저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
농학석사학위논문
Potyvirus 에 대한 신규 천연물 유래
물질의 항 바이러스성 특성 구명
Characterizing antiviral activity of the novel
natural compound against potyviruses
2018 년 8 월
서울대학교 대학원
농생명공학부 식물미생물학 전공
배미아
A THESIS FOR DEGREE OF MASTER SCIENCE
Characterizing antiviral activity of the novel
natural compound against potyviruses
BY
MIAH BAE
Department of Agricultural Biotechnology
The Graduate School of Seoul National University
i
ABSTRACT
Characterizing antiviral activity of the novel natural
compound against potyviruses
Miah Bae
Major in Plant Microbiology
Department of Agricultural Biotechnology
The Graduate School of Seoul National University
Plant diseases caused by virus infections reduce the quantity and quality of the
crop product and thus cause significant financial losses all over the world. Unlike
the control strategies for plant pathogenic fungi and bacteria, that of viral diseases
mainly depends on prevention such as quarantine, routine inspection and removal
of diseased plant(s), and development of virus resistant plant cultivar since no
commercial viricide is available yet. The genus Potyvirus is one of the most
ii
important plant RNA virus group and infects wide range of plant species including
several major agricultural plants such as potatoes, tomatoes, soybean, and peppers.
In this study, I tried to identify antiviral activity of the novel compound from Farm
Hannong. For testing the antiviral activity, the compound was sprayed on the
Nicotiana benthamiana plants twice prior to virus challenge inoculations and
observed viral replication as well as symptom developments on systemic leaves.
Inhibition of virus replication was only observed against potyviruses such as potato
virus Y and pepper mottle virus among tested viruses including tomato spotted wilt
virus, pepper mild mottle virus, cucumber mosaic virus, pepper mottle virus, potato
virus X, and potato virus Y. Furthermore, similar level of inhibitory activity was
also observed against another potyviruses such as soybean mosaic virus and chilli
veinal mottle virus. No virus replication was observed on the upper systemic leaves.
To identify genes and/or signal transduction pathway(s) responsible for this
phenomenon, I investigated induction or suppression of major genes related to
several signal transduction pathways upon treatment of the novel compound by
conducting qRT-PCR using specific primer sets for each gene. Interestingly, gene
expression levels of EIN2 and PDF1.2 involved in JA/ET hormone pathway were
increased in the treated plants compared to those of mock plants at four days after
compound treatment. Taken together, my results suggest that this natural compound
might be the candidate agent specifically inhibiting potyvirus infections and/or
replications.
iii
Keywords: potyvirus, natural compound, antiviral activity, inhibition of virus
replication, induced resistance, plant defense pathways
iv
CONTENTS
Page
ABSTRACT………i
CONTENTS………iv
LIST OF TABLES………vi
LIST OF FIGURES………vii
I. INTRODUCTION………1
II. MATERIALS AND METHODS………3
1. Plant material and virus inoculation………3
2. Foliage spray………3
3. GFP imaging………3
4. Reverse transcription-polymerase chain reaction (RT-PCR)…………4
v
III. RESULTS………7
1. Treatment of the natural compound does not show side effect(s) on
N. benthamian………7
2. The natural compound shows inhibitory activity against potyvirus
infections………7
3. Treatment using 1% of the compound was effective in protecting N.
benthamiana from potyvirus infections ………12
4. The antiviral activity of the natural compound against potyviruses on
different host plants………13
5. Effect of the compound treatment on replication of PVX…………16
6. Effective dose of the natural compound……….18
7. Effect of the compound on curing virus infections………18
8. Effects of treatment with the natural compound on expression of
several genes related to major signal transduction pathway……21
IV. DISCUSSION………24
V. LITERATURE CITED .………28
VI. ABSTRACT IN KOREAN ………35
vi
LIST OF TABLES
Page
Table 1. List of primers used for qRT-PCR to investigate genes expression
profiles of the selected genes in Nicotiana benthamiana………5
vii
LIST OF FIGURES
Page
Fig. 1. Testing possible side effect(s) of the natural compound on Nicotiana
benthamiana………9
Fig. 2. Effect of treating the natural compound against six viruses on N.
benthamiana ………10
Fig. 3. Inhibitory effect of the compound at different concentrations, i.e. 1%
and 0.2% (v/v), against PVY, PepMoV and ChiVMV on N.
benthamiana………14
Fig. 4. Inhibitory effect of the compound at different concentrations, i.e. 1%
and 0.2% (v/v), against SMV and PepMoV on soybean and pepper
plants, respectively………15
Fig. 5. Replication of PVY viral RNA ………17
Fig. 6. Minimum inhibitory concentration of the compound against PepMoV
………19
Fig. 7. Curative effect of the compound against PVY………20
Fig. 8. The expression pattern of defense related genes ………22
1
INTRODUCTION
Viruses, pathogens that are smaller than bacteria, can infect most, if not all, living organisms such as human, animal, plant, insect, and fungus. Among them, plant viruses can decrease the yield and reduce commercial value result in economic losses (Bos, 1982). For example, potato virus Y infection resulted in annual potato losses of up to 2.8 tons per hectare in Kenya (Nyamwamu et al., 2014). In addition, tomato spotted wilt virus caused losses around US$1 billion every year all over the world (Karavina et al., 2017). According Animal and Plant Quarantine Agency in Korea (2018), plum pox virus is controlled as the import prohibition disease agent since it will be able to cause great damage if it is introduced into the country. Several control strategies such as removal of plants with symptoms, insect control, cross protection using mild strain virus, and virus resistance cultivar are used in agriculture because there is no commercially available viricide (Nishiguchi et al., 2011; Hooks et al., 2006). Thiouracil, ribavirin and acibenzolar-S-methyl have been reported to be effective in inhibiting replication of plant viruses such as potato virus X, potato virus Y, tobacco mosaic virus. However, they were not registered as viricides due to side effect, environmental pollution of dosages for inhibition activity and short lasted time of inhibitory effect against virus in field (Hansen. 1989; Bawden and Kassanis. 1954; Lerch B. 1987; Oostendorp et al,. 2001). Therefore, development or identification of novel antiviral compounds that can be used as viricide is needed.
