)가확인되었다[13]. 본연구에서는이미두종류의 GH11 xylanases의유전자와효소특성이보고된바있는, P. woosongensis로부터신규 GH10 xylanase 유전자를클로 닝하고재조합대장균에서생산된효소를정제하여그특성 을조사하였다.
재료 및 방법
사용 균주와 플라스미드
Tryptic soy broth (TSB; 17 g of tryptone, 3 g of soytone, 2.5 g of dextrose, 5 g of NaCl, 2.5 g of K2HPO4 per liter, pH 7.2)를사용하여 37℃에서약 20시간진탕배양하여얻 은 P. woosongensis YB-45 (KCTC 3953)의균체로부터 유 전체 DNA를 분리하였다. Escherichia coli DH5α(F− λ−
glnV44 thi-1 recA1 gyrA96 deoR nupG purB20 φ80dlacZΔM15 Δ(lacZYA-argF)U169 hsdR17(rm− mk+))와 플라스미드 pUC19는유전자조작을위한숙주와 vector로사 용되었다.
Xylanase 유전자 클로닝
P. woosongensis의부분적인유전체염기서열로부터유추 된신규 xylanase 유전자를클로닝하기위해 upper primer YB45C37-10F (CTTGCTGCAGATGGAATCGCGCAAAAG AAAGC; 밑줄은 PstI 위치)와 lower primer YB45C37-10R (ACTCTCTAGATTAATTCCACAAATATACGTTATCC; 밑 줄은 XbaI 위치)을합성하였다. P. woosongensis의유전체 DNA를주형, YB45C37-10F와 YB45C37-10R을 primers로 하고 pfu-X DNA polymerase (Solgent Co., Korea)를사용 하여 xylanase 유전자를증폭하였다. PCR은 95℃에서 3분 간처리후 95℃ (25초), 56℃ (40초), 72℃ (1분 30초)의반 응을 25회반복하고최종적으로 72℃에서 20분간반응함으 로써수행하였다. 증폭된 DNA 단편을 PstI과 XbaI으로동 시에절단하여 agarose gel 전기영동을한후 xylanase 유전 자를함유한 DNA를추출하여동일한효소로처리한 pUC19 에도입하였다. 이를 E. coli DH5α에형질전환하고 oat spelt xylan (0.5%)과 ampicillin (100 μg/ml)이첨가된 LB 배지에
도말하여투명환을보이는콜로니를 xylanase 유전자가클
로닝된형질전환주로선발하였다.
Xylanase의 정제
P. woosongensis의 xylanase 유전자가도입된재조합대 장균을 ampicillin이첨가된 LB 배지에접종하여 37℃에서 18시간동안배양한후균체를회수하고 50 mM Tris 완충 액(pH 8.0)에현탁하였다. 초음파로균체를파쇄하고원심분 리한 후균체 파쇄상등액을채취하여 ammonium sulfate (30−75%)로침전시키고투석하였다. 동일완충용액으로평 형화한 DEAE-Sepharose 컬럼(Sigma-Aldrich Co., USA)에 단백질용액을주입하고컬럼에흡착되지않은효소용액을 회수한후고농도의 ammonium sulfate 용액을첨가하여최 종적으로 1 M ammonium sulfate와 20 mM Tris (pH 8.0) 을포함하는용액이되도록하였다. 효소액과동일용액으로 활성화된 Phenyl-Sepharose 컬럼에효소액을주입하고정 제를 실시하였으며 컬럼에 흡착된 효소는 ammonium sulfate의농도를 1.0 M에서 0 M까지역으로농도구배를주 어용출하였다. Xylanase 활성을갖는분획을 SDS-PAGE로 분석하여정제된분획만을 10 mM sodium phosphate 완충 액 (pH 6.0)으로투석하여정제효소로사용하였다. 정제된 효소의아미노말단의아미노산배열을결정하기위해서는 기초과학연구소서울분원의 Procise 491 protein sequencer
(Applied Biosystems, USA)를사용하였다.
