INTRODUCTION
Microbial antagonists that mediate fungal plant disease suppression is suggested as one of the best alternatives for chemical protection, which is less dependent on chemicals and is environmentally safe. Although several genus of rhizobacteria were reported to have antifungal property, Bacillus sp. are of a great choice since they tolerate extreme heat and desiccation by producing endospores (Handelsman and Stabb 1996; Emmert and Handelsman 1999). The anta-gonistic activity of rhizobacteria towards fungal pathogens is usually related to the production of antifungal compounds
and extracellular hydrolytic enzymes (Fguira et al. 2005; Joo 2005; Mukherjee and Sen 2006). Chitinase and β-1,3-glu-conase are considered to be important hydrolytic enzymes in the lysis of fungal cell walls such as, for example, cell walls of Fusarium oxysporum, Sclerotinia minor, and S. rolfsii (Singh et al. 1999; EI-Tarabily et al. 2000; El-Katatny et al. 2001). Bacillus and related genera also produce peptide antibiotics like bacilopeptins, fusaricidins, iturins, mattacin, and different kinds of polymyxins that play a major role in plant disease suppression (Kajimura et al. 1995; Beatty and Jensen 2002; Martin et al. 2003) as well as post-harvest disease management (Klich et al. 1994). Antifungal meta-bolites of Bacillus sp. are generally polypeptides composed of only amino acids but sometimes contain other residues, such as in the case of rhizocticin A, which is a
phosphono-─ ─ 189 ──
Radiation-Induced Mutagenesis of Antifungal Metabolite
Producing Bacillus sp. HKA-17
Young-Keun Lee and M. Senthilkumar*
Radiation Research Division for Biotechnology, Korea Atomic Energy Research Institute, Jeongeup 580-185, Korea
Abstract -- Bacillus sp. Strain HKA-17, isolated from the surface sterilized root nodule of Glycine
max, inhibited several fungal plant pathogens. It produced a diffusible extracellular antifungal metabolite that was extracted with n-butanol. The crude extract was purified through Superdex™ 75 10/300 GL FPLC column. FT-IR spectrum of the FPLC purified-antifungal metabolite confirm-ed the presence of peptide and glycosidic bonds in its structure. Gamma inducconfirm-ed mutagenesis of HKA-17 was carried out at an LD99dose (8.46 kGy) to generate a mutant library. By screening the mutant library through a duel plate assay with Alternaria alternata, we selected one mutant with enhanced biocontrol activity (HKA-17e1) and two defective mutants (HKA-17d1 and HKA-17d2). Overproducing mutant recorded the largest inhibition zone (16.25±±0.86 mm) compared to any other mutant clone as well as wild type, and could be used as a potential biocontrol agent for plant disease suppression. The effect of HKA-17 antifungal metabolite on hyphal morphology was clearly demonstrated through scanning electron microscopy. The crude extract of defective mutant HKA-17d1 did not induce any changes in hyphal morphology of A. alternata. However, antifungal meta-bolites of HKA-17 induced abnormal hyphal structures such as hyphal shrivelling, the bulging and swelling of intercalary cells, fragmentation, and cell lysis.
Key words : Alternaria alternata, Bacillus, Antifungal metabolite, Gamma irradiation
* Corresponding authors: Senthilkumar Murugesan, Tel. +82-63-570-3304, Fax. +82-63-570-3309, E-mail. [email protected]
oligopeptide (Kluger et al. 1990). Antifungal peptides pro-duced by non-ribosomal peptide synthetase (NRPS) of
Bacillus are reported to have amino acids which are unique
and not commonly found in proteins. Bacillomycin D con-tains D-amino acid residues in addition to L-amino acid residues, while plipastatin B1 contains unusual amino acid residue ornithine (Peypoux et al. 1981; Tsuge et al. 1995). Zwittermicin-producing strains Bacillus cereus UW85, B.