2
RNA genome of approximately 10 kb and is regarded as one of the most important plant RNA virus (Wylie et al., 2017). They comprise approximately 30% of currently known plant viruses and are transmitted by aphid in non-persistent manner (Ivanov et al., 2014; Whitfield et al., 2015). Potyviruses have broad host ranges infecting both monocotyledonous and dicotyledonous plants. Members of potyviruses include potato virus Y (PVY), pepper mottle virus (PepMoV), soybean mosaic virus (SMV), and sugarcane mosaic virus (Urcuqui-Inchima et al., 2001).
In this study, I tested antiviral activity of the novel compound from Farm Hannong against various viruses such as PVY, potato virus X (PVX), cucumber mosaic virus (CMV), pepper mild mottle virus (PMMoV), tomato spotted wilt virus (TSWV), and PepMoV. Inhibition of potyvirus replications was observed on Nicotiana benthamiana plants treated with the novel natural compound. To identify genes and/or signal transduction pathway(s) responsible for this phenomenon, I also investigated induction or suppression of major genes related to several signal transduction pathways upon treatment of the novel compound by conducting qRT-PCR using specific primer sets for each gene.
3
MATERIAL AND METHODS
1. Plant material and virus inoculation
All used plants such as N. benthamiana, pepper, soybean were grown at 25℃ in growth chamber with 16h light and 8h dark cycles. Virus was inoculated with sap using 0.05 M potassium phosphate buffer (pH 7.4). The powder formulation of natural compound from Farm Hannong was diluted in water as 0.2% to 1% concentration and add 100 ppm of Tween 80.
2. Foliage spray
Each diluted compound was sprayed twice, i.e. first spray onto the three-week-old
N. benthamiana and the second spray 4 days later, and then challenge inoculated with tested
viruses by sap inoculation at 3 days post-2nd spray.
3. GFP imaging
Green fluorescent protein was observed under the Dark readerTM Hand Lamp
HL28T UV Lamp (Clare Chemical Research, Inc., USA) using Nikon D80 digital SLR camera with UV filter.
4
4. Reverse transcription-polymerase chain reaction (RT-PCR)
Total RNAs were extracted using RNAiso Plus (TaKaRa, Japan) from the upper systemic leaves of the plants. cDNA was synthesized with oligo dT primers. RT-PCR was carried out using virus specific primer sets. Total of 20 µl PCR reaction mixture was used for PCR reaction containing 0.1 µl of Ex-Taq, 2 µl of 10 X PCR buffer, 1.6 µl of dNTP mixture (TaKaRa, Japan), 1 µl of cDNA (50ng/ µl), 1 µl each of forward and reverse primers, and 13.3 µl of H2O.
5. Quantitative real-time RT-PCR (qRT-PCR)
After the second spraying, the leaves were collected at 1 hour, 2 and 4 days post-treatment. Total RNA extraction was conducted using RNeasy Plant Mini kit (Qiagen) from the collected leaves. For the qRT-PCR, cDNA was prepared using random hexamer primers. Total of 10 µl of reaction mixture containing 5 µl of 2XSYBR Green supermix (Bio-Rad), 1 µl each of forward and reverse primers, 1 µl of cDNA (25 ng/µl), and 2 µl of H2O was
5
Table 1. List of primers used for qRT-PCR to investigate genes expression profiles of the
selected genes in Nicotiana benthamiana.
Primer Sequence (5'->3') NbHSP90 Fw ATGGCGGAGGCAGAGACGTT NbHSP90 140 Rv CGGATCTTGTCCAGAGCATCGGAA NbSGT1 111 Fw GCTTTTCGCCGACCGTGCTC NbSGT1 240 Rv ACAGGCCAACCCCTTACGCA NbEDS1 271 Fw TGTTGGCACAGATGAGGTAGCCA NbEDS1 414 Rv CCCGACGAGTGCCCTGCAAA NbNDR1 102 Fw CAATAATACATGCTGCTTTAAGTGCATCC NbNDR1 208 Rv CTAAAGAGCAAGATGGTTTTGTGGTTCT NtNPR1 273 Fw TGCGGCTTCTCCGGAGTTTG NtNPR1 378 Rv CGGACTCCTCGCCGACAAAA NbEIN2 6 Fw GCCGGAGCTATGGGGGAAAT NbEIN2 148 Rv GGGGGCTTGATTTTTGCTGCC NbCOI1 385 Fw TGGCGAAGGGATGCCTTGAA NbCOI1 511 Rv CGGTCAAGCAAAACCAGCCG NbRbohB 713 Fw TCTGGGATCAAGTGGCTGACCA NbRbohB 846 Rv GTTGGCAGACGCGCTAAGGC NbNOA1 1116 Fw TCCTCAAAGTCGTCTCCGTGCT NbNOA1 1245 Rv AATTCGCACGAGGCCTCCCC NbPDF1.2-Fw CTTCAAGCAAAGCTGCAGCCAAAG NbPDF1.2-Rv CTATGCACTAAGCCATGTGTGTTTG NbPR5 Fw ATGCGCAGCCCCTATTAAC NbPR5 Rv TGGGTTGTTACATCCACCTTG NbERF1 Fw GTTAACGCCGTCAAGTTGGT NbERF1 Rv AGAGGCGGCACCTCAAATA
6
NbICS1 12 Fw GAACCATGTCCTCCTCCCGC NbICS1 98 Rv GGGTGAGAAGTTCCGTTGAAGCA NbPal1 1305 Fw TGGAAAGTGGCAACCCTGCT NbPal1 1427 Rv CACTCCTCGCCCGGTTGACCG Nb actin Fw CCAGGTATTGCTGATAGAATGAG Nb actin Rv CTGAGGGAAGCCAAGATAGAG Nb Ubi3 Fw GCCGACTACAACATCCAGAAGG Nb Ubi3 Rv TGCAACACAGCGAGCTTAACC7
RESULTS
1. Treatment of the natural compound does not show side effect(s) on N. benthamiana
To investigate any possible side effect(s) caused by foliage spraying of the natural compound on the plants, the compound was treated on the N. benthamiana without virus infection. After the second spraying, any significant changes between the treatment and control plants were observed for at least 6 weeks. No notable difference on plants between control and treated plants was observed (Fig. 1) indicating that the treatment of the natural compound has no side effect.