Xylanase 반응특성 분석
Xylanase 활성은적정반응조건에서 15분동안반응후에 xylan (0.5%)으로부터유리된환원당을 3, 5-dinitrosalicylic acid 방법으로정량하여결정하였다. 발색단기질인 para- nitrophenyl-β-xylopyranoside (pNPX)의분해활성을측정하
기위해서는기질농도를 1 mM로하고적정반응조건에서
10분동안반응후반응액의 2배부피의 1 M Na2CO3 용액 을첨가하여반응을종결시키고 405 nm에서흡광도를측정 함으로써방출된 pNP의양을결정하였다. 효소의활성도 1 unit는 1분동안 1 μmol의환원당또는 pNP를유리시키 는효소량으로정의하였다[4, 14]. 반응온도와 pH가효소활 성에미치는영향을조사하기위하여 30−65℃와 pH 4.5−9.0 의범위에서 xylanase 활성을각각측정하였다. 열안정성은 55−65℃범위의온도에서 xylanase 효소액을방치하면서시 간별로채취한효소액의잔존활성을측정하여결정하였다. 금속이온이나화합물을반응액에 5 mM이되도록첨가하여 효소활성을측정함으로써각물질이효소활성에미치는영 향을결정하였다. 효소의반응동력학계수는 oat spelt xylan 을기질로하여 최적조건에서초기 반응속도를측정한후 Lineweaver-Burk plot에서결정하였으며정제효소의단백 질농도는 Bradford 방법으로결정하였다. Xylanase에의한 최종가수분해산물을조사하기위해 1% 자일로올리고당을 과량의정제효소로 45℃에서 4시간동안처리하였다. 반응 액을 95℃에서 3분간방치하고원심분리하여효소단백질을 제거한 후 반응상등액을 silica gel-precoated thin layer plate (Merck, Gemany)에점적하여 TLC를수행하였다. 전 개용매로는 chloroform, acetic acid와증류수(4.3 : 5 : 0.7, (v/v)) 혼합용액을사용하였고발색제로는 9 ml ethanol, 0.5 ml p-anisaldehyde, 0.5 ml sulfuric acid와 glacial acetic acid 몇방울을혼합한용액을사용하였다[4].
Xylanase 활성 염색
효소단백질의 SDS-PAGE를수행한후, polyacrylamide gel을 25% isopropanol와 50 mM sodium phosphate (pH 6.0) 혼합용액으로세척하고추가적으로 50 mM sodium phosphate (pH 6.0)로세척하였다. Polyacrylamide gel 위 에 0.2% 수용성 oat spelt xylan과 50 mM sodium phosphate (pH 6.0)를함유한 agarose gel을중층한후 50℃에서 1시간 반응하였다. Polyacrylamide gel은단백질염색을하였으며, 중 층한 agarose gel은 0.2% congo red 용액으로 30분간염색 하고 1 M NaCl 용액으로탈색하여 xylan 분해단백질을관 찰하였다.
결과 및 고찰
P. woosongensis xylanase 유전자의 클로닝과 염기서열 Paenibacillus 속의균주중에서 P. curdlanolyticus B-6의 7 종류 xylanases (Xyn10E, Xyn10C, Xyn11A, Xyn10D, Xyn10A, Xyn10B, PcMulGH9) [3, 5, 6], P. barcinonensis BP-23의 5종류 xylanases (Xyn10A, Xyn10B, Xyn10C, Xyn30D, Xyn11E) [2, 7, 8], Paenibacillus sp. H2C의 4 종 류 xylanases (Xyn5A, Xyn10B, Xyn11A, Xyn30A) [13], 그리고 Paenibacillus sp. W-61의 3 종류 xylanases (Xyn1, Xyn3, Xyn5) [10, 11]에대한유전자나효소특성이조사되 었다. 이들효소는 GH10, GH11, GH30 또는 GH5에속하는 것으로 GH10 xylanase가가장많았다. P. woosongensis의 유전정보로부터이미보고된 2 종류의 GH11 xylanases외에 GH10 xylanase로추정되는유전자가확인되어이를 PCR 로 클로닝하기 위한 primers를 제조하였다. Primers YB45C37-10F와 YB45C37-10R를사용하여 PCR을수행한 결과예상되는크기의 1.5-kb xylanase 유전자단편이증폭 되었으며이를 PstI과 XbaI으로절단하여 pUC19에도입함 으로써재조합플라스미드 pYX7을제조하였다.