subtilis CL27, and B. pumilus CL45 were potentially used
for the control of Pythium aphanidermatum, Botrytis sp., and
Alternaria brassicola, respectively (Smith et al. 1993;
Lei-fert et al. 1995). Since the production of antifungal metabo-lites, extracellular enzymes, and antibiotics appears to be responsible for plant disease suppression by biocontrol bac-teria, it is hypothesized that superior biocontrol activity might be achieved by increasing production levels of these metabolites. Several successful endeavors have been made to enhance the biocontrol potentiality of Trichoderma spec-ies by exposing the spores to chemical, e.g., ethyl methane sulfonate (EMS), or physical mutagens, e.g., gamma ray (Youssef and Aziz NH 1999). Two mutants, namely Th50M6 and Th50M11, with enhanced biocontrol activity were deve-loped by exposing the wild type T. harzianum conidia to gamma irradiation (Mohamed and Haggag 2006). The bio-control efficiency of Bacillus strains can also be further improved by radiation-induced or chemical mutagenesis. In our laboratory, Bacillus lentimorphus strain WJ5a17 with increased antifungal activity over wild strain WJ-5 was developed through gamma radiation-induced mutagenesis (Lee et al. 2003). The present study was carried out with the aim to develop mutant clones of Bacillus sp. HKA-17 with enhanced as well as defective antifungal activity. We also studied the chemical nature of antifungal metabolite and its effect on Alternaria alternata hyphal morphology.
MATERIALS AND METHODS
1. Bacterial and fungal isolatesBacillus sp. Strain HKA-17 was originally isolated from
a surface-sterilized soybean root nodule (Hung et al. 2007). The isolate is maintained as 25% glycerol stock under -80�C as well as under refrigerated conditions. Fungal phytopatho-gens, as listed in Table 1, are maintained in PDA plates and stored under refrigerated condition.
2. Antagonistic activity of HKA-17 against phytopathogens
Agar discs from the actively growing region of fungal pathogens were collected and placed at the center of the PDA plates. HKA-17 was streak inoculated on the same plate at 2.5 cm away from the fungal disc and incubated at 28±2�C for 3~5 days. The plates were observed for the presence of a fungal growth inhibition zone.
3. FPLC purification and FT-IR spectral analysis
Bacillus sp. HKA-17 was grown in one liter of potato dex-trose broth for 48 h, 200 rpm at 28±2�C. The culture super-natant was prepared by centrifugation at 5,000 rpm for 20 min and added with an equal volume of n-butanol. The aqueous phase as well as butanol extract was tested for anti-fungal activity with Alternaria alternata. Active crude extract in butanol was dried completely and dissolved in HPLC-grade methanol. Fast Protein Liquid Chromatography (FPLC) with Superdex™ 75 10/300 GL (GE Healthcare, Sweden) was used to purify the crude extract. The sample was eluted with a buffer containing 150 mM of Tris-Cl (pH 7.0) and 50 mM of NaCl at a flow rate of 0.5 ml min-1. 500 μl fractions corresponding to each peak were collected, pooled together, and dialyzed with water. Fractions were tested for antifungal activity and then lyophilized. The FPLC purified active frac-tion was pelletized with potassium bromide (KBr). Pellets were subjected to spectral analysis in transmission mode between wavelengths of 4,000-400 cm-1using a resolution of 4 cm-1/20-kHz scan speed in FT-IR spectrophotometer.