2. The natural compound shows inhibitory activity against potyvirus infections
To identify activity of this compound, I conducted the first screening against the six viruses belonging to 5 different genus. PVX of the genus Potexvirus (Jeevalatha et al., 2016), CMV of the genus Cucumovirus (Moyle et al., 2018), TSWV of the genus
Tospovirus (Moyle et al., 2016), PVY and PepMoV of the genus Potyvirus (Zhang et al.,
2015), and PMMoV of the genus Tobamovirus (Secrist and Ali, 2018) were used in this experiment. For PVX, CMV and PepMoV inoculation, GFP fused infectious DNA clones were used as inocula, while each virus-infected leaves was used for PVY, TSWV, and PMMoV inoculations into N. benthamiana.
8
For initial screening, each tested virus was inoculated onto the plants that were pre-treated with 1% concentration of the compound. In case of PVX::GFP and CMV::GFP, the expression of GFP was observed on the upper systemic leaves of both treated and untreated plants (Fig. 2A and B). For the screening of PMMoV and TSWV, the symptoms of two viurses were observed on both of treated and untreated plants (Fig. 2D and E). However, the treated plants infected by PVY did not have viral symptoms, and the clear viral symptom of PVY was observed on the upper leaves of untreated plants (Fig. 2F). Untreated plant showed GFP expression on the systemic leaves after PepMoV::GFP inoculation, by contrast, none of GFP expression was observed in the upper leaves of treated plants inoculated by PepMoV::GFP (Fig. 2C). These data suggest that this natural compound has inhibitory activity against potyvirus infections including PVY and PepMoV.
9
Fig. 1. Testing possible side effect(s) of the natural compound on Nicotiana benthamiana. H, untreated healthy; 1%, treatment with 1% of the compound; 0.2%, treatment with 0.2% of the compound.
11
Fig. 2. Effect of treating the natural compound against six viruses on N. benthamiana. After treatement, viruses such as PVX::GFP (A); CMV::GFP (B), PVY (C); TSWV (D); PVY (E); PepMoV::GFP (F) were inoculated. The plants which were inoculated using GFP fused infectious clone were observed under UV (A-C). The plants inoculated using infected tissue were observed under light (E-F). C and H indicate untreated control with indicated virus inoculation and un-inoculated healthy plants, respectively.
12
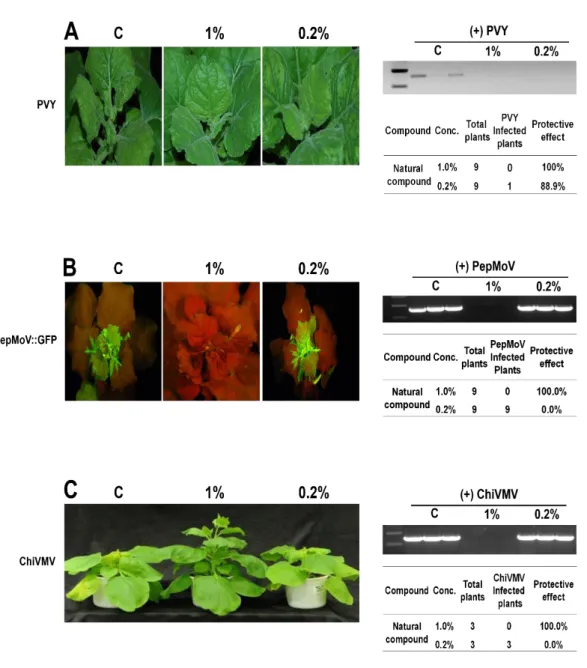
3. Treatment using 1% of the compound was effective in protecting N. benthamiana from potyvirus infections.
In previous result, treatment with 1% (v/v) of the compound was effective in inhibiting potyvirus infections. To further estimate the efficacy of this compound at lower concentrations, the compound was treated at a concentration of 0.2% (v/v) on N.
benthamiana. In this experiment, for screening the viral symptoms were observed visually
on the upper leaves and virus was detected by RT–PCR using virus specific primer. As a result, treatment with 1% showed 100% protective effect and treatment with 0.2% has 88.9% protective effect against PVY (Fig. 3A). Accordingly, treatment with 0.2 to 1% of compound was effective to prevent PVY infection. In the case of PepMoV, 100% protective effect was observed for the 1% compound treatment, but was not effective in preventing virus infections with 0.2% treatment (Fig. 3B). Therefore, PepMoV viral replication and systemic movement were inhibit only on plants treated with 1% (v/v) compound treatment. To test inhibitory activity of this compound against other species of potyvirus, efficacy test was performed against chilli vainal mottle virus (ChiVMV) (Zhang et al., 2015). In case of ChiVMV, the plants treated with 0.2% of compound appeared to have the viral symptoms and virus was detected by RT-PCR (Fig. 3C). However, treatment with the compound that was diluted into 1% concentration was effectively prevents ChiVMV replication and symptoms development on upper systemic leaves. These results indicated that treatment with 1% (v/v) of this compound has effectivity against another species of potyvirus.
