INTRODUCTION
Black raspberry (Rubus occidentalis L.) is a perennial sh-rub distribute in the eastern parts of North America. It has traditionally been used as a tea and medicine for treating stomach aches and bowel complaints by North-Amercian Indians. The black raspberry contains high levels of nu-trients and chemopreventive vitamins and phytochemicals, including beta-carotene, ellagic acid, ferulic acid, antho-cyanins, ellagitannins and coumaric acid (Harris et al. 2001;
Xue et al. 2001; Han et al. 2005). The biological activities of berries are largely related to the content of a diverse range of phenolic-type phytochemicals such as flavonoids (anthocyanins, flavonols, and flavanols), tannins [condens-ed (proanthocyanidins) and hydrolysable (ellagitannins and gallotannins)], stilbenoids (e.g., resveratrol), phenolic acids (hydroxybenzoic and hydroxycinnamic acid derivatives), and lignins (Perkins-Veazie and Nonnecke 1992; Seeram 2006).
The phenylpropanoid pathway provides a large number of biological important secondary metabolites such as ligins, flavonoids, hydroxycinnamic esters and coumarins. The secondary metabolites are closely associated with fruit
─ ─ 97 ──
Cloning and Characterization of Cinnamate-4-Hydroxylase Gene
from Rubus occidentalis L.
Eun Mi Lee, Seung Sik Lee, Byung Chull An, Shyamkumar Barampuram, Jae-Sung Kim, Jae-Young Cho1, In-Chul Lee2and Byung Yeoup Chung* Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute,
Jeongeup 580-185, Korea
1Department of Applied Life Sciences, Chonbuk National University, Jeonju 561-756, Korea
2Senior Industry Cluster Agency, Youngdong University, Chungbuk 370-701, Korea
Abstract -- Cinnamate-4-hydroxylase (C4H) is a key enzyme of phenylpropanoid pathway, which leads a variety of secondary metabolites to participate in differentiation and protection of plant against environmental stresses. In this study, we isolated a full-length cDNA of the C4H gene from a black raspberry (Rubus occidentalis L.), using a reverse transcriptase-PCR and rapid amplifi-cation of the cDNA ends (RACE)-PCR. The full-length cDNA of the RocC4H gene contained a 1,515 bp open reading frame (ORF) encoding a 504 amino acid protein with a calculated mole-cular weight of about 57.9 kDa and an isoelectric point (pI) value of 9.1. The genomic DNA analy-sis revealed that RocC4H gene had three exons and two introns. By multiple sequence alignment, RocC4H protein was highly homologous with other plant C4Hs, and the cytochrome P450-featur-ed motifs, such as the heme-binding domain, the T-containing binding pocket motif (AAIETT), the ERR triad, and the tetrapeptide (PPGP) hinge motif, were highly conserved. Southern blot analysis revealed that RocC4H is a single copy gene in R. occidentalis.
Key words : Black raspberry, Cinnamate-4-hydroxylase, Phenylpropanoid pathway, RACE, Southern blot
* Corresponding author: Byung Yeoup Chung, Tel. +82-63-570-3331, Fax. +82-63-570-3339, E-mail. [email protected]
quilities in addition to structural and defense-related func-tions. Ligins play a key role in plant mechanical support, solute conductance and physical barrier against pathogen. Various flavonoid compounds act as signaling molecules or antagonistic elements in defense and other physiological activities of plants (Weisshaar and Jenkins 1998; Dixon and Steele 1999).