4. Radiation sensitivity
Cell pellets were collected by centrifuging 20 ml of a log phase bacterial isolate cultured in the LB broth. Pellets were
Table 1. Antifungal activity of HKA-17 against phyto-pathogens
Fungus Inhibition zone (mm)
Alternaria alternata 11.25±0.96 Alternaria solani 10.75±0.50 Botrytis cinerea 8.50±1.29 Cladosporium sp. 10.25±0.96 Colletotrichum gloeosporioides 8.00±0.82 Fusarium oxysporum 8.50±0.58 Phytophthora capcisi 7.00±0.82 Pythium ultimum 7.25±0.96 Rhizoctonia solani 11.50±0.58 Sclerotinia sclerotiorum 9.75±0.96
washed twice with sterile-distilled water and suspended (~107~109CFU ml-1). 500 μl of cell suspensions were
transferred to 1.5 ml micro-centrifuge tubes and irradiated at different doses expressed in kGy, as shown in Fig. 2. Sam-ples were irradiated in a cobalt-60 irradiator (capacity: 250,000 Ci, dose rate 920 Gy hr-1, AECL) at the Advanced
Radiation Technology Institute, Korea Atomic Energy Research Institute, South Korea. The bacterial population in irradiated samples was determined by serial dilution and plate count method. The D10-value was determined by
plot-ting a radiation dose (kGy) on the X-axis and a survival ratio on the Y-axis. The negative reciprocal of the slope indicated the D10-value. LD99value, the radiation dose required to
kill 99% of viable bacterial cells was determined by using the formula LD99==log (0.01)/slope.
5. Radiation mutagenesis and selection
To induce the mutagenesis, bacterial cell suspensions were prepared as described above and exposed to gamma irradia-tion at the dose of LD99. Irradiated samples were serially diluted and plated on NA agar and incubated at 28±2�C for 24 h. Master plates were prepared from the mutant clones and stored under refrigerated conditions. The antifungal activity of mutant clones was determined by duel plate assay with A. alternata on PDA plates. Overproducing as well as antifungal activity defective mutants were selected and stored as pure cultures.
6. Scanning electron microscopy
Mycelium of A. alternata was treated with a crude
antifun-gal metabolite with a concentration of 10 mg ml-1of water. Hyphae were removed after 5 h of incubation, fixed in glutaraldehyde buffer for 4 h in room temperature, dehy-drated in graded alcohol, and dried. Gold sputter-coating was carried out under reduced pressure in an inert argon gas atmosphere (Agar sputter coater P7340). After sputter-coat-ing, specimens were examined under a scanning electron microscope (JSM 6390) operated at 30 kV.
RESULTS AND DISCUSSION
Bacillus sp. Strain HKA-17, isolated from surface steriliz-ed root nodules of Glycine max, showsteriliz-ed an inhibitory effect on the growth of Rhizoctonia bataticola (Senthilkumar et al. 2008). We subsequently tested its antifungal activity against several phytopathogens belonging to oomycetes, ascomy-cetes, and basidiomycetes. The hyphal growth of all test fungi was inhibited by HKA-17. The extent of inhibition
Fig. 1. Scanning electron micrograph of Bacillus sp. HKA-17.
Fig. 2. Radiation sensitivity of Bacillus sp. HKA-17.
Fig. 3. FT-IR spectrum of the crude antifungal metabolite of
Bacil-lus sp. HKA-17. Radiation sensitivity kGy 0 2 4 6 8 10 12 14 16 18 Log (Nx/No) 0 -1 -2 -3 -4 -5 -6 -7 y==-0.2364x-2.7804 R2==0.9725 Transmittance (%) 100.0 99.5 99.0 98.5 98.0 97.5 97.0 96.5 3500 3000 2500 2000 1500 1000 Wavenumber cm-1
varied depending on the fungal pathogen (Table 1). Pythium
ultimum and Phytophthora capsici are less sensitive to the
antifungal metabolites of HKA-17, while Rhizoctonia solani and A. alternata are highly sensitive and recorded maximum growth inhibition zone. The result also indicated that the antifungal metabolites are diffusible and extracellular in nature, which was confirmed with the antifungal activity of filter-sterilized, 48 h-old culture supernatant. Antifungal