13
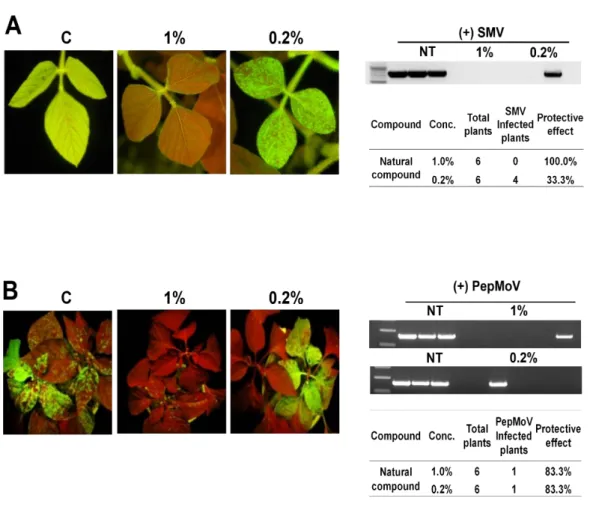
4. The antiviral activity of the natural compound against potyviruses on different host plants
To investigate the effect of this compound in inhibiting potyvirus replication and/or infection on different host plants, I carried out similar experiments on soybean (Glycine
max (L.) Merr.; Ahangaran et al., 2013) and pepper (Capsicum spp.; Kenyon et al., 2014).
Soybean mosaic virus (SMV) and PepMoV expressing GFP from full-length infectious DNA clones were inoculated on soybean and pepper, respectively. SMV::GFP viral replication and systemic movement were inhibited on the plants treated with 1% of compound (Fig. 4A). The expression of GFP was observed on the systemic leaves in four soybean plants among six plants that treated with 0.2% of compound. Treatment with 0.2% of compound was decreased protective effect of the compound comparing to the treatment with 1% of compound. However, treatment using both 0.2% and 1% of the compound inhibited PepMoV::GFP replication in upper systemic leaves of pepper plants in 5 out of 6 tested pepper plants (Fig. 4B). These results demonstrated that natural compound was effective not only on N. benthamiana but also on soybean and pepper plants as well.
14
Fig. 3. Inhibitory effect of the compound at different concentrations, i.e. 1% and 0.2% (v/v), against PVY (A), PepMoV (B) and ChiVMV (C) on N. benthamiana. C represents untreated control with virus infections.
15
Fig. 4. Inhibitory effect of the compound at different concentrations, i.e. 1% and 0.2% (v/v), against SMV (A) and PepMoV (B) on soybean and pepper plants, respectively. C represents untreated control with virus infections.
16
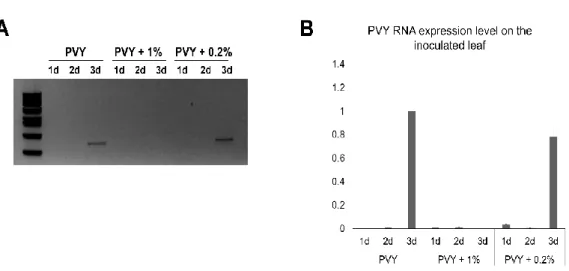
5. Effect of the compound treatment on replication of PVY
To confirm inhibitory effect of the compound on the upper systemic leaves was caused by reduced or no replication of the PVY on the inoculated leaves, I conducted RT-PCR and qRT-RT-PCR analyses to determined viral RNA replication using total RNAs extracted from inoculated leaves. Leaf disc samples were collected from inoculated leaves at 1, 2, and 3 days post inoculation (dpi), respectively, and the extracted total RNAs were then used for RT-PCR and qRT-PCR analyses using PVY coat protein (CP) specific primers. Interestingly, I could not detect any amplified DNA band by conducting RT-PCR with PVY CP specific primers using total RNAs extracted from inoculated leaves that were treated with 1% (v/v) of compound and inoculated with PVY (Fig. 5A). Result of qRT-PCR also showed that the inoculated leaves using 1% of compound was effectively inhibited the accumulation of PVY viral RNA (Fig. 5B). In contrast, DNA band of expected size was detected from samples treated with water and with with 0.2% of the compound by RT-PCR after 3 dpi. qRT-PCR result also showed similar result. The accumulation levels of PVY viral RNA in inoculated leaves on water treated (control) and on 0.2% (v/v) treatment of the compound were significantly higher than that treated with 1% of compound at 3dpi. These results indicated that treatment using 1% (v/v) of the compound inhibited replication of PVY viral RNA in the inoculated leaves and thus possibly able to restrain viral movement to the systemic leaves.
17
Fig. 5. Replication of PVY viral RNA. Total RNAs were extrated from water (control), 1% (v/v), and 02%(v/v) of the compoundt treated PVY inoculated local leaves at 1, 2, and 3 dpi and subjected to RT-PCR (A) and qRT-PCR (B).
18
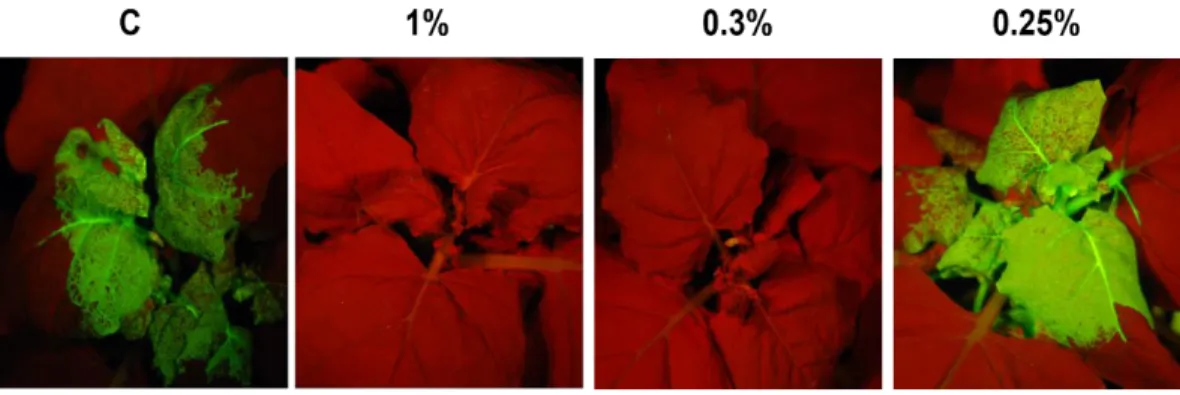
6. Effective dose of the natural compound
To find out minimum inhibitory concentration of this compound, sap from pPepMoV::GPF infected plant, which had a 100% infection rate in the plants treated with 0.2% of the compound, was inoculated on the plants that were pre-treated with different concentration of the compound. As shown in Fig. 6, expression of GFP was observed on the systemic leaves of the plants treated with 0.25% (v/v) of the compound. However, the expression of GFP was not appeared on the plants treated with 1% (v/v) and 0.3% (v/v) of the compound. These results suggest that this compound is effective in inhibiting PepMoV replication and systemic movement when used with 0.3% (v/v) or higher concentration.