클론된유전자단편의염기서열을결정한결과 481개아 미노산잔기의단백질을코드하는 1,446 bp의 xylanase 유 전자가확인되었다 (GenBank accession number MF062088).
유전자로부터유추된아미노산배열을 SignalP5.0 프로그램 (http://www.cbs.dtu.dk/services/SignalP/)으로분석한결과 아미노 말단의 32개 잔기가 signal peptide로예측되었다 (Fig. 1). Signal peptide는아미노산배열이매우다양하므
로그상동성을비교하기어려운것으로알려져있지만[15],
예측된 signal peptide는염기성아미노산이배열한아미노 말단과 소수성 아미노산이 배열한 중앙 지역 및 signal peptidase의 절단 지점 앞에 AXA 배열을 포함하고 있어 signal peptide의전형적인특징을보였다[16, 17]. 한편개 시코돈으로부터 6 bp가떨어진상부지역에리보좀결합위 치로판단되는염기서열 (AGGAGGT)이관찰되었다. 다른 xylanase와 P. woosongensis xylanase의 상동성
클론된 P. woosongensis의 xylanase는 GH10에속하는 xylanase의활성영역(33−318 잔기)과 CBM으로추정되는 영역(319−481 잔기)이배열된다영역효소로 확인되었으 며 Xyn10A로 명명하였다. P. woosongensis의 Xyn10A (PwXyn10A)를미국의 NCBI database에등록된다른효소 와아미노산배열을비교한결과 P. macerans의 xylanase (KFM94218; 484 잔기)와상동성이 76%로가장높았다. 이 외에 Paenibacillus sp. KCOM 302 (RRJ62552; 478 잔기),
Paenibacillus sp. 598K (GBF76989; 478 잔기) 및 Planifilum fulgidum (SFG13237; 480 잔기)의 xylanases와는 크기가 유사하고 70−77% 수준의상동성을보여 PwXyn10A는신규 의 xylanase로확인되었다. 상기의상동성이높은 xylanase 는모두유전체에서추론된아미노산배열과비교한결과이 며효소적특성이확인된 xylanase와비교하였을때아미노 산배열과그크기가가장유사한것은발효분뇨의유전체 은행에서탐색된 xylanase (ManF-X10)이며[18], 그크기가 14개아미노산이차이가있고아미노산배열상동성이 69%
로확인되었다. 실제 ManF-X10은아미노말단이완전하게 클로닝되지못한상태이다(Fig. 1).
P. curdlanolyticus B-6의 xylanase D (PcXyn10D;
BAK22544)는활성영역과 Fn3 및 CBM3로구성되어있으 며[5], 활성영역이 PwXyn10A와 79% 정도상동성을보이지 만 CBM은상동성이거의없다. 반면에 ManF-X10는상동 성이 70%로 PcXyn10D 보다는낮지만활성영역뿐아니라 CBM의아미노산배열도 PwXyn10A와유사성이있다. 한편 활성영역으로만구성된 P. curdlanolyticus B-6의 xylanase Fig. 1. Comparison of the P. woosongensis xylanase 10A with others. The amino acid sequences of six xylanases from P. wooson- gensis (PW; AVI01408), uncultured bacterium (UB; ACJ48973), P. curdlanolyticus B-6 (PD; BAK22544, PC; BAM22646), P. barcinonensis BP- 23 (PB; CBA13561), and Paenibacillus sp. KCTC8848P (PS; AAG23527) are aligned to maximize similarities with gaps (hyphens) by Clustal W method. Residues identical to the amino acid sequence of the P. woosongensis Xyn10A are indicated by asterisks in other sequences.
Amino acid sequence corresponding to the signal peptide predicted by SignalP5.0 server is italicized and underlined. Two putative cat- alytic residues are marked by box in the amino acid. Numbers at the end of each line correspond to the amino acid position in the protein.
