INTRODUCTION
Escherichia coli (E. coli) is regarded as an attractive organ-ism for biohydrogen production through dark and anaerobic fermentation since it is fast-growing, nonsporulating, and well-characterized in physiological and biochemical aspects (Redwood et al. 2008). Formate is the sole precursor of molecular hydrogen in E. coli (Ordal and Halvorson, 1939), being converted into CO2and H2by the formate hydrogen-lyase complex (FHL) (Sawers 2005). The FHL enzyme com-plex comprises seven proteins, HycB, C, D, E, F, G and
FDH-H, which are encoded by the hyc operon and the fdhF gene. Among these polypeptides, the HycE is one of three NiFe-hydrogenases in E. coli and is also referred to as Hyd-3 or hydrogenase 3 (Sawers 2005). Functional assembly and maturation of Hyd-3 necessitates HypA-E and HypF proteins, which are encoded by the hypA-E operon and the hypF gene (Magalon et al. 2001; Blokesch et al. 2002).
Unlike E. coli producing biohydrogen through the dark fermentation, green algae and cyanobacteria can also synthe-size and evolve molecular hydrogen by direct and indirect biophotolysis systems through photosynthesis (Levin et al. 2004). Bidirectional hydrogenase is the major enzyme for biohydrogen production in green algae and cyanobacteria. A unicellular cyanobacterium Synechocystis sp. strain PCC
─ ─ 107 ──
Dissecting the Hydrogenase Expression and Activity of
Transformed Escherichia coli with the Bidirectional
NiFe-hydrogenase from Synechocystis sp. PCC 6803
Yu Ran Moon, Min Hee Lee, Byung Chull An, Byung Yeoup Chung, Jae-Sung Kim,Youn-Il Park1, Cha Soon Kim2and Jin-Hong Kim* Advanced Radiation Research Institute, Korea Atomic Energy Research Institute,
Jeongeup 580-185, Korea
1Department of Biology, Chungnam National University, Daejeon 305-764, Korea 2Radiation Health Research Institute, Korea Hydro & Nuclear Power Co., LTD,
Seoul 132-703, Korea
Abstract-- Synechocystis bidirectional hydrogenase genes (hoxEFUYH) and their putative promo-ter regions were isolated and transformed into E. coli. The hox genes were transcribed in the E. coli cells carrying the vector construct of pCC1FOS::phox::hox or pCC1FOS::pT7::hox and trans-lated into HoxEFUYH proteins, suggesting that the putative hox promoter can be constitutively activated in E. coli. Accordingly, the total hydrogenase activity was markedly increased up to 192% or 169% in the transformed cells, while the hydrogen uptake was decreased up to about 30% of the negative control. Although the gene expression of LexA, the only one transcription regulator pro-ven for the hox genes, was substantially decreased in the pCC1FOS::phox::hox cells after γγ-irradia-tion of 30 Gy, the expression levels of HoxEFUYH proteins were not altered significantly. Thus, it is also suggested that other transcription regulators as well as LexA might contribute to activa-tion of the hox promoter in E. coli.
Key words : Biohydrogen, Bidirectional hydrogenase, Escherichia coli, gamma rays, Synechocystis
* Corresponding authors: Jin-Hong Kim, Tel. +82-63-570-3333, Fax. +82-63-570-3339, E-mail. [email protected]
which is encoded by the hox operon (hoxEFUYH genes) and post-translationally modified by the HypA-F and HoxW pro-teins (Tamagnini et al. 2007).