7. Effect of the compound on curing virus infections
As mentioned above, I confirmed the protective effect of this compound against potyviruses (Fig. 3 and 4). To determine whether this conpound has curing effect against potyviruses, 1% of the compound was sprayed two times on the plants that were infected with PVY with typical viral. Typical PVY viral symptoms was observed and maintained on the upper systemic leaves of both water treated control and 1% of the compound treatment (Fig. 7). There was no significant difference on delaying symptom appearance or lessening symptom severity between control and the treatment plants. Taken together, these results demonstrated that this compound has no curing effect against PVY.
19
Fig. 6. Minimum inhibitory concentration of the compound against PepMoV. C represents water treated control while 1%, 0.3%, and 0.25% represent the compound trreated at indicated dilution (v/v). PepMoV expressing GFP (PepMoV::GFP) was challenge inoculated 2 days post-treatment and observed for expression of GFP under UV light.
20
Fig. 7. Curative effect of the compound against PVY. H represents un-inoculated healthy plant. C and 1% represent water treated control and 1% (v/v) of the compound treated plants, respectively. Each plant was infected with PVY prior to treatment. Diluted compound or water was applied when plants showed typical PVY symptoms.
21
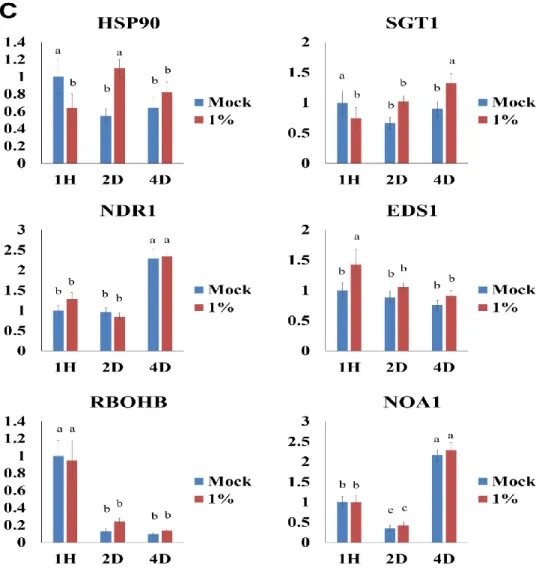
8. Effects of treatment with the natural compound on expression of several genes related to major signal transduction pathways
To get more insights on how this compound cause inhibitory effect on potyvirus replications, qRT-PCR was performed using specific primer sets detecting several genes related to major signal transduction pathways. ICS1, PAL1, NPR1, and PR5 genes were selected as candidate genes for the SA hormone pathway while COI1, EIN2, ERF1, and PDF1.2 genes were used as candidates for JA/ET hormone pathway. HSP90, SGT1, EDS1, and NDR1 genes that are R gene related pathway were also selected. In addition, RBOHB and NOA1 genes related to HR and NO pathway, respectively, were selected. To observe expression profiles of these selected genes, I collected leaf samples at an hour, 2 days, and 4 days after treatment of the compound at 1% (v/v) concentration without virus challenge inoculation, extracted total RNAs, and applied the for qRT-PCR analysis. The expression of HSP 90, SGT1, EDS1, NDR1, RBOHB, and NOA1 did not show significant difference between control and treated plants at all time points tested (Fig. 8C). The SA related genes had similar gene expression level between control and treated plants (Fig. 8A). However, the expression of EIN2 and PDF1.2, JA/ET hormone pathway related genes, were increased in treated plants at 4 days after treatment compared to those in control (Fig. 8B). These results suggest that this compound might regulate JA/ET hormone pathway related to defense responses and thus show inhibitory effect against potyvirus infections.
23
Fig. 8. The expression pattern of defense related genes. ICS1, PAL1, NPR1 and PR5 genes are related to the SA hormone pathway (A). COI1, EIN2, ERF1 and PDF1.2 genes are involved in JA/ET hormone pathway (B). HSP90, SGT, NDR1 and EDS1 genes are related to R gene and additionally RBOHB and NOA1 genes are required to HR and NO pathway respectively (C).