Table 2. FT-IR spectrum of the crude antifungal extract
Frequency (cm-1) Possible assignment
3432 NH2stretching (amines), H-bonded OH groups
2924 Aliphatic C-H stretching
2854 Acyl group
1760~1670 C==O, carboxylic acid, aldehyde, ketones 1660~1535 NH2bending, C==O, C==N stretching
1074 C-O, C-C stretching, C-O-H,
C-O-C deformation (glycopeptide)
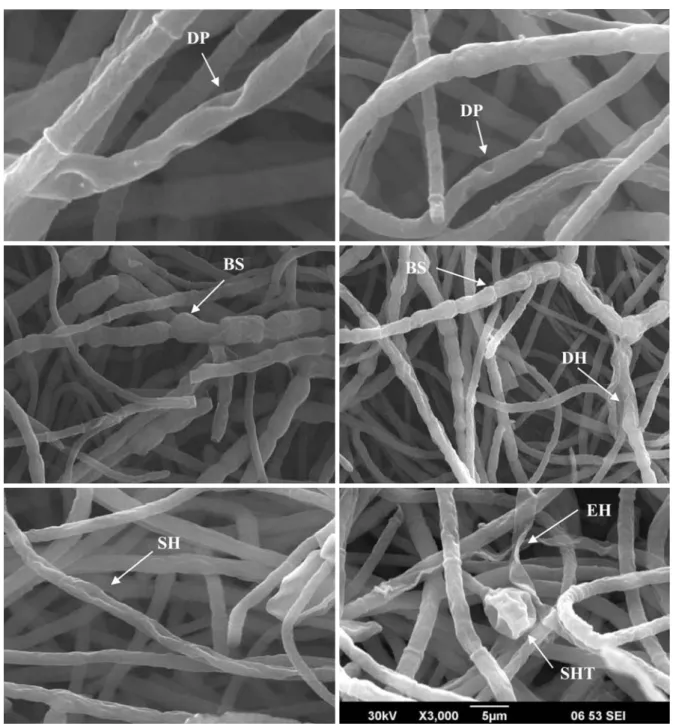
Fig. 4. Crude antifungal metabolite induced morphological changes on A. alternata hyphae. DP-depressions on hyphae, BS-bulging and swel-ling of intercalary cells, DH-degrading mycelium, SH-shriveswel-ling hyphae, EM-emptied hyphae due to lysis, and SHT-swelswel-ling of a hyphal tip.
metabolite of 48 h-old culture supernatant was extracted in n-butanol and concentrated to dryness. The crude extract was dissolved in HPLC-grade methanol and subjected to FPLC for further purification. Fractions corresponding to the 7th
peak showed the antifungal activity against A. alternata. Active fractions were pooled together, dialyzed, and then lyophilized. Functional groups and chemical bonds present in the FPLC purified fraction was analyzed through FT-IR. The major peaks were observed at wave numbers of 3,432, 2,924, 1,639, and 1,074 cm-1(Fig. 3). Possible functional
groups or bonds corresponding to the peak wave numbers are listed in Table 2. Broad peak at 3,432 cm-1and 1,660~
1,535 cm-1corresponding to the NH
2stretching and bending,
and the C==O and C==N stretching, respectively, indicated the peptide nature of the antifungal metabolite. A peak at a wave number of 1,074 cm-1indicated the presence of sugar
moiety/glycosidic bond. Similar kind of antifungal metabo-lite was characterized from Paenibacillus lentimorbus WJ5 (Lee et al. 2007).
Gamma-induced mutagenesis, a common tool to alter bio-control efficiency (Vaidya et al. 2003; Gohel et al. 2004; Mohamed and Haggag 2006), was used to develop enhanced and defective mutants. Exposing a wild-type culture of
Tri-choderma harzianum to gamma irradiation induced two
stable salt-tolerant mutants namely, Th50M6 and Th50M11. Gamma mutant Th50M6 displayed a maximum antagonistic activity against F. oxysporum f. sp. lycopersici on either nat-ural or saline media compared to the wild strain that exhibit-ed a distinct decrease in its biological control activity on the saline medium than in the natural one by 60 and 50%, res-pectively. At the same time, F. oxysporum was completely inhibited when amended with 50% culture filtrate of Th50M6 and Th50M11, compared with 74.5 and 12.6% of the wild type at the natural and saline medium, respectively (Moham-ed and Haggag 2006). Alcaligenes xylosoxydans EMS 33, a hyperchitinase mutant that produced 3.4 fold more chitinase than the wild type was developed initially through UV irra-diation followed by EMS mutagenesis (Vaidya et al. 2003). We initially studied the radiation sensitivity of HKA-17. The D10 and LD99values of HKA-17 were determined as 4.23
kGy and 8.46 kGy, respectively. A mutant library was deve-loped by exposing HKA-17 to gamma rays at the LD99value
(8.46 kGy). By screening approximately 700 clones from the mutant library through duel-plate assay with A. alternata, we selected one mutant with enhanced biocontrol activity
and two defective mutants. Overproducing mutant recorded the largest inhibition zone (16.25±0.86 mm) compared to any other mutant clones as well as wild type and could be used as potential biocontrol agent for plant disease suppress-ion. Defective clones were designated as HKA-17d1 (clone No. 54) and HKA-17d2 (clone No. 368), while the mutant with enhanced activity was HKA-17e1 (clone No. 148).