Since the bidirectional hydrogenase has higher enzymatic activity than the hydrogenase 3 (Maeda et al. 2007), E. coli bearing the hox genes can exert more for biohydrogen pro-duction under anaerobic conditions. However, due to differ-ences in the biohydrogen production system between two organisms, transformation of cyanobacterial hox genes in E.
coli for biohydrogen production is challenged by a few key
questions as follows: 1) HycE modifying enzymes can work on post-translational modification of HoxEFUYH for func-tional maturation? 2) cyanobacterial bidirecfunc-tional hydroge-nase can utilize in vivo redox equivalents in E. coli? And 3) the putative hox promoter, which has proved to be regulated by a transcription factor, LexA, can exert on transcription of the hox genes for control of biohydrogen production in
E. coli? Although these questions remain to be addressed, it
was reveled by a recent report that HoxUYH, part of the bidirectional hydrogenase, induced more production of bio-hydrogen in E. coli by repressing the innate enzyme activi-ties of uptake hydrogenases, Hyd-1 and -2 (Maeda et al. 2007).
In the present work, we have isolated the full-length
hox-EFUYH genes from Synechocystis sp. strain PCC 6803 and
transformed an E. coli strain KRX with pCC1FOS vectors carrying T7 promoter/hoxEFUYH and hox promoter/
hoxEFUYH. Experimental evidence confirms that
cyanobac-terial bidirectional NiFe-hydrogenase could acquire function-ality through the assembly and maturation system of E. coli for the innate hydrogenase. Additionally, transcriptional activity of the cyanobacterial hox promoter in E. coli is first demonstrated and discussed.
MATERIALS AND METHODS
1. Bacterial strains, growth conditions, and
γγ-irradiation
Escherichia coli (E. coli KRX, a derivative of E. coli strain
K12) cells containing pCC1FOS::pT7, pCC1FOS::pT7::hox, or pCC1FOS::phox::hox, were initially streaked from -80 �C glycerol stocks on Luria-Bertani (LB) agar plates
(Sam-cultured from a single colony on the LB agar plates in fresh LB medium at 37�C with shaking at 250 rpm. Overnight, aerobic cultures (25 ml) were used to inoculate 75 ml of LB medium (with 0.2% (w/v) rhamnose) in 250-ml Erlenmeyer flasks, and these cultures were sparged for 5 min with nitro-gen gas, sealed, and incubated anaerobically at 37�C for 6 h with shaking at 250 rpm (Maeda et al. 2007). Alternatively, the growth curves of the cells were obtained by measuring the increase in the OD600of seed cultures (the initial value 0.05) freshly prepared from the overnight cultures. γ-Irradia-tion of the cells (OD600==0.5) was carried out at a dose rate of 2 or 30 Gy for 10 min using a 60Co gamma irradiator (IR-222, MDS Nordion Inc., Kanata, Canada) in the Advanced Radiation Technology Institute (ARTI).
For isolation of Synechocystis sp. PCC 6803 genomic DNA (Porter 1988), Synechocystis cells were cultured in BG-11 liquid medium (Rippka et al. 1979) at 28�C and 50μmol m-2s-1fluorescent white light with shaking at 120 rpm.
2. Constructing pCC1FOS::pT7::hox and pCC1FOS::phox::hox
The 6495-bp chromosomal sequences encoding hox-EFUYH were amplified into three fragments from the geno-mic DNA of Synechocystis sp. PCC 6803 using the Maxi-me™ PCR PreMix kit (iNtRON Biotechnology, Sungnam, Korea) and specific primers as follows: hoxEcoRI sense/anti-sense primers, 5 ′-CCGGAATTCATGACCGTTGCCACC-GATCGC-3 ′/5′-TGTTCGTAGCCCCCTAGGGCGATGT-ATTCA-3′; hoxAvrII sense/antisense, 5′-TGAATACATC-GCCCTAGGGGGCTACGAACA-3 ′/5′-TGATTGGTCTT-GTTTAAACGCCACAAAACA-3′; hoxPmeI sense, 5′-TG-TTTTGTGGCGTTTAAACAAGACCAATCA-3′; and hoxHindIII antisense, 5 ′-CCCAAGCTTTTAATCCCGCT-GGATGGACTT-3′. The PCR conditions are 94�C for 2 min followed by 40 cycles at 94�C for 30 s, 45�C for 45 s, and 72�C for 4 min. The PCR products were cloned into the pGEMT-easy vector (Promega, Madison, WI, USA) after digested with different restriction enzymes EcoRI, PstI/PmeI and AvrII/HindIII. Then, the full-length sequences of hox-EFUYH were sub-cloned and merged into the pCC1FOS vector.