Cinnamate-4-hydroxylase (C4H), a member of the cyto-chrome P450 monooxygenase superfamily, catalyzes the hydroxylation of trans-cinnamate to p-coumarate (4-hydro-xy trans-cinnamate). Most eukaryotic C4Hs are 50~60 kDa proteins and localize to the endoplasmic reticulum (ER) (Ro et al. 2001; Ro et al. 2002; Achnine et al. 2004). C4H is regulated in a pattern of temporal and spatial gene expression correlated with its role in lignifications (Bell-Lelong et al. 1997; Koopmann et al. 1999) and highly inducible by light, wounding, pathogen infection and elici-tor (Fahrendorf and Dixon 1993; Bell-Lelong et al. 1997; Mizutani et al. 1997). In addition, it was significantly induc-ed by gamma irradiation (Nagata et al. 2005). Many C4Hs have been isolated and characterized in herbal plants. How-ever, there are little reports about the isolation of C4H genes from woody plants. In woody plants, C4H is particularly important for the biosynthesis of lignin, the second most abundant biopolymer after cellulose, and C4H is likely to play a critical role in the ability of phenylpropanoid meta-bolism to channel carbon from primary metameta-bolism into the biosynthesis of lignin and other polymers in trees. To better understand the role of C4H in woody plants, we isolated and characterized the C4H gene from R. occidentalis. This report will provide the basis of further research for the role of the C4H gene in the phenylpropanoid metabolism in R. occidentalis.
MATERIALS AND METHODS
Plant material and growth conditionBlack raspberry (Rubus occidentalis) plants were grown in green house of Chonbuk National University (Jeonju, Korea) under normal photoperiod conditions. Leaves were immediately frozen in liquid nitrogen after harvesting and stored at -70�C until used.
Isolation of the full-length cDNA and genomic DNA of RocC4H
Total RNA was extracted from leaves of R. occidentalis using a cetyltrimethylammonium bromide (CTAB) method (Jaakola et al. 2001). To isolate the full-length cDNA of RocC4H, 5′- and 3′-RACE were performed using a gene-specific primer according to the SMARTTMRACE cDNA Amplification Kit (Clontech Laboratories, CA, USA) by following the manufacturers instruction. Two gene-specific primers, RocC4H-GSP1 (5 ′-CAGGACATGGTGTTCACC-GTCTACGG-3′; for 3′-RACE) and RocC4H-GSP2-2 (5′-TTGTTCGTGAAGAACGGGACCGTCATG-3′; for 5′-RACE), were designed from the conserved sequences of other C4H genes. RACE-PCR conditions were 5 cycles of 94�C for 30 sec, 72�C for 2 min, then 5 cycles of 94�C for 30 sec, 70�C for 30 sec, 72�C for 3 min, and 30 cycles of 94�C for 30 sec, 68�C for 30 sec, 72�C for 3 min. The amplified RACE-PCR products were purified, ligated into pGEM-T Easy vector (Promega, Madison, USA) and clon-ed into E. coli strain XL1-blue MRF′ followed by a sequen-cing. After comparing and aligning the sequences of the 5′ and 3′ RACE-PCR products, the cDNA of RocC4H was isolated from a reverse transcriptase (RT)-PCR. The cDNA library prepared by a Maxime RT premix kit (iNtRON BIOTECHNOLOGY, Seongnam, Korea) as a template. PCR was performed using Maxime PCR PreMix (i-MAX II) (iNtRON BIOTECHNOLOGY, Seongnam, Korea) with gene specific primers under the following condition: 94�C for 30 sec, 63�C for 30 sec, and 72�C for 2 min with 30 cycles. The primers were designed according to the known cDNA sequences from the RACE-PCR products. Sequ-ences of the primers were as follows: sense primer (Roc-C4H-F), 5′-ATGGATCTCCTCCTCATGGAGAAGA-3′; antisense primer (RocC4H-R), 5 ′-TTATGTCCTTGGCTT-CATGACTATGG-3′.
In order to isolate the genomic DNA of RocC4H, we ex-tracted the genomic DNA from the leaves by a modified CTAB method (Doyle and Doyle 1987), and then PCR was performed with the primers of RocC4H-F and RocC4H-R covering the coding region under the following condition: 94�C for 30 sec, 63�C for 30 sec, and 72�C for 3 min with 30 cycles. The PCR products were purified, cloned into a pGEM-T Easy vector (Promega, Madison, USA) and intro-duced into E. coli strain XL1-blue MRF′ by heat-shock
method. The cloned RocC4H gene was identified by sequ-encing.