C (PcXyn10C; BAM22646), P. barcinonensis BP-23 xylanase A (PbXynA; CBA13561) 및 Paenibacillus sp. KCTC 8848P xylanase B (PsXynB; AAG23527)는 PwXyn10A의활성영 역과상동성이각각 76%, 74%, 74%로나타났다.
Xylanase의 정제
탄수화물에작용하는많은수의효소들이아미노산배열
의유사성에근거하여 CAZy 데이터베이스에서분류되었지
만, 실제로는일부효소만이그기능과특성에대해분석되 었다. PwXyn10A의효소특성을조사하기위해재료및방 법에기술한대로 pwxyn10A 유전자를함유한 E. coli DH5α 배양균체로부터 PwXyn10A를정제하였다. 균체파쇄상등 액을 ammonium sulfate로처리하여 얻은 단백질용액을 DEAE-Sepharose 컬럼에주입한결과 PwXyn10A는컬럼에 흡착되지않고그대로용출되었다. 결합되지않은효소액을
Phenyl-Sepharose 컬럼 크로마토그래피로 정제한 결과
PwXyn10A는 Phenyl-Sepharose 컬럼에흡착되었으며 0.6− 0.3 M ammonium sulfate 농도에서용출되었다. 정제단계
별단백질을 SDS-PAGE와활성염색으로분석한결과균체
파쇄상등액, 황산암모늄침전단백질과 DEAE-Sepharose 컬
럼에결합되지않는단백질용액에서동일하게 47 kDa 및
33 kDa의상이한분자량을갖고 xylanase 활성을보이는두 개의단백질밴드가관찰되었다(Fig. 2A). Phenyl-Sepharose 컬럼에흡착되지않은용액에서는효소활성이관찰되지않 았으며 ammonium sulfate 농도차에의해용출된분획중활
성을갖는분획에서오직 33 kDa의단백질만이정제효소
로관찰되었다(Fig. 2B).
pwxyn10A의유전자로부터 signal peptide가제거된단백
질의 분자량이 48.5 kDa으로 추정되었으며 이것은 SDS- PAGE에서활성을보이는큰단백질과유사하였다. 최종적 으로정제된 33 kDa 크기의 xylanase는분자량이예상보다 매우작으므로정제된 xylanase가 PwXyn10A의어느부분 에해당하는지확인하기위해정제효소를 SDS-PAGE로전
개한후 PVDF 막으로옮겨아미노말단의서열을분석하였
다. 그결과아미노말단배열이 GIANGSKF로확인되었으 며이는 pwxyn10A 유전자로부터유추된 PwXyn10A의 33− 40번째잔기에해당하였다. 이러한결과는 SignalP5.0 프로 그램에서아미노말단의 32 잔기가 signal peptide로예측된 결과와일치하였다(Fig. 1). PwXyn10A의활성영역으로예 상되는 33−318 aa의분자량은 31.8 kDa로계산되며정제된 단백질과거의일치하는것을알수있다. 이로보아카르복 실말단의 CBM으로추정되는영역이절단된것으로판단 되며정제된효소를 PwXyn10A-CD로명명하였다. 재조합 대장균에서생산된단백질이균체내단백질분해효소에의
해분해되어[19], 크기가다른여러개활성단백질로생산
된예가 Paenibacillus sp. DG-22의 GH10 xylanase [20]와 Clostridium josui Xyn10A [21]에서도보고되었다. 이들효 소는카르복실말단이서로다르게절단되어크기가다르지 만아미노말단이동일한 3개의활성효소단백질로생산되 었다.