The putative hox promoter region up to 540 bp upstream from the hoxE (Oliveira and Lindblad 2005), was amplified
from the genomic DNA of Synechocystis sp. PCC 6803
using the primers, hoxPro-F 5
′-GTGCACCAGCTCGGTT-TTACCTCTCTT-3′ and hoxPro-R
5′-CCGGAATTCATC-CTCAAAAAGGAGCAATAC-3′. The PCR conditions are
94�C for 2 min followed by 40 cycles at 94�C for 20 s, 45�C
for 30 s, and 72�C for 4 min. In contrast, the T7 promoter
was obtained from the pET28α vector plasmid DNA using
the primers, T7Pro-F 5
′-GTGCACCACATTCACCACCC-TGAATTG-3′ and T7Pro-R
5′-CCGGAATTCGCTGCTG-CCCATGGTATA-3′.
All the sequences of hoxEFUYH genes, hox, and T7 pro-moters cloned into the respective pCC1FOS vector were identified by sequencing and finally verified after merged into the pCC1FOS::pT7::hox and pCC1FOS::phox::hox.
3. RNA extraction, reverse transcription (RT)-PCR and SDS-PAGE analysis
Total RNA was extracted from E. coli cell cultures using RNeasy Mini Kit (Qiagen, Chatsworth, CA, USA) according to the supplier’s recommendation. For complete DNA re-moval, RNase-Free DNase Set (Qiagen, Chatsworth, CA, USA) provided efficient on-column digestion of DNA during RNA purification. One microgram total RNA was reverse-transcribed in an RT system, Maxime™ RT PreMix
(iNt-RON Biotechnology, Sungnam, Korea) for 60 min at 45�C
and 5 min at 95�C. Gene-specific sense/antisense primers
are 5
′-TGTTAGACGCCACCATGAAG-3′/5′-CCAAGCA-GACCACACAGGTA-3′ for hoxE,
5′-ACCGGGGATAA-GTTACTGG-3′/5′-CTAGCTAAATCGGGGTCAG-3′ for
hoxH, 5
′-GTGTCGAAACCAGCTCTCT-3′/5′-CTGTAA-ACCGTCGTCTGC-3′ for hypA,
5′-CAGAGTAATTCTG-CCACTCC-3′/5′-CTCGACGGTATCGGTATG-3′ for hypF,
5
′-GTTCTACCTCCAGCAGAATG-3′/5′-CCGACTACC-GTCTGTTCTAC-3′ for hycE,
5′-GTTGCAGGAAGAGG-AAGAAG-3′/5′-CCGTTACGTACATCCTGAGT-3′ for
lexA, and 5
′-ACTGGAACTGAGACACG-3′/5′-TCAGAT-GCAGTTCCCAG-3′ for 16srRNA. All the primers were
designed from mRNA sequences of the respective gene on the NCBI site. The 16srRNA was used as a control gene to normalize for differences in the amounts of total RNA. The
subsequent PCR was carried out with 1μl of a total 20 μl
RT reaction mixture in a PCR system, Maxime™ PCR Pre-Mix (iNtRON Biotechnology, Sungnam, Korea). The PCR conditions are 94�C for 2 min followed by 30 cycles of 94�C
for 20 s, 55�C for 10 s, and 72�C for 30 s. The resultant RT-PCR products were electrophoresed and analyzed on a 1.0 % (w/v) agarose gel containing ethidium bromide (EtBr).