24
DISCUSSION
New plant viruses have been emerging continuously since virus replication time is short and the evolution of virus is fast. Moreover, damages caused by plant viruses has been increased in agriculture (Hanssen et al,. 2010; Islam et al,. 2017; Varma and Malathi. 2003), but no applicable viricide that can be used to cure these damages (Islam, 2017). In this study, I tried to find the novel antiviral compound which can be used in treating viral diseases. So far, most previously identified compounds showed side effects on the plants (Hansen. 1989; Bawden and Kassanis. 1954; Lerch B. 1987; Oostendorp et al,. 2001). Based upon my experiment results, however, treatment of the natural compound onto plants did not show any significant side effects on N. benthamiana, pepper, and soybean plants (Fig. 1). At the first screening, viral symptoms of both PVY and PepMoV, from genus
Potyvirus, were not observed on the upper systemic leaves treated with the compound
among six tested viruses such as CMV, PepMoV, PMMoV, PVX, PVY, and TSWV (Fig. 2). In addition, replication of SMV which is another species of potyvirus, was also inhibited on the plants treated with 1% of compound (Fig. 4A). Phylogenetic analysis of PVY and SMV revealed that these two viruses are distantly related although both viruses are classified into the same genus (Gibbs and Ohshima. 2010). Treatment of the compound was effective on N. benthamiana used as an experimental plant (Goodin et al., 2008) as well as on both soybean and pepper plants (Figs. 3 and 4). These results suggest that treatment of this compound could be effective against broad species of potyvirus on various host plants. Most members of potyviruses form similar particles and thus might have similar epitope(s)
25
outside virions (Wylie. 2017). Plant viral coat protein (CP) plays crucial role in cell to cell and long distance movement, protection viral RNA, assembly and determinant for vector transmission during virus infection (Besong-Ndika et al,. 2015; Dolia et al,. 1995; Seo et al,. 2016; Tatineni et al,. 2018). Therefore, it is possible that viral protein(s) like CP, which is related with the resistance activity, somehow were activated upon the compound treatment and further were recognized as epitope(s), resulting in an activation of defense responses (Kang et al,. 2005; Van Der Biezen and Jones. 1998). As the result of these defence response activation, potyviruses failed to replicate and thus viral CP was not detected in the inoculated leaves (Fig. 5). The minimum inhibition concentration of the compound was up to treatment with 0.3% concentration that is lower than treatment with 1% (Fig. 6). Price is also an important consideration for the commercialization of pesticide (Whitford et al,. 2006) and if they are effective at lower concentrations, they are considered to be more competitive as candidates for the development of viricide.
The hormone pathway of both SA and JA/ET is well known as a plant defense response to biotic and abiotic stresses (Pieterse et al., 2009; Vallad and Goodman. 2004; Zarate et al., 2007). This response is able to induce resistance in whole plant and to protect viral infection (Waziri. 2015). Treatment of SA or JA can induce systemic acquired resistance as well (Dong. 1998; Gaffney et al,. 1993; Gao et al,. 2013). I conducted qRT-PCR to investigate expression level of genes which are related to defense hormone pathways. As a result, at 4 dpi, expressions of EIN2 and PDF1.2 genes involved in JA/ET hormone pathway in treated plants were increased compared to those of water treated
26
control (Fig. 8). These results suggest that the JA / ET hormone pathway was induced after treatment of the compound, result in resistance mechanism. The JA / ET pathway has been reported to proceed in the order of EIN2, ERF1, and PDF1.2 (Pieterse et al., 2009; Zarate et al., 2007). However, in this experiment, no significant difference in the expression level of ERF1 gene between untreated and treated plants was observed. Additional experiments with NahG (SA pathway) and jar1 (JA/ET pathway) mutants that are not able to induce systemic resistance through defense hormone pathway are necessary to confirm that the JA/ET pathway is responsible for the resistance response that induced by natural compound (Jia et al,. 2016; Pieterse and Van Loon. 1999). It is also very interesting and thus worth pursuing additional experiments to understand how the induction of several genes related with the JA/ET pathway effectively inhibit only potyvirus replications.
In summary, I found the natural compound was effective against potyvirus. Treatment with 1% of compound before virus inoculation was very effective against PVY, PepMoV, ChiVMV, and SMV infections on N. benthamiana, soybean, and pepper plants. However, post treatment using 1% of compound was not effective in curing potyvirus infections. Result of qRT-PCR showed that the expression level of some genes related to JA/ET pathway was increased. I assumed that inhibitory activity of this compound might be regulated by genes involved in JA/ET hormone pathway. In the future, it will be possible to use this natural compound as a plant virus control agent assuming further study on understanding the resistance mechanism upon the natural compound treatment is conducted by using NahG (SA pathway) and jar1 (JA/ET pathway) mutants and by comparing
27
28
LITERATURE CITED
Ahangaran, A., Habibi, M. K., Mohammadi, G. H. M., Winter, S., and Garcia-Arenal, F. 2013. Analysis of Soybean mosaic virus genetic diversity in Iran allows the characterization of a new mutation resultion in overcoming Rsv4-resistance. J. Gen. Virol. 94: 2557-2568.
Arasimowicz, M., and Floryszak-Wieczorek, J. 2007. Nitric oxide as a bioactive signalling molecule in plant stress responses. Plant Sci. 172: 876-887.
Bawden, F. C., and Kassanis, B. 1954. Some effects of thiouracil on virus-infected plants. J. Gen. Microbiol. 10: 160-173.
Besong-Ndika, J., Ivanove, K. I., Hafren, A., Michon, T., and Makinen, K. 2015. Co-translational CP-mediated inhibition of potyviral RNA translation. J Virol. 89: 4237-4248.
Bos, L. 1982. Crop losses caused by viruses. Crop Protection 1: 263-282.
Chen, L. J., Zou, W. S., Wu, G., Lin, H. H., and Xi, D. H. 2018. Tobacco alpha-expansin EXPA4 plays a role in Nicotiana benthamiana defence against tobacco mosaic virus. Planta 247: 355-368.
Deng, X. G., Zhu, T., Zou, L. J., Han, X. Y., Zhou, X., Xi, D. H., Zhang, D. W., and Lin, H. H. 2016. Orchestration of hydrogen peroxide and nitric oxide in brassinosteroid-mediated systemic virus resistance in Nicotiana benthamianaI. Plant J. 85: 478-493.
29
J. C. 1995. Protein determinants involved in cell-to-cell and long-distance movement of tobacco etch potyvirus. Virology 206: 1007-1016.
Dong, X. 1998. SA, JA, ethylene, and disease resistance in plants. Curr. Opin. Plant Biol. 1: 316-323.
Gibbs, A., and Ohshima, K. 2010. Potyviruses and the digital revolution. Annu. Rev. Phytopathol. 48: 205-223.
