Kor. J. AppL. MicrobioL. Bioen강
Vol.14, No.1, 51- 55! 1986)
Regulation of Cephalosporin C Acetyl-hydrolase in Cephalosporin C Fermentation by
Cephalosporium acremoη ium M- l13
Sang- Ho Choi
,
Jeong- KukLe
e, Y
oung- HeeKh
o and Tae-Ic
k MheenGenetic Engineering Center
,
KAIST,
Seou\,
Korea (Received October 22,
1985)Cephalosporium acremonium M-113 에 의한
Cephalosporin C 발효에 있어 서
Cephalosporin C Acetyl- hydrolase 의 조절
최상호·이정국·고영희·민태익
한국과학기술원 유전공학센터 (1 985년 10월 22일 수랴)
In the ferlllenlaliun uf cephaluspurin C (CPC) using a mutant strain, CephιJ!osporium ocremoniUlI/ M-113, CPC was converted inlo deacelyl cephalospurin C (DCPC) by cephalosporin C acetyl hydrolase (CAH). After complele consulllp- lion of glucüse in ferlllentaliün brolh, Ihe CAH aClivity appeared as CPC slarted 10 acculllulale and Ihen increased rapidly. The CAH activity was regulated by glucose concentralion and Ihe regulatüry Illechanism was not by inhibilion bul by repression. During Ihe CPC fermentation, cünlinuous feeding of glucose at low level into fermentaliün brolh cuuld regulate the CAH activily as weII as increase Ihe production of CPC by suppressing Ihe conversiun of CPC inlo DCPC
51
EarIy, Jeffery el 0/(1) found that either enzymatic or alkaline hydrolysis of CPC produced a compound wilh reduc- ed biulogical activity. This new compound, designaled deacetyl cephalospürin C (DCPC), was slrucluraIIy similiar to CPC, excepl Ihal Ihe acetoxyl group of CPC was replaced with a hydroxyl function.(2)
DCPC has been detected in both intraceIIular and ex- traceIIuar fluids of Cepholosporium spp. Huber el 01.(3) sug gesled that the origin of DCPC in CPC fermentatiün broths be produced by non enzymatic hydrolysis of CPC. They detected esterase activity in cell free extracts of C
“
crellloniulIl, but these extracts failed to deesterify CPCand linaIIy convert it to DCPC. J udging Irom the fermenta tion prolile 01 this mutant, it has been suggested that the strain may produce CAH. Then they have tried to detect the enzyme activity and isolated the enzyme from the mutant culture broth lor the lirst time. The lindings lead to the con- cIusion that the accumulation 01 DCPC by the mutant was made by the action 01 CAH
Fujisawa el 0/(4) have found that one of the CPC negative mutants accumulate CPC in the early stage of fermentation
Lately, Hinnen el 01.(5) also have shown Ihat under cer- tain conditions C ocremoniulll is capable 01 producing CAH They have purilied and characterized this enzyme. In addi- tion, evidence has been presented that the synthesis of CAH is regulated by the nature 01 the carbon sources.
In the present study, we have lound that CAH activity was repressed by glucose and tried to maintain the CPC pro-
52
Table 1. Composition of fermentation media
Synthetic medium (%) Complex medium (%)
Sucrose 3.6 Sucrose 2
Glucose 2. 7 Glucose 0.5
D. L-methionine O. 5 Peanut-meal 3 Methyloleate O. 2 Soybean-meal 3
(NH
‘),
SO‘
O. 75 DL-methionineKH
,
PO‘
1.5 CaCO,
K
,
HPO‘
2. 1 Borax 0.05Salt mixture* 10 Methyloleate O. 2
* Na
,
SO‘
0.75 g,
MgSO‘
0.18 g,
Fe (NH‘),
(SO‘),
'6H
,
O O. 15 g,
CaCI,
0.06 g,
MnSO‘
0.3 g,
Zn- SO‘
'7H,
O 0.03g,
CuSO.'5H,
O 0.0075g,
per 100 ml distilled water.duclivily al maximum level in lermenlalion brolh by regula.