비록 큰 단백질은 정제되지 못하였지만, PwXyn10A의
mature protein의 예상 분자량과 일치하고 또한 크기가
33 kDa인 PwXyn10A-CD의 아미노 말단 배열이 signal
peptide로예측된배열다음인것으로보아재조합대장균
에서생산된 PwXyn10A는세포질에서 periplasm으로이동 하는과정에서 signal peptide가제거되었을것으로예상된 다. 실제재조합대장균에서 signal peptide를함유한여러 종류의단백질이 signal peptide가제거된 mature protein의 형태로 periplasm에존재한다고보고되었다[22, 23]. 한편크 기 및 활성영역과 CBM의 배열이 PwXyn10A와 유사한 ManF-X10은카르복시말단에 hexahistidine이융합된상태 로재조합대장균으로부터생산되어 Ni-NTA 컬럼을통해 정제되었으며정제단백질의크기는약 50 kDa로보고되었 다[18]. 이로 보아 ManF-X10은 Paenibacillus sp. GH10 xylanase [20]나 PwXyn10A와는달리재조합대장균에서카
르복시말단이절단되지않았거나, Ni-NTA 컬럼크로마토
그래피를통해서카르복시말단에 His tag이융합된형태의
단백질만이정제된때문에차이가있는것으로추측된다.
Xylanase의 반응특성
Phenyl-Sepharose 컬럼크로마토그래피과정을통해최
종적으로정제된 PwXyn10A-CD를사용하여효소의특성을
조사하였다. 반응온도와 pH가 xylanase 활성에미치는영향 Fig. 2. Xylanase zymogram (A) and SDS-PAGE (B) of
PwXyn10A-CD purified from cell-free extract of E. coli (pYX7).
Lanes: 1, cell-free extract; lane 2, precipitate with ammonium sul- fate (30-75%); 3, unbound protein on DEAE-Sepharose column;
4, purified PwXyn10A-CD; M, molecular weight markers. Molec- ular size is shown in kilodaltons to the right side of the gel.
을조사한결과 Fig. 3A에보인 바와같이 55−60℃와 pH 6.0에서최대활성을보였으며 pH 완충액종류에따라활성 에차이가크게나타났다. pH 6.0의경우 citrate 완충액보 다는 phosphate 완충액의 활성이 높으며 특히 pH 8.0은 Tris 완충액이 phosphate 완충액보다약 2.3배이상의활성 을보였다. 열안정성조사를위해 55−65℃에서효소를방치 하면서시간별잔존활성을측정하였다. 그결과 60℃에서 6시간동안방치한후에도활성이완전하게유지하였으나, 65℃에서는 1시간방치하였을때잔존활성이 1−2% 수준으 로확인되었다(결과미제시). pH에대한안정성을측정하기 위해 pH 3−9 범위의완충액에서 1시간동안방치한후잔존 활성을결정하였다. pH 3과 4에서약 66%와 84%의잔존활 성을보였고 pH 6−9 범위에서는전혀실활되지않고활성 이그대로유지되었다(Fig. 3B).
GH10의활성영역으로만구성된 xylanase인 PcXyn10C [3]와 P. amylolyticus KCTC 3005의 PaXN_10 [9]은 pH 7.0과 50℃에서 모두 최대 활성을 보여 최적온도가 PwXyn10A-CD보다 약간낮았다. PcXyn10C는 60℃에서 1시간방치후에그활성이그대로유지되지만 70℃에 1시간 방치하면완전히실활되어열안정성이 PwXyn10A-CD와유 사하였다. pH에 대한 안정성은 산성과 중성 부근에서는 PcXyn10C와 PwXyn10A-CD는유사하였지만, pH 9.0에서
PwXyn10A-CD는안정하여실활이일어나지않는데비해
PcXyn10C는크게실활되어 25% 잔존활성을보이는차이가 있었다. ManF-X10은 pH 7과 40℃에서최대활성을보이며 열안정성이낮아 50℃에서 30분간방치하였을때잔존활성 이 4%로보고되었다. ManF-X10은 PwXyn10A-CD와상동 성이높음에도불구하고온도가효소활성과안정성에미치
는영향이큰차이를보였다. CBM이효소의열안정성을높
이고 pH 안정성이나반응조건에영향을미치는경우가있 는데, 정제된 ManF-X10은정제된 PwXyn10A-CD와는달리 CBM 영역을포함하고있는데열안정성이낮았다. ManF- X10의 CBM은 mannan, xylan과 cellulose에결합하는기능 을하는것으로밝혀졌다[18]. P. terrae HPL-003의 xylanase (KRICT PX-3)는 CBM을함유한 GH10의효소인데 CBM을 제거할경우 열안정성이약 40% 감소하였고, 최적온도도 55℃에서 45℃로 낮아진 것으로 보고되었다[24]. 그러나 Bacillus sp. SN5의 GH11 xylanase는 CBM을제거할경우 도리어열안정성이증가하는것으로보고된바있다[25]. 한 편 PwXyn10A의 활성영역과 76% 상동성을 보이는 P.