Bioinformatic analysis
Sequence analyses were carried out on the website (http: //www.ncbi.nlm.nih.gov/, http://www.expasy.org/). Multi-ple sequence alignment of RocC4H and other plant C4Hs were performed using the Clustal W program (Thompson et al. 1997) from web sites (http://www.ebi.ac.uk/Tools/clu-stalw/index.html, http://www.genome.jp/). Phylogenetic tree was genertated by the neighbour-joining method (Ki-mura 1980; Saitou and Nei 1987) using the Clustal W pro-gram. Network Protein Analysis server (NPSA) (http:// npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/ npsa_server.html) was used for the secondary structure prediction.
Southern blot analysis
Genomic DNA was extracted from the leaves by a modi-fied CTAB method (Doyle and Doyle 1987). Genomic DNA (10μg) was digested for overnight at 37�C with EcoR I, Hind III, Nhe I and Xba I (Takara, Shiga, Japan) respec-tively. The digested DNA was separated by 0.8% agarose gel electrophoresis and then transferred onto a positively charged nylon membrane (Zeta-Probe blotting membrane; Bio-Rad, CA, USA) by a capillary transfer (Sambrook and Russell 2001). The blot was immobilized with a UV cross-linker. The blot was hybridized with [32P]-labeled RocC4H
cDNA probes by a random primer DNA labeling kit Ver. 2 (Takara, Otsu, Japan). Prehybridization and hybridization reactions were carried out in church buffer (Church and Gilbert, 1984) [0.5 mM Na2HPO4, pH 7.2, 1 mM EDTA,
1% (w/v) BSA, 7% (w/v) sodium dodecyl sulfate (SDS)] at 65�C. After hybridization, the blot was washed twice with 2
×SSC (1×is 150 mM NaCl, 15 mM sodium citrate), 0.1%
SDS at for 20 min at 65�C, and then washed twice with 1×
SSC, 0.1% SDS at for 10 min at room temperature. The blots were then exposed to X-ray film for appropriate times at -70�C.
RESULTS AND DISCUSSION
Cloning and characterization of RocC4H
The full-length cDNA sequence of the RocC4H gene was
obtained using 3′- and 5′-RACE-PCR. It was 1,744-bp long. There was 54-bp of the 5′ untranslated region (UTR) and 175-bp of the 3′ UTR including a poly (A) tail. Sequence analysis of the cDNA show that it contains a 1,515-bp ORF (including stop codon, TAA) which encodes a polypeptide of 504 amino acids with a estimated molecular weight of 57.9 kDa and an isoelectric point (pI) value of 9.1 (using the software of Compute pI/Mw tool at http://www.expasy.org).
55ATG57 was proposed to be the initial start codon which
has the start signal (A/GXXATGG) for effective translation for eukaryotic genes (Kozak 1984) (Fig. 1a).
Specific primers derived from the start and stop regions of the cDNA had been designed for PCR to obtain the genomic RocC4H gene. A 3,278 bp of genomic RocC4H containing the start and stop codon was obtained. By the comparison of the genomic DNA and cDNA sequence, genomic RocC4H gene contains three exons and two introns (Fig. 1). All of the splicing site sequences are in accordance with GU/AG consensus rule.