lion 01 CAH aclivily
Materials and Methods
Microorganisms
A high produclive mulanl slrain, C acremonium M.113 which had been selecled among 30.000 colonies 01 mulagen lrealed C. ucremoflium A TCC 20339 was used lhroughoul lhis experimenl. Alcaligenes {uecalis ATCC 8750 was used as a lesl organism lor lhe delerminalion 01 CPC produced in cullure brolh
Media and culture conditions
CPC producing microorganism was Ireeze.dried in vials lor slorage. Composilion 01 lhe media used lor lermenlalion are described in Table 1. These media had been oplimized lor lhe maximum produclion 01 CPC by C. acremoniurn M.113 slrain. Flask experiment was carried out using I1 Erlenmeyer flask containing 200 ml 01 synthetic medium. In.
oculum size was 2% (v/v) and shaked at 280C and 250 rpm lor 8 days. 2.5 1 Jar lermentor (MARUBISHI, Japan) experi.
ment was also performed with complex medium. In this case. inoculum size was 10% (v/v) with aeration 01 2 vvm.
and agilation 01300 rpm lor 8 days. In order to determine the effecl 01 glucose concentralion on CAH activity, 100 ml Erlen.
myer f1ask cultures containing 20 ml complex medium withoul methyloleate and Borax were perlormed
Identification of fermentation products
The lermenlalion products were identilied by thin iayer chromalography (TLC). Cellulose plates (Eastman Kodak.
wilh f1uorescenl indicalor) were used wilh lollowing two
Kor. J. Appl. M icrobiol. Bioeng
alternative solvent systems by ascending techniques after sufficient saturation; upper phase 01 (A) n.butanol: acetic acid: water (3: 1: 1, v/v) or (B) n.propanol: water (7 :3. v씨 The chromatograms were air dried, and the position 01 antibiotic zones were conlirmed by U.V. light.(3.4.6)
Assay of antibiotics
CPC titers were determined by the agar dillusion cylinder method with A. (aecalis as test organism. Hydroxylamine method(7) was used lor chemical assay 01 total cephalo- sporins. This method is nol selective to cephalosporins. there- lore all CPC derivatives. eg. DCPC. Deacetoxy cephalosporin C. and CPC were determined as the same contrihlltions.
Measurement of CAH activity
CAH activity was measured by using HPLC according to the method suggesled by While el al.(8) The reaction mixture.
conlaining 200μI 01 96mM CPC, 100μI 01 pOlassium phosphate buffer(O.OIM, pH 7.0) and 900μI 01 lillrated broth as enzyme source, was incubated lor 4 hrs at 280C. After in.
cubation, lhe reaction was slopped by adding 1 ml 01 phenol.
chlorolorm solution. Then CPC and DCPC were assayed‘ En.
zyme aclivities were determined by subslracting amount 01 DCPC lormed 'in blank Irom thal lormed in sample per reac.
lion lime. One unit 01 this enzyme activity was delined as the amount which produce 1 μ mole 01 DCPC per minute under the assay condition‘
Determination of glucose concentration
Enzymatic analysis using peroxidase-glucose oxidase (PGO. Sigma) was applied to determine the glucose concen.
tration in the lermentalion broth.
Results and Discussion
Conversion of CPC into DCPC in fermentation broth byCAH
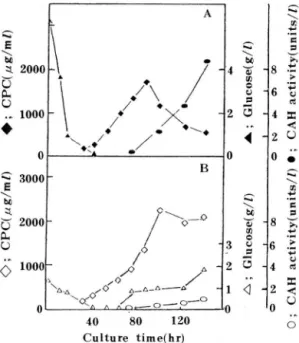
Fig. 1. showed the change 01 the lermentation variables in synthetic medium using I1 Ilask culture. The CPC syn.
thesis reached maximum level at 96 hrs and then decreased rapidly but total CPC determined by chemical assay was almosl constant. 11 would be thought lhat CPC was broken down to its derivatives. other cephalosporins. 50. lermenta.
tion procucts 01 late production phase were developed on TLC in order to identify the converted products 01 CPC. The RI values 01 standard and samples shown in Table 2. sug- gested that CPC should be converled into DCPC.
To understand the reason 01 this conversion whether by enzymatic or nonenzymatic reaction. lollowing experiment
53
빠
M
B
d y //
4t
n //
L mnu
e //
-m 때 때
//’
-oh
a
c//
•
2g 빼 쩌
·m
ljfv
m 뼈
m
t :
U E
l cu
(뜯n
S
며× 』@}QA}Q)gQEO
·m C
았
P b [ u E
--m
M FM페
없 빼 야 M 때
“…
많
m
1.0 2.0
2.0 4.0 6.0 Incubation time(hr) Vol.