barcinonensis BP-23의 PbXynA는 pH 9.5와 60℃에서최대 활성을보여최적반응조건이 PwXyn10A-CD와는큰차이가 있었으며알칼리조건에서안정한유사한경향을보였다[2].
Paenibacillus sp. NF1의 xylanase (XynNF) 분자량은 37 kDa이며 PwXyn10A-CD와유사하게 60℃와 pH 6.0에서최 대활성을보였다. 반면에 70℃와 80℃에서 1시간방치후 70.6%와 42.6%의높은잔존활성을보여 XynNF는 PwXyn10A- CD 보다열안정성이높을뿐아니라 Paenibacillus 속균주 의 xylanase 중에서열안정성이높았다[26].
기질에따른반응성을조사한결과 Table 1에보인바와
같이 PwXyn10A-CD는 arabinoxylan에대한분해활성이가 장높았고, oat spelt xylan의분해능이 birchwood xylan 보 다약간 높았으며 carboxymethyl cellulose (CMC), locust bean gum과같이 xylan이아닌섬유질계물질을분해하지 못하였다. P. curdlanolyticus B-6의 Xyn10D에서 CBM을 제거하고활성영역만존재하는 PcXyn10D-rCD는 birchwood Fig. 3. Temperature and pH optima (A), and pH stability (B) of the PwXyn10A-CD. Temperature profile (open symbols) was obtained by measuring the xylanase activity at different temperatures with fixed pH 6.0 for oat spelt xylan hydrolysis. The pH profile (open symbols) was obtained by measuring the enzyme activities at various pHs with constant temperatures of 50℃. Buffers (50 mM) used were as follows: sodium citrate (-●-, pH 4.0-6), sodium phosphate (-▲-, pH 6-8), and Tris (-◆-, pH 8-9). pH Stability was determined by measuring the residual activities after pre-incubation for 1 h at different pH buffers. Each curve represents the average of three inde- pendent experiments with standard errors of 3%.
xylan과 oat spelt xylan에대한비활성이 PwXyn10A-CD와 유사하였으며[5], PcXyn10C는 birchwood xylan에대한활 성이 oat spelt xylan에비해 2배정도높았다[3]. Paenibacillus sp. NF1의 XynNF는 oat spelt xylan에대한 정제효소의 비활성이 3,081 U/mg으로다른효소들에비해매우높았으 며, birchwood xylan에대한활성도유사하였고 CMC는분 해하지못하였다[26]. ManF-X10은 wheat arabinoxylan에 대한활성이가장높고 oat spelt xylan이 birchwood xylan 에대한활성보다약간높았다[18]. 한편 pNP 화합물을분 해하지못하는다수의 xylanase와는달리 PwXyn10A-CD는 pNPX, pNP-β-glucopyranoside와 pNP-β-galactopyranoside에
대해 높지는 않지만분해 활성을 갖는 특성을 보였다. P.
curdlanolyticus B-6의 PcGH43B6 [14]는 GH43에속하는
효소로 xylanase 활성과 arabinofuranosidase, β-xylosidase 활성을모두보였으며, pNPX에대한활성은 PwXyn10A 보 다약 6배수준으로높았으나, xylan 분해활성은 PwXyn10A 에비해약 1.5% 미만에해당하는미약한수준이었다.
최적반응조건에서 oat spelt xylan을기질로하여 5−10 mg/ml의농도범위에서 PwXyn10A-CD의초기반응속도를 구해 Vmax와 Km값을결정한결과 298.8 U/mg과 2.47 mg/
ml로측정되었다. XynNF는 3,364 U/mg과 5.64 mg/ml으로 보고되어비활성과같이 Vmax 값이 PwXyn10A-CD에비해 매우 높았다[26]. P. barcinonensis BP-23의 PbXynA는 birchwood xylan을기질로하여 Vmax와 Km이 50.67 U/mg 과 2.93 mg/ml [2], ManF-X10은 rye arabinoxylan에대해 49.5 U/mg과 2.8 mg/ml로각각결정된바있다[18].