For SDS-PAGE, the cell cultures, which were normalized
to the OD600of 0.5~0.8, were harvested by centrifugation
at 16,600 g for 1 min and solubilized for 5 min in a sample buffer, containing 24 mM Tris-HCl, pH 6.8, 20% (v/v) gly-cerol, 0.8% (w/v) sodium dodecyl sulfate, 5.8 mM 2-merca-ptoethanol, and 0.4% (w/v) bromophenol blue. Then, the solubilized proteins were applied onto a 12% polyacrylamide denaturing gel for electrophoresis and finally stained with Coomassie Brilliant Blue R-250.
4. Hydrogen production and uptake assay
Hydrogen production of cell suspensions taken directly from anaerobic cultures of transformed E. coli was measured as previously described by Appel et al. (2000) using a liquid phase Clark-type electrode unit (DW2) and an electrode con-trol unit (Oxylab system, Hansatech Inst., Norfolk, UK). The
electrode was oppositely polarized at a voltage of -700 mV
by inversely connecting to the control unit. Hydrogenase activity was expressed as the rate of hydrogen evolution from the cell suspension incubated with 5 mM methylviologen
(MV) and 10 mM sodium dithionite at 25�C.
Hydrogen uptake of cell suspensions was measured as the
increase in absorbance as oxidized MV (ε604==13.9 mM-1
cm-1) is reduced [MV2++ + +1/2 H2→ MV++ + +H++ ] (Maeda et
al. 2007). Oxidized MV solution was sparged first with
nitrogen gas for 10 min, was poured into cuvettes that were sealed with rubber stoppers, and was sparged with pure hydrogen gas for 10 min. Whole cell suspensions (0.5 ml) were mixed into the cuvettes and the change in absorbance at 604 nm during 5 min was monitored using a double beam UV/Vis spectrophotometer (UVICON 923, Bio-tek Kontron Inst., Winooski, VM).
RESULTS AND DISCUSSION
1. Expression of hoxEFUYH genes and operation of hox promoter in E. coli
To introduce cyanobacterial hydrogenase genes into the well-characterized organism E. coli for biohydrogen produc-tion through dark and anaerobic fermentaproduc-tion, the 6495-bp
hydrogenase locus of hoxEFUYH genes and the upstream 540-bp putative hox promoter region were isolated from
Synechocystis sp. strain PCC 6803 (Fig. 1B). The hox genes
were cloned with the T7 or hox promoter into the pCC1FOS vector, constituting the pCC1FOS::pT7::hox and pCC1FOS ::phox::hox constructs (Fig. 1C).
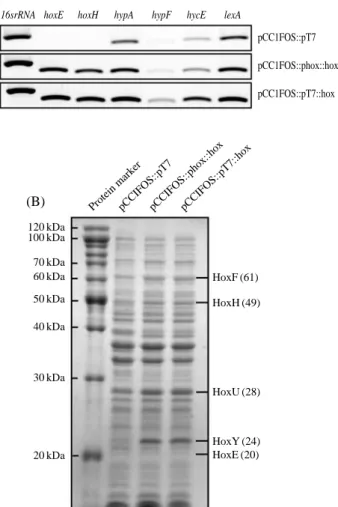
Expressions of the introduced hox genes were clearly iden-tified in both the E. coli pCC1FOS::pT7::hox and pCC1-FOS::phox::hox cells, as shown by the transcriptional analy-sis of hoxE and hoxH (Fig. 2A). The putative hox promoter region has been recently characterized in the Synechocystis strain (Oliveira and Lindblad 2005; Gutekunst et al. 2005). A transcription regulator, LexA, is involved in activation of the promoter. Although in vivo experiments showed that E.
coli promoters can be efficient in cyanobacteria (Li and
Golden 1993) and that some cyanobacterial promoters are transcribed in E. coli (Mazouni et al. 2003), in vitro experi-ments revealed that cyanobacterial RNA polymerases could not precisely initiate transcription at enterobacterial pro-moters nor could the E. coli enzyme act precisely on
promo-ters of cyanobacterial genes (Schneider et al. 1987; Schyns
et al. 1998). Therefore, the functional activity of the
cyano-bacterial hox promoter in E. coli is interesting. This result shows that the cyanobacterial hox promoter can operate con-stitutively and strongly in E. coli transcription system. How-ever, the transcription levels of hycE, hypA, and hypF, which encode the large subunit of the innate hydrogenase 3 or the accessory proteins for activation of the three hydrogenase isoenzymes in E. coli (Lutz et al. 1991; Jacobi et al. 1992; Sauter et al. 1992), were not altered significantly by intro-ducing the hox genes (Fig. 2A). Similar results have been published previously (Maeda et al. 2007).