14
, No. 1,s셈00 많00빠 0야「
훌흙웰짧헬%뻐규팩ol협꿇l 뤘 ~ §뤘 500빨 났; 싹치쓸 i5~
~ 01 (1 /r-까/
,-0I
81 등
‘i
’
i、、1야
’
~6
f
,;:l'il I V
잉
1 23
4000υ
’
"H
r.:> 。
lι
서 2
r
t 잉 ~20000'+
was perfurmed using dialized supernatent of precipitated ferlllelltatiull brüth by
80
% ammoniulll sulfate as enzyOle suurces (Fig. 2.). Under the standard assay conditiüns described above. the reaction proceeded linearly fur at least 4 hrs and DCPC furlllatiün was prupurtiundl tu enzyme cün.cé'llIratiulI as sllUwn in Fig. 2. Fronl these results it could be c(JIlcluded that cOllversion of CPC into DCPC was enzYOlatic reactiull Ilut mechanical ur chelllical reactiull
CUllversiull of CPC was alsu übserved ill ferrllelltatiün
pressiull uf CAH activity
Effect of glucose concentration on CAH activity Fig. 4. shüwed the effects üf initial glucose concentration üll CAH activity. The CAH activity was lüwer and appeared
l1lure lately with increasing concentration üf glucose. In case
:SNJ/l 옳
。。 ;찢 ι: 헬 1; £
Culture time(hr)
on the CPC productivity and CAH activity
CPC CAH
- 0- - . - -口 -.-
-@ -
뼈
M
째 때
(‘E\야X)Qιμ syslé'11l 싸 ilh cUIlIf> lex IllediullI as slluWIl ill Fig. J. Tlle CAH
a Cl I 、Ity 、이 as af>peared after glucuse def>letiull alld 1이Jidly ill creased Willl the decrease üf CPC alld SilllUltdllé'Uusly ill- crease uf DCPC. lhe allluullt üf tulal Ce f> llalusf>ürills, 11 이Never, did nüt challge su excessively. Illilial appearallce uf 5111all allluullt üf DCPC J1liglll be ré'sulled frUIII bY f> ruduclS ill Ille lasl slep of the biüsYlllhesis, tlI e cUllversiulI ul DC! 'C illlu C!)C, Ilül frülll Ilydrulysis uf CPC by CAH(2j ÏÎle cluse rela liUIISllip belweell glucuse CUllcelltratiull alld CAH activity
‘IruSl:' [Ile 니 ue5liul' wlletller glucuse Illight have effecl ull sup-
Table 2. Identification of fermentation products of C. acremonium 1\1-113 by TLC
R f value
Solvent system
B
*
Solvent system A AnlibioticsS ample
0.39
Standard
N.D Sample
0.43
Standard
** N. D.
Unknown (mav be pen N)
CPC
0
,33
0.28 0
,33
0
‘28 0
,32
0.19
0‘ 37O . 1 9
Glucose(%)
0 .5;
0; 1. 0: 2.0:DCPC
*
Solvent systems are described in materials and methods* *
Not determinedsynthesis. Therefore, if CAH activity was inhibited by glucose, CAH activity was to be detected after glucose deple- tion in both cycloheximide treated culture or not. CAH activi- ty, however, was not detected in cycloheximide added culture in which protein synthesis was inhibited after glucose depletion as shown in Fig. 5‘ From these results, we conclud- ed that CAH activity was regulated by glucose repression Regulation of CAH activity by continuous feeding of glucose to the fermentation system
Glucose consumption rate was determined during the CPC fermentation. It was 0.4-0.6g gIucose/l/hr at the Iog phase of growth and about 0.2g glucosel I/hr at the stationary phase. Therefore, 20% gIucose stock solution was fed con- tinuously at a rate of 0.2g glucosel //hr enough to compensate glucose consumption rate at production phase of CPC. This rate of glucose feeding was selected from the following aspects that CPC procuction might be repressed at higher rate and CAH activity might be depressed at lower rate even though CPC production could not be repressed‘
As shown in Fig. 6. CPC titer was increased by above 20% and was maintained relatively at constant level in glucoSE•fed fermentation system compared with fermenta- tion without glucose feeding. CAH activity in glucose fed fermentation was much lower than that in control fermenta tion. Therefore increase of CPC titer in glucose fed fermenta- tion might be the results 01 removal of glucose repression of CPC production and CPC hydrolysis by CAH to DCPC. In conclusion, continuous glucose feeding at a rate of 0.2g glucoselllhr was very effective to increase CPC production and to repress CAH activity
Kor. J. Appl. Microbiol. Bioeng
/박\펴4훌
a / 8 / - r
54
800
때 빼 째
(‘;I\뼈a)QAυ
160
Fig.5. Effect of cycloheximide on the CPC pro- ductivity and CAH activity
CPC Cycloheximide ( - ) ; - 0 - Cycloheximide (+) ; - • -
• Cycloheximide addition time f
-- A A
‘
「ν
--
40
01 2.0% glucose, CPC was maintained almost constantly eventhough its maximum titer was low due to catabolite repression of CPC production by glucose
To elucidate mechanism of glucose effect on CAH activi- ty, cycloheximide (1 OO,u냥Iml) was added to fermentation borth daily after 100 hr (•in Fig. 5.) at which glucose in lermentation broth was disappeared completely. Cyclohex- imide is known to antifungal agent which inhibits protein
。 t
..