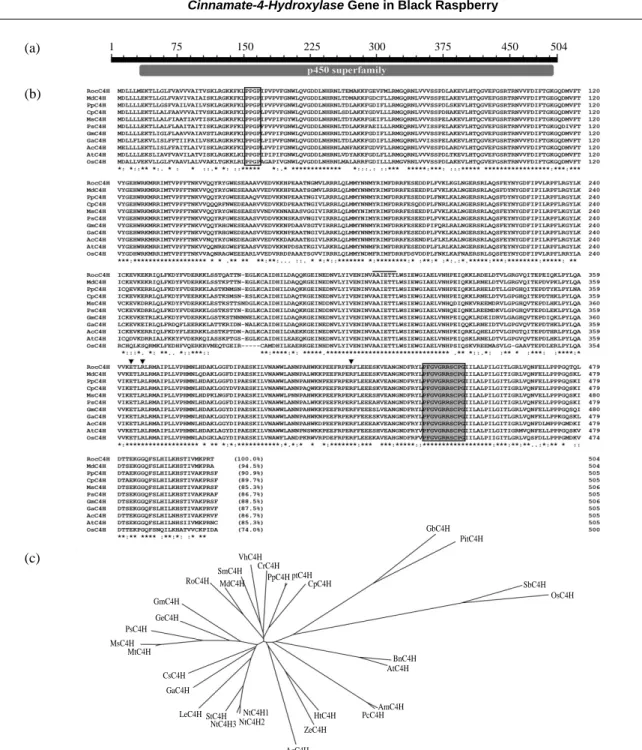
By the web-based database search with Conserved Do-mains (at http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb. cgi), multi-alignment and phylogenetic analysis, it was re-vealed that the RocC4H belongs to the cytochrome P450 superfamily (Fig. 2). A deduced amino acid sequence of the RocC4H protein had considerable high sequence homology with those of C4H in other plant. For example, RocC4H shows identities of 94.5% to Malus×domestica, 90.9% to
Populus trichocarpa×Populus deltoids, 89.7% to Citrus×
paradisi, 85.3% to Medicago sativa, 86.7% to Pisum sati-vum, 88.5% to Glycine max, 87.5% to Gossypium arbore-tum, 86.7% to Allium cepa, 85.3% to Arabidopsis thaliana, and 74% to Oryza sativa. RocC4H exhibits the characteri-stics of cytochrome P450 gene family members (Fig. 2), such as the heme-iron binding region PFGVGRRSCPG, PPGP tetrapeptide near the N-terminus, the T-containing binding pocket motif (AAIETT) and the E363-R366-R420triad
(Chapple 1998; Chen et al. 2007). The conserved PPGP tetrapeptide is necessary for proper heme-incorporation and P450 protein stability (Szczesna-Skorupa et al. 1993). The central part of the heme-iron domain (gray boxed) contains a proline residue (Pro448) which is essential for
heme-pro-tein interactions in plant C4Hs (Schalk et al. 1999). To obtain insight into the secondary structure of RocC4H, we conducted secondary structure prediction for the pri-mary sequences using NPSA server. The secondary
struc-tures of RocC4H are composed of alpha helices (48.41%, 244 aa), extended strands (6.94%, 35 aa), random coils (42.06%, 212 aa) and ambiguous states (2.58%, 13) (Fig. 3). Alpha helices mainly disperse at the middle region and the N-terminus and connected by random coils. Extended strands mainly distribute at two regions: N-terminal region
(between second alpha helix and forth alpha-helix) and C-terminal region (~110-residue C-terminal region), but
there are not found in the middle regions, predominantly alpha helices region.
On the basis of sequence alignment, phylogenetic tree and secondary structure analyses, RocC4H may function as Fig. 1. Sequence and organization of the RocC4H gene. (a) Genomic DNA sequence and deduced amino acid sequence of RocC4H. The stop codon (TAA) is indicated as an asterisk in the sequence. Lower case letters represent UTR. Capital letters represent translated region. Hind III and Nhe I restriction endonuclease site is indicated by an underline and double-lined respectively. Polyadenylation signal sequ-ence (AATAA) is boxed. The nucleotides (upper line) and amino acid residues (lower line, a parenthesized passage) are numbered on the right. (b) Schematic representation of the genomic DNA orgnization of RocC4H. Positions of the exons (black boxes) and introns (white boxes) were determined by comparing the genomic DNA sequence with the cDNAs of RocC4H. UTR is indicated in gray.