Table 1. Comparison of specific activity between PwXyn10A and others.
Substrates Specific activity (U/mg)
PwXyn10A-CDa PcXyn10Cb PcXyn10D-rCDc PcGH43B6d XynNFe
Oat spelt xylan 220.9 188.25 218.4 1.46 3,081.1
Birchwood xylan 211.6 357.90 252.2 2.86 3,045.9
Arabinoxylan 301.2 - - 0.03 -
CMC ND - - - ND
pNPX 0.05 - - 0.30 -
pNPGc 0.02 - - ND -
pNPGa 0.01 - - ND -
pNPX; pNP-β-xylopyranoside, pNPGc; pNP-β-glucopyranoside, pNPGa; pNP-β-galactopyranoside, ND; not detected, -; not tested.
a; Xyl10A-CD of P. woosongensis in this study, b; Xyn10C of P. curdlanolyticus B-6 [3], c; Xyn10D-rCD of P. curdlanolyticus B-6 [5],
d; PcGH43B6 of P. curdlanolyticus B-6 [14], e; XynNF of Paenibacillus sp. NF1 [26].
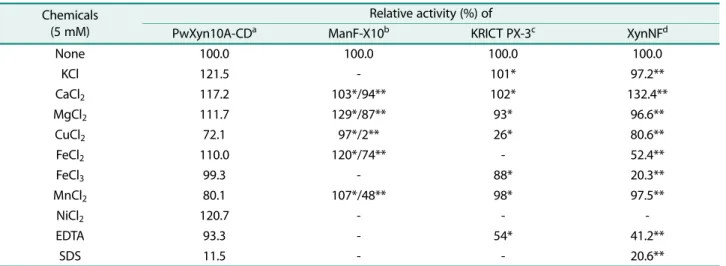
Table 2. Effects of metal ions and other reagents on the xylanase activity.
Chemicals (5 mM)
Relative activity (%) of
PwXyn10A-CDa ManF-X10b KRICT PX-3c XynNFd
None 100.0 100.0 100.0 100.0
KCl 121.5 - 101* 97.2**
CaCl2 117.2 103*/94** 102* 132.4**
MgCl2 111.7 129*/87** 93* 96.6**
CuCl2 72.1 97*/2** 26* 80.6**
FeCl2 110.0 120*/74** - 52.4**
FeCl3 99.3 - 88* 20.3**
MnCl2 80.1 107*/48** 98* 97.5**
NiCl2 120.7 - - -
EDTA 93.3 - 54* 41.2**
SDS 11.5 - - 20.6**
-; not tested.
*; effectors of 1 mM were added. **; all effectors of 10 mM were added.
a; Xyn10A-CD of P. woosongensis in this study, b; xylanase of uncultured bacterium [18], c; xylanases of P. terrae HPL-003 [27],
d; XynNF of Paenibacillus sp. NF1 [26].
금속이온을비롯한화합물이 PwXyn10A-CD 활성에미치 는영향을분석하여 ManF-X10, KRICT PX-3 및 XynNF와 비교하였다(Table 2). 5 mM의화합물이존재할때 PwXyn10A 는 Ca2+, Mg2+, Ni2+, Fe2+과 K+에의해서는활성이증가되 었으며 Fe3+에의해서는거의영향을받지않았다. 반면에 Cu2+와 Mn2+에의해활성이 20−30% 정도감소하였다. 금속 이온의 농도에 따라 차이는 있지만, Ca2+이나 Mg2+는 xylanase 활성을저해보다는향상시키는경우가많았다. Fe2+
의경우효소에따라 향상시키거나심하게저해하였으며, Fe3+와 Cu2+는대부분의효소활성을저해하였으며효소간에 차이가 컸으며, 농도에 따라서도 큰 편차를 보였다. PwXyn10A-CD는 5 mM EDTA에의해미약하게저해되는 것과는달리 KRICT PX-3 [27]과 XynNF는약 50% 수준의 활성이저해되었고, PwXyn10A-CD와 XynNF는모두 SDS 에의해서심하게활성이저해되었다.