Cephalosporium acremonium M -113 을 사용 찬 cephalosporin C (CPC) 발효에서 CPC 는 cephalos- porin C acetyl hydrolase (CAH) 에 의 해 서 deacetyl cephalosporin C (DCPC) 로 전환되었다.
CAH 활성도는 CPC 가 축적되기 시작하고 중의 포도당이 완전히 소비된 후에 냐타나서 히 증가하였다.
CAH 활성도는 포도당 농도에 의해서 조절되었우 며, 그 조절가작은 포도당에 의한 inbibition 이 아니 고 repression 임 이 확인되었다
CPC 딸효중 배양액에 포도당을 저농도로 계속 공급함으로써 CAH 활성도를 조절할 수 있었고、 그 결과 CPC 의 DCPC 로의 전환을 억제하여 CPC 생 산성을 증가시컴 수 있었다.
지 격 애 급 요
겉 3000
、、
b애
::t
L
r~、、" 。Ö
2000~ /ι
/
u .6
o
10001
.~ 거) \ ,.{'
o
‘^ _ /
ß-l'l\∞?: 2L-。一。-。
40 80 120
Culture time(hr)
Fig.6. CPC fermentation with or without con- tinuous glucose feeding
A ; Without glucose feeding
B ; With glucose feeding at rate of O. 2g/l/hr Medium Complex meclium without Borax and Me
thyloleate
(‘\mii:z)ki;;“어
ZJRQ
Z에Q ·(~\mι::i)‘s;;μiN ”
。” gu CU j*
9‘
nu
QO
Cu j*
9“
nv
‘\뼈)ωm-s-g
‘
”4
2
0 A
?/
\
.. ·k.
·/
/
/
--
//
•
/
•
、
‘
•
\
---η
‘ ---‘
nu
nu
m m
‘ (
E\ 빼
a
)Q
A}
Q
• “
‘\빼)QmRE-〔)
이 ” 3 -2
1 0 B 0
0
Vol.14, No.l,
References
1. Jeffery, J. D.A. , E.P. Abraham and G.G. F. Newton Biochem. J, 81
,
591 (1 961)‘2. Robert, B. Kammer: Chemislry and biology of,β -Laclam Anliblollcs, (Robert, B. Morin and Marin, Gorman ed.) Academic Press, Newyork, V이 3, 287 (1982)
3. Huber, F.M., R.H‘ Baltz and P.G. caltrider: Appl. Mlcro blol , 16(7), 101 I (1968)
4. Fujisawa, Y. , H‘ Shirafuji and T. Kanzaki: Agr. Blol.
55
Chem. , 39(6), 1303 (1975).
5‘ Hinnen, A. and J. Nuesch ’ Antimlcrob. Ag. Chemolher., 9(5), 824 (1976).
6. Fujisawa, Y., H. Shirafuji, M. Kida, K. Nara, M. Yoneda and T. Kanzaki: Agr. Biol. Chem., 39(6), 1295 (1975).
7. Mays, D.L., F.K. Bangert, W.
c.
Cantrell and W.G. Evans:Anal. Chem. , 47(1 3), 2229 (1 975)
8. White, E.R. and M. Fox‘J Anlibiol., 35(1 1), 1538 (1982).
9. Kennel, Y.M., C.J. Behmer and A.L. Demain: Enzyme Microb. Technol.