To observe the expression of the cyanobacterial proteins HoxEFUYH in the pCC1FOS::pT7::hox and pCC1FOS:: phox::hox cells, SDS-PAGE analysis of the whole cell ex-tracts was performed. The bidirectional hydrogenase is com-posed of five subunits, HoxE, HoxF, HoxU, HoxY and HoxH, with apparent molecular weights of about 20, 61, 28, 24 and 49 kDa, respectively (Fig. 1A). The molecular weight of the native protein (375 kDa) indicates a dimeric assembly Fig. 1. Schematic representation of the cyanobacterial bidirectional hydrogenase and its vector constructs. (A) Schematic diagram of the native
bidirectional hydrogenase, Hox (EFUYH)2(Levin et al. 2004; Tamagnini et al. 2007), (B) Organization of the locus containing the genes
encoding the bidirectional hydrogenase in Synechocystis PCC 6803 (Oliveira et al. 2005). (C) Vectors pCC1FOS::pT7, pCC1FOS:: pT7::hox, and pCC1FOS::phox::hox. The abbreviations, which are designated pT7 or phox, represent the T7 or the putative hox promo-ter, respectively. The putative hox promoter region, which includes binding sites for the LexA transcription activator, is up to 540 bp upstream from the hoxE.
of the enzyme complex, Hox (EFUYH)2(Schmitz et al.
2002). The four subunits, HoxF, HoxH, HoxU, and HoxY, were clearly expressed in both the pCC1FOS::pT7::hox and pCC1FOS::phox::hox cells (Fig. 2B). In contrast, the expres-sion of the smallest subunit HoxE was not observable. This subunit may not have been dissociated from the enzyme com-plex or the larger subunits. However, a recent report indicat-ed that only the HoxU subunit was clearly detectable in a similar E. coli expression system of the hoxEFUYH genes (Maeda et al. 2007). This difference appears to depend on the expression vector system or the cell culture condition applied.
2. Inhibition of hydrogen uptake and enhancement of hydrogenase activity
Increased hydrogen production by E. coli in the absence of uptake hydrogenase activity has been reported by differ-ent authors (Bisaillon et al. 2006; Penfold et al. 2006; Red-wood et al. 2008). Moreover, the expression of the cyano-bacterial hydrogenase HoxEFUYH in E. coli was suggested to increase hydrogen production by inhibiting hydrogen up-take of the innate hydrogenase 1 and 2 (Maeda et al. 2007). Similarly, in the present study, the hydrogen uptake of the pCC1FOS::phox::hox and pCC1FOS::pT7::hox cells was decreased up to about 30% of the negative control cells (Table 1). In contrast, the hydrogenase activities were in-creased up to 192% and 169% in the pCC1FOS::phox::hox and pCC1FOS::pT7::hox cells, respectively, indicating a considerable contribution from HoxEFUYH (Table 1). How-ever, despite this increase in the in vitro assay of hydroge-nase activities, overexpressed HoxEFUYH proteins may not mediate hydrogen production in vivo in the pCC1FOS::phox ::hox and pCC1FOS::pT7::hox cells. Since some mutation events related to the expression and maturation of the innate hydrogenase 3 could eliminate the benefit of expressing the cyanobacterial hydrogenase in E. coli, the enzymatic activity of HoxEFUYH was reported to depend on the in vivo role of hydrogenase 3 (Maeda et al. 2007).