Gaffney, T., Friedrich, L., Vernooij, B., Negrotto, D., Nye, G., Uknes, S., Ward, E., Kessmann, H., and Ryals, J. 1993. Requirement of salicylic acid for the induction of systemic acquired resistance. Science 261: 754-756.
Gao, W., Long, L., Zhu, L., Xu, L., Gao, W., Sun, L., Liu, L., and Zhang, X. 2013. Proteomic and virus-induced gene silencing (VIGS) analyses reveal that gossypol, brassinosteroids and jasmonic acid contribute to the resistance of cotton to
Verticillium dahiae. Mol. Cell. Proteomics 12: 3690-3703.
Gibbs, A., and Ohshima, K. 2010. Potyviruses and the digital revolution. Annu. Rev. Phytopathol. 48: 205-223.
Goodin, M. M., Zaitlin, D., Naidu, R. A., and Lommel, S. A. 2008. Nicotiana benthamiana: Its history and future as a model for plant-pathogen interaction. Mol. Plant-Microbe Interact. 21: 1015-1026.
Hansen, A. J. 1989. Antiviral chemicals for plant-disease control. Crit. Rev. Plant Sci. 8: 45-88.
Hanssen, I. M., Lapidot, M., and Thomma, B. P. H. J. 2010. Emerging viral diseases of tomato crops. Mol. Plant-Microbe Interact. 23: 539-548.
30
Hooks, C. R. R., and Fereres, A. 2006. Protecting crops from non-persistently aphid-transmitted viruses: A review on the use of barrier plants as a management tool. Virus Res. 120: 1-16.
Islam, W. 2017. Management of plant virus diseases; farmer’s knowledge and our suggestions. Hosts Viruses 4: 28-33.
Islam, W., Zhang, J., Adnan, M., Noman, A., Zaynab, M., and Wu, Z. 2017. Plant virus ecology: A glimpse of recent accomplishments. Appl. Ecol. Env. Res. 15: 691-705. Ivanov, K. I., Eskelin, K., Lohmus, A., and Makinen, K. 2014. Molecular and cellular
mechanisms underlying potyvirus infection. J. Gen. Virol. 95: 1415-1429.
Jeevalatha, A., Kaundal, P., Kumar, P., Raigond, B., Gupta, M., Kumar, A., Sharma, S., Sagar, V., Nagesh, M., and Singh, B. P. 2016. Analysis of the coat protein gene of Indian potato virus X isolates for identification of strain groups and determination of the complete genome sequence of two isolates. Eur. J. Plant Pathol. 145: 447-458.
Jia, X., Meng, Q., Zeng, H., Wang, W., and Yin, H. 2016. Chitosan oligosaccharide induces resistance to tobacco mosaic virus in Arabidopsis via the salicylic acid-mediated signalling pathway. Sci. Rep. 6:26144.
Jones, R. A. C. 2006. Control of plant virus diseases. Adv. Virus Res. 67: 205-244.
Kang, B., Yeam, I., and Jahn, M. 2005. Genetices of plant virus resistance. Annu. Rev. Phytopathol. 43: 581-621
Karavina, C., and Gubba, A. 2017. Detection and characterization of tomato spotted wilt virus infecting field and greenhouse-grown crops in Zimbabwe. Eur J Plant Pathol.
31
149: 933-944.
Kenyon, L., Kumar, S., Tsai, W. S., and Hughes, J. D. 2014. Virus sisease of peppers (Capsicum spp.) and their control. Adv. Virus Re. 90: 297-354.
Lerch, B. 1987. On the inhibition of plant-virus multication by ribavirin. Antivir. Res. 7: 257-270.
Liu, Y. L., Burch-Smith, T., Schiff, M., Feng, S. H., and Dinesh-Kumar, S. P. 2004. Molecular chaperone Hsp90 associates with resistance protein N and its signaling proteins SGT1 and Rar1 to modulate an innate immune response in plants. J. Biol. Chem. 279: 2101-2108.
Moran, P. J., and Thompson, G. A. 2001. Molecular responses to aphid feeding in
Arabidopsis in relation to plant defense pathways. Plant Physiol. 125: 1074-1085.
Moyle, R., Pretorius, L. S., Dalton-Morgan, J., Persley, D., and Schenk, P. 2016. Analysis of the first complete genome sequence of an Australian tomato spotted wilt virus isolate. Australas Plant Path. 45: 509-512.
Moyle, R., Pretorius, L. S., Shuey, L. S., Nowak, E., and Schenka, P. M. 2018. Analysis of the complete genome sequence of cucumber mosaic virus strain K. Genome Announc. 6: e00053-18.
Nishiguchi, M., and Kobayashi, K. 2011. Attenuated plant viruses: preventing virus diseases and understanding the molevular mechanism. J. Gen. Plant Pathol. 77: 221-229.
Nyamwamu, PA., Mukoye, B., Osogo, AK., Omuse, CN., Ajanga, S., and Were, HK. 2014. Distribution and biological characterization of potato virus Y in Kenya. J.
Agri-32
Food & Appl. Sci. 2: 258-264.
Oostendorp, M., Kunz, W., Dietrich, B., and Staub, T. 2001. Induced diseases resistanve in plants by chemicals. Eur. J. Plant Pathol. 107: 19-28.
Pieterse, C. M. J., and Van Loon. 1999. Salicylic acid-independent plany defence pathways. Trends Plant Sci. 4: 52-58.
Pieterse, C. M. J., Leon-Reyes, A., Van der Ent, S., and Van Wees, S. C. M. 2009. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 5: 308-316.
Raffaello, T., and Asiegbu, F. O. 2017. Small secreted proteins from the necrotrophic conifer pathogen Heterobasidion annosum s. l. (HaSSPs) induce cell death in
Nicotiana benthamiana. Sci. Rep. 7: 8000.
Ramegowda, V., Senthil-Kumar, M., Ishiga, Y., Kaundal, A., Udayakumar, M., and Mysore, K. S. 2013. Drought stress acclimation imparts tolerance to Sclerotinia
sclerotiorum and Pseudomonas syringae in Nicotiana benthamiana. Int. J. Mol.