Defective mutants that no longer produce antifungal meta-bolite were used indirectly to prove the role of antifungal metabolite on the suppression of fungal growth as well as its effect on hyphal morphology through scanning electron microscopy. Crude extracts of defective mutant HKA-17d1 did not induce any changes in the hyphal morphology of A.
alternata. However, antifungal metabolites of HKA-17
induced abnormal hyphal structures such as thickness, bul-bous roundedness of treated hyphae, or hyphal tip swelling (Fig. 4). Hyphal fragmentation shrivelling, the bulging and swelling of intercalary cells, and lysis were also observed. Similarly, culture filtrate of Pseudomonas aeruginosa K-187 induced growth aberration, hyphal swelling, and lysis of many fungi due to its high content of chitinase enzyme. In conclusion, we demonstrated the role of antifungal metabo-lite on hyphal growth inhibition and a mutant clone Bacillus sp. HKA-17e1 with enhanced biocontrol activity could be used for effective plant disease management.
ACKNOWLEDGMENT
This study was carried out under the Nuclear R&D Pro-gram of the Ministry of Education, Science and Technology, Republic of Korea.
REFERENCES
Beatty PH and Jensen SE. 2002. Paenibacillus polymyxa pro-duced fusaricidin type antifungal antibiotics active against
Leptosphaeria maculans, the causative agent of blackleg
disease of canola. Can. J. Microbiol. 48:159-169.
El-Katatny MH, Gudelj M, Robra KH, Elnaghy MA and Gübitz GM. 2001. Characterization of a chitinase and an endo-β-1,3-glucanase from Trichoderma harzianum rifai T24 invo-lved in control of the phytopathogen Sclerotium rolfsii.
Appl. Microbiol. Biotechnol. 56:137-143.
Sivasithamparam K, McKenna F and Hardy GE. 2000. Bio-logical control of Sclerotinia minor using a chitinolytic bac-terium and actinomycetes. Plant Pathol. 49:573-583. Emmert EAB and Handelsman J. 1999. Biocontrol of plant
disease: a (Gram) positive perspective. FEMS Microbiol.
Lett. 171:1-9.
Fguira LF, Fotso S, Ameur-Mehdi RB, Mellouli L and Laatsch H. 2005. Purification and structure elucidation of antifungal and antibacterial activities of newly isolated Streptomyces sp. strain US80. Res. Microbiol. 156:341-347.
Gohel V, Megha C, Vyas P and Chhatpar HS. 2004. Strain im-provement of chitinase producing isolate Pantoea dispersa for enhancing its biocontrol potential against fungal plant pathogens. Annals of Microbiol. 54:503-515.
Handelsman J and Stabb EV. 1996. Biocontrol of soil borne plant pathogens. Plant Cell. 8:1855-1869.
Hung PQ, Senthilkumar M, Govindasamy V and Annapurna K. 2007. Isolation and characterization of endophytic bacteria from wild and cultivated soybean varieties. Biol. Fertil.
Soil. 44:155-162.
Joo GJ. 2005. Production of an anti-fungal substance for bio-logical control of Phytophthora capsici causing
phytophtho-ra blight in red-peppers by Streptomyces halstedii. Biote-chnol Lett. 27:201-205.