3. Expression of HoxEFUYH proteins and lexA gene after γγ-irradiation
RecA and LexA proteins in E. coli play important roles in the SOS repair system for the DNA damages induced by
genotoxic agents, e.g., UV and γ rays (Walker 1984).
How-ever, the only one regulator LexA has been proven to acti-Table 1. Hydrogen uptake and hydrogenase activity in the control
and transformed E. coli cells. Hydrogen uptake activities of cell suspensions were measured by the oxidized methyl-viologen-based hydrogen uptake assay (Maeda et al. 2007), while total hydrogenase activities were determined in the dark by adding methylviologen and dithionite as artificial electron donors (Appel et al. 2000). Values are means±
S.E. (n==3 or 4).
Hydrogen uptake Hydrogenase activity Strain
(ΔA604for 5 min)
(pmol H2/ml Cell
suspension/OD600)
pCC1FOS::pT7 0.90±0.13 20.4±0.7
pCC1FOS::phox::hox 0.27±0.07 39.1±0.3
pCC1FOS::pT7::hox 0.26±0.06 34.5±1.2
Fig. 2. Expression of hox-related genes and proteins in the
trans-formed E. coli cells, pCC1FOS::pT7, pCC1FOS::phox::hox, and pCC1FOS::pT7::hox. (A) RT-PCR of hoxE, hoxH,
hypA, hypF, hycE, and lexA transcripts. The 16srRNA was
used as a control gene to normalize for differences in the amounts of total RNA. (B) SDS-PAGE of the whole cell extracts from the E. coli cells.
16srRNA hoxE hoxH hypA hypF hycE lexA
pCC1FOS::pT7 pCC1FOS::phox::hox pCC1FOS::pT7::hox
Protein markerpCCIFOS::pT7pCCIFOS::phox::hoxpCCIFOS::pT7::hox 120 kDa 100 kDa 70 kDa 60 kDa 50 kDa 40 kDa 30 kDa 20 kDa HoxF (61) HoxH (49) HoxU (28) HoxY (24) HoxE (20) (A) (B)
vate the transcription of the hoxEFUYH genes in cyanobac-teria (Gutekunst et al. 2005; Oliveira and Lindblad 2005; Tamagnini et al. 2007). Similarly, the expression of hox genes in cyanobacteria was noticeably increased with that of lexA gene after γ-irradiation, depending on the dose of 2 to 30 Gy (data not shown). In the present study, the lexA gene was normally expressed in the pCC1FOS::phox::hox and pCC1FOS::pT7::hox cells as well as the negative control cells (Fig. 2A). To investigate the possible involvement of transcription factor LexA for the expression of hox genes in
the pCC1FOS::phox::hox cells, we carried out γ-irradiation
at a dose rate of 2 or 30 Gy for 10 min. The 30-Gy γ-irradia-tion caused about two-growth-cycle inhibiγ-irradia-tion in all the trans-formed cells including the negative control, while the 2-Gy didn’t induce significant retardation in the growth cycle (Fig. 3A). Unexpectedly, the gene expression of lexA was substan-tially decreased in the pCC1FOS::phox::hox and pCC1FOS ::pT7::hox cells after the 30-Gy γ-irradiation (Fig. 3C). This result may be partly attributed to the absence of recA gene in the E. coli KRX stain used for overexpression of
Hox-EFUYH proteins. However, the decrease of lexA gene expres-sion in the irradiated pCC1FOS::phox::hox cells couldn’t be correlated with the expression levels of HoxEFUYH pro-teins (Fig. 3B). These results suggest that other transcription regulators as well as LexA might contribute to activation of
hox promoter in E. coli.