(a) (b) 2218 Hind III 2407 Nhe I 2737 1641 1507 840 155
5-UTR Exon 1 Intron 1 Exon 2 Intron 2 Exon 3 3-UTR
Fig. 2. Multiple sequence alignment and phylogenetic relationship of C4H proteins. (a) The cytochrome P450 conserved domain. (b) Com-parison of the deduced amino acid sequeneces of RocC4H with C4Hs from other plants. Multiple alignments were performed using Clustal W. Amino acid residues are numbered at the right side, and the percentage of the number of identical amino acids shared with RocC4H is shown in parentheses at the end of the last lines. The Pro-rich domain (PPGP) is boxed. ERR tried is indicated by reverse triangle. The heme-binding domain (P-F-G/S-X-G-X-R/H-C-X-G/A/D) is gray boxed. The T-contain binding pocket motif (AAIETT) is lined. (c) Phylogenetic analysis of RocC4H and other C4H proteins. A phylogenic tree was constructed by the neighbour-joining method using the Clustal W program. GenBank and Swiss-Prot accession number of the proteins are as follows: MdC4H (Malus×
domestica, AAY87450), NtC4H1 (Nicotiana tabacum, ABC69411), NtC4H2 (Nicotiana tabacum, ABC69413), NtC4H3 (Nicotiana tabacum, ABC69412), SmC4H (Salvia miltiorrhiza, ABN48717), GmC4H (Glycine max, Q42797), LeC4H (Lithospermum erythror-hizon, BAB71716), StC4H (Solanum tuberosum, ABC69046), HtC4H (Helianthus tuberosus, Q04468), PpC4H (Populus trichocarpa
×Populus deltoids, AAG50231), GaC4H (Gossypium arboretum, AAG10197), VhC4H (Verbena×hybrida, BAE72875), CrC4H
(Catharanthus roseus, P48522), ZeC4H (Zinnia elegans, Q43240), AcC4H (Allium cepa, AAS48416), AmC4H (Ammi majus, AAO62904), PcC4H (Petroselinum crispum, Q43033), GeC4H (Glycyrrhiza echinata, Q96423), PtC4H (Populus tremuloides, ABF-69102), BnC4H (Brassica napus, ABF17873), MtC4H (Medicago truncatula, ABC59087), CpC4H (Citrus×paradisi, AAK57011),
CsC4H (Cucumis sativus, CAK95273), MsC4H (Medicago sativa, P37114), AtC4H (Arabidopsis thaliana, AAB58355), PsC4H (Pi-sum sativum, Q43067), GbC4H (Ginkgo biloba, AAW70021), PitC4H (Pinus taeda, AAD23378), SbC4H (Sorghum bicolor, AAK54447), OsC4H (Oryza sativa, NP_001055190).
(b) (c) (a) 1 75 150 225 300 375 450 504 VhC4H CrC4H PpC4H ptC4H GbC4H PitC4H SbC4H OsC4H BnC4H AtC4H AmC4H PcC4H HtC4H ZeC4H AcC4H NtC4H1 NtC4H2 NtC4H3 StC4H LeC4H GaC4H CsC4H MtC4H MsC4H PsC4H GeC4H GmC4H RoC4H MdC4H SmC4H CpC4H
a real C4H gene.
Southern blot analysis
In order to identify the copy number of C4H genes in the genome of R. occidentalis, genomic DNA was isolated and aliquots of genomic DNA (10μg/sample) were restricted with EcoR I, Hind III, Nhe I and Xba I respectively. EcoR I and Xba I recognition site did not exist within the genomic sequences of RocC4H, but single Hind III and Nhe I reco-gnition site exist within the genomic sequences, positioned at intron2 (Fig. 1a, b). The full-length RocC4H cDNA was used as a probe. Two bands can be observed in the lanes where DNA was digested with Hind III and Nhe I, and a
single hybridization signal was detected in the EcoR I and Xba I digested fragment lanes (Fig. 4). This result suggests that C4H exist as a single gene in the R. occidentalis. Like a RocC4H, the C4H genes were previously reported to have only single copy from various plant species, such as pea (Frank et al. 1996), Arabidopsis (Bell-Lelong et al. 1997; Mizutani et al. 1997), Parsely (Koopmann et al. 1999) and Ammi majus (Hubner et al. 2003), whereas C4H genes from alfalfa (Fahrendorf and Dixon 1993), periwinkle (Hotze et al. 1995), hybrid poplar (Ro et al. 2002) and Valencia orange (Betz et al. 2001) exist as multiple copies.