Kajimura Y, Sugiyama M and Kaneda M. 1995. Bacilopeptins, new cyclic lipopeptide antibiotics from Bacillus subtilis FR-2. J. Antibiot. 48:1095-1103.
Klich MA, Arthur KS, Lax AR and Bland JM. 1994. Iturin A: a potential new fungicide for stored grains. Mycopathologia.
127:123-127.
Kluger M, Loeffler W, Rapp C, Keen A and Jung G. 1990. Rhizocticin A, an antifungal phosphono-oligopeptide of
Bacillus subtilis ATCC 6633: biological properties. Arch. Microbiol. 153:276-281.
Lee YK, Kim JS, Jang YS and Lee HY. 2003. Biological con-trol of Pythium root rot by radiation induced mutant of
Bacil-lus lentimorbus WJ5a17. Korean J. Environ. Biol.
21:276-285.
Lee YK, Senthilkumar M, Kim JH, Swarnalakshmi K and Annapurna K. 2008. Purification and partial characterization of antifungal metabolite from Paenibacillus lentimorbus WJ5. World J. Microbiol. Biotechnol. 24:3057-3062. Leifert C, Li H, Chidburee S, Hampson S, Workman S, Sigee D,
Epton HAS and Harbour A. 1995. Antibiotic production and biocontrol activity by Bacillus subtilis CL27 and Bacillus
pumilus CL45. J. Appl. Bacteriol. 78:97-108.
Martin NI, Hu H, Moake MM, Churey JJ, Whittal R, Worobo RW and Vederas JC. 2003. Isolation, structural characteri-zation, and properties of mattacin (polymyxin M), a cyclic peptide antibiotic produced by Paenibacillus kobensis M.
J. Biol. Chem. 278:13124-13132.
Mohamed HAL and Haggag WM. 2006. Biocontrol potential of salinity tolerant mutants of Trichoderma harzianum against Fusarium oxysporum. Brazilian J. Microbiol. 37: 181-191.
Mukherjee G and Sen SK. 2006. Purification, characterization and antifungal activity of chitinase from Streptomyces
vene-zuelae P10. Curr. Microbiol. 53:265-269.
Peypoux F, Besson F, Michel G and Delcambe L. 1981. Struc-ture of Bacillomycin D, a new antibiotic of the iturins group.
Eur. J. Biochem. 118:323-327.
Senthilkumar M, Swarnalakshmi K, Govindasamy V, Lee YK and Annapurna K. 2009. Biocontrol potential of soybean bacterial endophytes against charcoal rot
fungus-Rhizocto-nia bataticola. Curr. Microbiol. 58:288-293.
Singh PP, Shin YC, Park CS and Chung YR. 1999. Biological control of Fusarium wilt of cucumber by chitinolytic bacte-ria. Phytopathol. 89:92-99.
Smith KP, Havey MJ and Handelsman J. 1993. Suppression of cottony leak of cucumber with Bacillus cereus strain UW85.
Plant Dis. 77:139-142.
Tsuge K, Ano T and Shoba M. 1995. Characterization of
Bacil-lus subtilis YB8, coproducer of lipopeptides surfactin and
plipastatin B1. J. Gen. Appl. Microbiol. 41:541-545. Vaidya RJ, Macmil SLA, Vyas PR and Chhatpar. 2003. The
novel method for isolating chitinolytic bacteria and its appli-cation in screening for hyperchitinase producing mutant of
Alcaligenes xylosoxydans. Lett. Appl. Microbiol.
36:129-134.
Wang SL, Yieh TC and Shih IL. 1999. Production of antifungal compounds by Pseudomonas aeruginosa K-187 using shrimp and crab shell powder as a carbon source. Enzyme
Microb. Technol. 25:142-148.
Youssef BM and Aziz NH. 1999. Influence of gamma-irradia-tion on the bioconversion of rice straw by Trichoderma
viride into single cell protein. Cytobiol. 97:171-183.
Manuscript Received: August 17, 2009 Revision Accepted: September 2, 2009