CONCLUSION
To characterize the expression of the cyanobacterial hyd-rogenase HoxEFUYH in E. coli for biohydrogen production, the putative hox promoter and the full-length hoxEFUYH genes were isolated and transformed into E. coli. The trans-cription and translation of the hoxEFUYH genes were con-stitutively induced in the E. coli cells with the hox promoter as well as the T7. The expression of HoxEFUYH proteins induced a two-fold increase in the total hydrogenase activity and a three-fold decrease in the hydrogen uptake. However, the radiation-induced decrease in the gene expression of Fig. 3. Change in the growth rates and the expressions of HoxEFUYH and lexA in the transformed E. coli cells after γ-irradiation. In the lower
panel of (A), open and closed symbols represent γ-irradiation at a dose rate of 2 or 30 Gy for 10 min, respectively. Error bars represent means±S.E. (n==3). Invisible bars are smaller than symbols. (B) SDS-PAGE of the whole cell extracts from the E. coli cells. (C)
RT-PCR of lexA transcripts. The 16srRNA was used as a control gene to normalize for differences in the amounts of total RNA.
pCC1FOS::pT7::hox_irradiated pCC1FOS::pT7::hox_control pCC1FOS::phox::hox_irradiated pCC1FOS::phox::hox_control pCC1FOS::pT7_irradiated pCC1FOS::pT7_control Protein marker 120 kDa 100 kDa 70 kDa 60 kDa 50 kDa 40 kDa 30 kDa 20 kDa HoxF (61) HoxH (49) HoxU (28) HoxY (24) HoxE (18) 16srRNA lexA 2.5 2.0 1.5 1.0 0.5 0.0 2.0 1.5 1.0 0.5 0.0 OD 600 OD 600 Non-irradiated pCC1FOS::pT7 pCC1FOS::phox::hox pCC1FOS::pT7::hox Irradiated 0 40 80 120 160 200 240 280 Time (min) (A) (B) (C)
LexA, the only one transcription regulator proven for the
hox genes, didn’t alter significantly the expression of
Hox-EFUYH proteins mediated by the hox promoter. Therefore, it is suggested that subsequent studies be conducted to eluci-date other transcription regulators involving in activation of
hox promoter in E. coli.
ACKNOWLEDGMENT
This study was supported by a grant (Code: M2ABA009) to J.-H. Kim from Korea Science and Engineering Founda-tion (KOSEF) and by the Nuclear R & D Program funded by the Ministry of Education, Science and Technology (MEST), Republic of Korea.
REFERENCES
Appel J, Phunpruch S, Steinmüller K and Schulz R. 2000. The bidirectional hydrogenase of Synechocystis sp. PCC 6803 works as an electron valve during photosynthesis. Arch.
Microbiol. 173:333-338.
Bisaillon A, Turcotte J and Hallenbeck PC. 2006. The effect of nutrient limitation on hydrogen production by batch cultures of Escherichia coli. Int. J. Hydrogen Energ. 31:1504-1508. Blokesch M, Paschos A, Theodoratou E, Bauer A, Hube M,
Hutch S and Böck A. 2002. Biochem. Soc. Trans. 30:674-680.
Gutekunst K, Phunpruch S, Schwarz C, Schuchardt S, Schulz-Friedrich R and Appel J. 2005 LexA regulates the bidirec-tional hydrogenase in the cyanobacterium Synechocystis sp. PCC 6803 as a transcription activator. Mol. Microbiol. 58: 810-823.
Jacobi A, Rossmann R and Böck A. 1992. The hyp operon gene products are required for the maturation of catalytically active hydrogenase isoenzymes in Escherichia coli. Arch.
Microbiol. 158:444-451.
Levin DB, Pitt L and Love M. 2004. Biohydrogen production: prospects and limitations to practical application. Int. J.
Hydrogen Energ. 29:173-185.
Li R and Golden S. 1993. Enhancer activity of light-responsive regulatory elements in the untranslated leader regions of cyanobacterial psbA genes. Proc. Natl. Acad. Sci. USA. 90: 11678-11682.