CONCLUSION
In this report, we firstly reported on the isolation and characterization of the full-length cDNA and genomic DNA sequences of C4H from Rubus occidentalis. The full-length cDNA of RocC4H has a 1,515-bp ORF (including stop codon) and the sequence encodes a polypeptide of 504 amino acid residues, with a calculated molecular weight of 57.9 kDa and an isoelectric point (pI) value of 9.1. The deduced amino acid sequences of the RocC4H protein had high identities with other C4Hs. They possess a conserved cytochrome P450 domain and all cytochrome P450-fea-tured motifs. RocC4H exists as a single copy in R. occiden-talis. This report will be helpful to carry out further research for the role of the C4H in the phenylpropanoid metabolism in black raspberry
ACKNOWLEDGEMENT
This study was carried out with the support of “Site Joint Cooperating Agricultural Research-promoting Project (Project No. 2007040108003702)”, RDA, and in part by Fig. 3. Secondary structure prediction of RocC4H. Construction of the consensus based on 3 different methods of prediction was conducted at NPSA structure prediction server (see information on the methods at npsa-pbil.ibcp.fr). Four kinds of line bars in descending in length represent alpha helix, extended strand, random coil and ambiguous states respectively.
100 200 300 400 500
Fig. 4. Southern blot analysis. Genomic DNA of R. occidentalis was extracted as described in MATERIALS AND METHODS. Genomic DNA (10μg ml-1) was respectively digested with
EcoR I (lane E), Hind III (lane H), Nhe I (lane N) and Xba I (lane X) followed by hybridization with [32P]-labeled
Roc-C4H probe. The sizes of the molecular weight markers are indicated in kb on the left side.
E H N X (kb) 10 8 6 4 2 RocC4H
the Nuclear R & D Program of the Ministry of Science and Technology, Republic of Korea.
REFERENCES
Achnine L, Blancaflor EB, Rasmussen S and Dixon RA. 2004. Colocalization of L-phenylalanine ammonia-lyase and cinnamate 4-hydroxylase for metabolic channeling in phenylpropanoid biosynthesis. Plant Cell 16:3098-3109. Bell-Lelong DA, Cusumano JC, Meyer K and Chapple C.
1997. Cinnamate-4-hydroxylase expression in Arabidopsis. Regulation in response to development and the environ-ment. Plant Physiol. 113:729-738.
Betz C, McCollum TG and Mayer RT. 2001. Differential expression of two cinnamate 4-hydroxylase genes in ‘Val-encia’ orange (Citrus sinensis Osbeck). Plant Mol. Biol. 46:741-748.
Chapple C. 1998. Molecular-Genetic analysis of plant cyto-chrome P450-dependent monooxygenases. Annu. Rev. Plant Biol. 49:311-343.
Chen AH, Chai YR, Li JN and Chen L. 2007. Molecular clon-ing of two genes encodclon-ing cinnamate 4-hydroxylase (C4H) from oilseed rape (Brassica napus). J. Biochem. Mol. Biol. 40:247-260.
Church GM and Gilbert W. 1984. Genomic sequencing. Proc. Natl. Acad. Sci. U. S. A. 81:1991-1995.
Dixon RA and Steele CL. 1999. Flavonoids and isoflavonoids -a gold mine for metabolic engineering. Trends Plant Sci. 4:394-400.
Doyle JJ and Doyle JL. 1987. A rapid DNA isolation proce-dure for small quantities of fresh leaf tissue. Phytochem. Bull. 19:11-15.
Fahrendorf T and Dixon RA. 1993. Stress responses in alfalfa (Medicago sativa L.). XVIII: Molecular cloning and expres-sion of the elicitor-inducible cinnamic acid 4-hydroxylase cytochrome P450. Arch. Biochem. Biophys. 305:509-515. Frank MR, Deyneka JM and Schuler MA. 1996. Cloning of
wound-induced cytochrome P450 monooxygenases ex-pressed in pea. Plant Physiol. 110:1035-1046.