PwXyn10A에의한 xylobiose, xylotriose, xylotetraose와 xylopentaose의 가수분해 산물을 TLC로 분석한 결과 xylobiose는거의분해하지못하였다. Xylotriose 이상의중 합도를갖는올리고당은동일하게 xylobiose와 xylose로분 해되었다(Fig. 4). Paenibacillus sp. HC1의 xylanase-I은 xylo 올리고당을 xylose와 xylobiose로분해하여 PwXyn10A- CD와유사하였다[12]. PwXyn11A는 xylotriose 이상의중합 도를갖는 XOS을분해하였으며주된분해산물이 xylose로 나타났으며, xylobiose는소량생성되었다[4]. 또한 PwXyn10A-
CD와는달리 xylotriose, xylotetraose와 xylopentaose에대 한분해능이높지는않았다.
요 약
Paenibacillus woosongensis의 xylanase 유전자를클로닝 하고그염기서열을결정하였다. Xylanase 유전자는 xyn10A 로명명되었으며, 481 아미노잔기로구성된단백질을코드 하는 1,446개뉴클레오티드로구성되었다. 추론된아미노산 배열에 따르면 Xyn10A는 glycosyl hydrolase family 10
xylanase와상동성이높은활성영역과카르복실말단에탄
수화물을결합하는것으로추정되는영역이포함된다영역 효소로 확인되었다. DEAE-Sepharose와 Phenyl-Separose 컬럼크로마토그래피과정을통해 P. woosongensis xyn10A 유전자를함유한재조합대장균의균체파쇄상등액으로부터 Xyn10A를정제하였다. 정제된 Xyn10A의아미노말단배열 이 GIANGSKF로결정되었으며이는 SignalP5.0 server로 예측된 signal peptide의다음아미노산배열과정확하게일 치하였다. 정제된 Xyn10A는 33 kDa 크기의절단된단백질 이며균체내분해에의해카르복시말단에서 CBM이제거 된것으로판단된다. 정제된효소는최적 pH와온도가 6.0 과 55−60℃이며 oat spelt xylan에대한반응동력학적계수 Vmax와 Km이 298.8 U/mg과 2.47 mg/ml로각각나타났다. 효소는 birchwood xylan이나 oat spelt xylan보다 arabinoxylan 에대한활성이높았으며 para-nitrophenyl-β-xylopyranoside에 대해낮은활성을보였다. Xyn10A의활성은 Cu2+, Mn2+과 SDS에의해서크게저해되었으며 K+, Ni2+과 Ca2+에의해 는상당하게증진되었다. 또한이효소는 xylobiose 보다중 합도가큰자일로올리고당을분해하였으며, 자일로올리고당 의최종가수분해산물은 xylose와 xylobiose로확인되었다.
Acknowledgments
This research is based on the support of 2019 Woosong University Academic Research Funding.
Conflict of Interest
The authors have no financial conflicts of interest to declare.
References
1. Sharma N, Sharma N. 2017. Microbial xylanases and their indus- trial applications as well as future perspectives: a review. Global J. Biol. Agric. Health Sci. 6: 5-12.
2. Valenzuela SV, Díaz P, Javier Pastor FI. 2010. Recombinant expression of an alkali stable GH10 xylanase from Paenibacillus Fig. 4. Thin-layer chromatogram of hydrolysis products of β-
1,4-linked xylooligosaccharides with purified PwXyn10A- CD. The reaction mixtures containing the purified xylanase and 1% xylooligosaccharides in 20 mM sodium phosphate buffer (pH 6.0) were incubated for 4 h at 45℃, respectively. Numbers 1 to 5 stand for xylose to xylopentaose in bottom and right side, respectively. Lanes 2E to 5E represent each hydrolyzate of xylo- biose to xylopentaose with purified PwXyn10A-CD.