Lutz S, Jacobi A, Schlensog V, Böhm R, Sawers G and Böck A. 1991. Molecular characterization of an operon (hyp) neces-sary for the activity of the three hydrogenase isoenzymes in
Escherichia coli. Mol. Microbiol. 5:123-135.
Maeda T, Vardar G, Self WT and Wood TK. 2007. Inhibition of hydrogen uptake in Escherichia coli by expressing the hydrogenase from the cyanobacterium Synechocystis sp. PCC 6803. BMC Biotechnol. 7:25 (http://www.biomedcen-tral.com/1472-6750/7/25).
Magalon A, Blokesch M, Zehelein E and Böck A. 2001. Fide-lity of metal insertion into hydrogenases. FEBS Lett. 499: 73-76.
Mazouni K, Domain F, Chauvat F and Cassier-Chauvat C. 2003. Expression and regulation of the crucial plant-like ferredo-xin of cyanobacteria. Mol. Microbiol. 49:1019-1029. Oliveira P and Lindblad P. 2005. LexA, a transcription regulator
binding in the promoter region of the bidirectional hydro-genase in the cyanobacterium Synechocystis sp. PCC 6803.
FEMS Microbiol. Lett. 251:59-66.
Ordal EJ and Halvorson HO. 1939. A comparison of hydrogen production from sugars and formic acid by normal and vari-ant strains of Escherichia coli. J. Bacteriol. 38:199-220. Penfold DW, Sargent F and Macaskie LE. 2006. Inactivation
of the Escherichia coli K-12 twin arginine translocation system promotes increased hydrogen production. FEMS
Microbiol. Lett. 262:135-137.
Porter RD. 1988. DNA transformation. pp. 703-712. In: Meth-ods in Enzymology, Section VI. Vol 167, Cyanobacteria (Packer L and Glazer AN eds.), Academic Press Inc., San Diego, CA.
Redwood MD, Mikheenko IP, Sargent F and Macaskie LE. 2008. Dissecting the roles of Escherichia coli hydrogenases in biohydrogen production. FEMS Microbiol. Lett. 278:48-55. Rippka R, Deruelles J, Waterbury JB, Herdman M and Stanier
RY. 1979. Generic assignment, strain histories and proper-ties of pure cultures of cyanobacteria. J Gen. Microbiol.
111:1 -61.
Sambrook J, Fritsch EF and Maniatis T. 1989. Molecular clon-ing: A laboratory manual. 2nd ed., Cold Spring Harbor La-boratory Press, Cold Spring Harbor, NY.
Sauter M, Böhm R and Böck A. 1992. Mutational analysis of the operon (hyc) determining hydrogenase 3 formation in
Escherichia coli. Mol Microbiol. 6:1523-1532.
Sawers RG. 2005. Formate and its role in hydrogen production in Escherichia coli. Biochem. Soc. Trans. 33:42-46. Schmitz O, Boison G, Salzmann H, Bothe H, Schütz K, Wang
S and Happe T. 2002. HoxE - a subunit specific for the pen-tameric bidirectional hydrogenase complex (HoxEFUYH) of cyanobacteria. Biochim. Biophys. Acta 1554:66-74. Schneider GJ, Tumer NE, Richaud C, Borbely G and Haselkorn
R. 1987. Purification and characterization of RNA polyme-rase from the cyanobacterium Anabaena 7120. J. Biol.
Promoter recognition by a cyanobacterial RNA polymerase:
in vitro studies with Calothrix sp. PCC 7601 transcriptional
factors RcaA and RcaD. Plant Mol. Biol. 36:649-658. Tamagnini P, Leitão E, Oliveira P, Ferreira D, Pinto F, Harris
DJ, Heidorn T and Lindblad P. 2007. Cyanobacterial hydro-genases: diversity, regulation and applications. FEMS
Miro-Walker GC. 1984. Mutagenesis and inducible responses to deo-xyribonucleic acid damage in Escherichia coli. Microbiol
Rev. 48:60-93.
Manuscript Received: June 3, 2009 Revision Accepted: June 11, 2009