Sci. 14: 9497-9513.
Secrist, K., and Ali, A. 2018. First complete genome sequence of pepper mild mottle virus from chili pepper in the United States. Genome Announc. 6: e00331-18.
Seo, J. K., and Kim, K. H. 2016. Long-distance movement of viruses in plants. In: Current Research Topics in Plant Virology. pp. 153-172. Springer.
Tatineni, S., McMechan, A. J., and Hein, G. L. 2018. Wheat streak mosaic virus coat protein is a determinant for vector transmission by the wheat curl mite. Virology 514: 42-49.
33
Tran, P. T., Choi, H., Choi, D., and Kim, K. H. 2016. Virus-induced gene silencing reveals signal transduction components required for the Pvr9-mediated hypersensitive response in Nicotiana benthamiana. Virology. 495: 167-172.
Urcuqui-Inchima, S., Haenni, A. L., and Bernardi, F. 2001. Potyvirus proteins: a wealth of functions. Virus Res. 74: 157-175.
Vallad, G. E., and Goodman, R. M. 2004. Systemic acquired resistance and induced systemic resistance in conventional agriculture. Crop Sci. 44: 1920-1934.
Van Der Biezen, E. A., and Jones, J. D. G. 1998. Plant disease-resistance proteins and the gene-for gene concept. Trends Biochem. Sci. 23: 454-456.
Varma, A., and Malathi, V. G. 2003. Emerging geminivirus problemsL A serious threat to crop production. Ann. Appl. Biol. 142: 145-164.
Waziri, H. M. A. 2015. Plants as antiviral agents. J. Plant Pathol. Microbiol. 6: 1.
Whitfield, A. E., Falk, B. W., and Rotenberg, D. 2015. Insect vector-mediated transmission of plant viruses. Virology 479: 278-289.
Whitford, F., Pike, D., Burroughs, F., Hanger, G., Johnson, B., and Brassard, D. 2006. The pesticide marketplace, discovering and developing new products. http://www.extension.purdue.edu/extmedia/PPP/PPP-71.pdf.
Wylie, S. J., Adams, M., Chalam, C., Kreze, J., Lopez-Moya, J. J., Ohshima, K., Praveen, S., Rabenstein, F., Stenger, D., Wang, A. M., Zerbini, F. M., and ICTV Report Consortium. 2017. ICTV virus taxonomy profile: Potyviridae. J. Gen. Virol. 98: 2893-2893.
34
acid defenses and suppresses effectual jasmonic acid defenses. Plant Physiol. 143: 866-875.
Zhang, S. B., Zhao, Z. B., Zheng, L. M., Liu, J., Peng, J., Yan, F., Li, F., Xie, Y., Cheng, Z. B., Zhou, X. G., Zhang, D. Y., and Liu, Y. 2015. Complete genome analysis of a novel recombinant isolate of Pepper veinal mottle virus from mainland China. Virol. J. 12: 1-4.
35
Potyvirus 에 대한 신규 천연물 유래 물질의
항 바이러스성 특성 구명
배미아
초록
식물 바이러스는 작물의 상품가치를 떨어트리고, 수확량을 감소시켜 전세계적 으로 큰 경제적 손실을 일으킨다. 곰팡이나 세균과 같은 병원균과는 달리 바 이러스의 경우 바이러스 병을 방제할 수 있는 화학적 방제제가 개발되어 있지 않기 때문에 바이러스 벡터를 구제, 감염된 식물을 제거, 바이러스 저항성 품 종 재배 등의 방법으로 병을 예방하고 병의 확산을 막고 있다. Potyvirus는 주요 식물 RNA 바이러스 속 중 하나로, 경제적으로 중요한 작물인 감자나 토마토, 콩과 고추 등에 감염하여 지속적인 피해를 일으키고 있다. 본 연구에 서는, 팜한농으로부터 분양 받은 신규 물질의 항 바이러스성 특성을 구명하고 자 하였다. 식물에 피해를 주는 대표적인 6종의 바이러스(cucumber mosaic virus, pepper mottle virus, pepper mild mottle virus, potato virus X, potato virus Y, tomato spotted wilt virus)를 대상으로 초기 검정을 수행하였다. 담배식물(Nicotiana benthamiana)에 항 바이러스성 물질을 2번 경엽처리 해준
뒤 바이러스의 증식과 병징을 관찰한 결과 약제를 처리한 식물의 상엽에서 대 조구에 비해 potato virus Y와 pepper mottle virus의 증식이 억제되는 것을 관찰되었다. 또한 Potyvirus 속으로 분류되는 chilli veinal mottle virus와
36
soybean mosaic virus 약제를 처리한 식물의 상엽에서 바이러스 증식 억제 효과가 있었으며, 실험 식물인 담배뿐만 아니라 재배 작물인 콩과 고추에서도 바이러스 방제 효과가 있음을 확인하였다. 이후, 이 현상에 어떤 식물 방어 hormone pathway가 관여하는지 알아보고자 qRT-PCR을 통해 저항성 관련 유전자의 발현양을 비교 분석 해 본 결과, 약제를 처리한 식물에서 JA/ET hormone pathway의 EIN2와 PDF1.2의 유전자 발현양이 대조구에 비해 증 가된 것을 확인할 수 있었다. 이러한 결과는 약제 처리 시 JA/ET pathway가 유도되어 potyvirus속에 포함되는 바이러스들에 저항성 반응이 나타난 것이 라고 추측된다. 본 연구를 통해 신규 천연물 유래 물질이 Potyvirus의 감염과 증식을 억제할 수 있음을 확인하였고, 화학적 방제제의 후보 물질로서 개발이 기대된다. 주요어: Potyvirus, 천연물, 항바이러스성 특성, 바이러스 증식 억제, 유도 저 항성, 식물 방어 기작