Han C, Ding H, Casto B, Stoner GD and D’Ambrosio SM. 2005. Inhibition of the growth of premalignant and malig-nant human oral cell lines by extracts and components of black raspberries. Nutr. Cancer 51:207-217.
Harris GK, Gupta A, Nines RG, Kresty LA, Habib SG, Frankel WL, LaPerle K, Gallaher DD, Schwartz SJ and Stoner GD. 2001. Effects of lyophilized black raspberries on azoxy-methane-induced colon cancer and 8-hydroxy-2 ′-deoxy-guanosine levels in the Fischer 344 rat. Nutr. Cancer 40:
125-133.
Hotze M, Schroder G and Schroder J. 1995. Cinnamate 4-hydroxylase from Catharanthus roseus, and a strategy for the functional expression of plant cytochrome P450 pro-teins as translational fusions with P450 reductase in Esche-richia coli. FEBS Lett. 374:345-350.
Hubner S, Hehmann M, Schreiner S, Martens S, Lukacin R and Matern U. 2003. Functional expression of cinnamate 4-hydroxylase from Ammi majus L. Phytochemistry 64: 445-452.
Jaakola L, Pirttila AM, Halonen M and Hohtola A. 2001. Isola-tion of high quality RNA from bilberry (Vaccinium myrtil-lus L.) fruit. Mol. Biotechnol. 19:201-203.
Kimura M. 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 16:111-120.
Koopmann E, Logemann E and Hahlbrock K. 1999. Regula-tion and funcRegula-tional expression of cinnamate 4-hydroxylase from parsley. Plant Physiol. 119:49-56.
Kozak M. 1984. Compilation and analysis of sequences up-stream from the translational start site in eukaryotic mRNAs. Nucleic Acids Res. 12:857-872.
Mizutani M, Ohta D and Sato R. 1997. Isolation of a cDNA and a genomic clone encoding cinnamate 4-hydroxylase from Arabidopsis and its expression manner in planta. Plant Physiol. 113:755-763.
Nagata T, Yamada H, Du Z, Todoriki S and Kikuchi S. 2005. Microarray analysis of genes that respond to gamma-irra-diation in Arabidopsis. J. Agric. Food Chem. 53:1022-1030. Perkins-Veazie P and Nonnecke G. 1992. Physiological
chan-ges during ripening of raspberry fruits. HortScience 27: 331-333.
Ro DK, Ehlting J and Douglas CJ. 2002. Cloning, functional expression, and subcellular localization of multiple NADPH-cytochrome P450 reductases from hybrid poplar. Plant Physiol. 130:1837-1851.
Ro DK, Mah N, Ellis BE and Douglas CJ. 2001. Functional characterization and subcellular localization of poplar (Po-pulus trichocarpa×Populus deltoides) cinnamate
4-hydro-xylase. Plant Physiol. 126:317-329.
Saitou N and Nei M. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4:406-425.
Sambrook J and Russell DW. 2001. Molecular cloning: a laboratory manual, 3rd edition. Cold Spring Harbor Labora-tory Press, New York, USA.
Schalk M, Nedelkina S, Schoch G, Batard Y and Werck-Reic-hhart D. 1999. Role of unusual amino acid residues in the proximal and distal heme regions of a plant P450, CYP 73A1. Biochemistry 38:6093-6103.
Seeram NP. 2006. Berries. Academic Press, London, UK. Szczesna-Skorupa E, Straub P and Kemper B. 1993. Deletion
of a conserved tetrapeptide, PPGP, in P450 2C2 results in loss of enzymatic activity without a change in its cellular location. Arch. Biochem. Biophys. 304:170-175.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F and Hig-gins DG. 1997. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25:4876-4882.
Weisshaar B and Jenkins GI. 1998. Phenylpropanoid bio-synthesis and its regulation. Curr. Opin. Plant Biol. 1:251-257.
Xue H, Aziz RM, Sun N, Cassady JM, Kamendulis LM, Xu Y, Stoner GD and Klaunig JE. 2001. Inhibition of cellular transformation by berry extracts. Carcinogenesis 22:351-356.
Manuscript Received: June 27, 2008 Revision Accepted: July 30, 2008