저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
이학 석사학위 논문
초파리 PERI
OD 단백질의
아미노산 926-977이 생체시계의
작동에 관여하는 분자적 기전
아 주 대 학 교
대 학 원
의생명과학과/
신경과학전공
선 우 철
초파리 PERI
OD 단백질의
아미노산 926-977이 생체시계의
작동에 관여하는 분자적 기전
지도교수
김 은 영
이 논문을 이학 석사학위 논문으로 제출함.
2010년
8월
아 주 대 학 교
대 학 원
의생명과학과/
신경과학전공
선 우 철
선우철의 이학 석사학위 논문을 인준함.
심사위원장
김
은
영
(
서명)
심 사 위 원
주
일
로
(
서명)
심 사 위 원
이
상
윤
(
서명)
아 주 대 학 교
대 학 원
2010년 6월 25일
- 국문요약 -
초파리 PERIOD 단백질의 아미노산 926-977 이 생체시계의
작동에 관여하는 분자적 기전
생체시계의 분자적 기전은 transcriptional-translational feedback loop 이다. 초파리 feedback loop 의 첫 번째 loop 에서는 bHLH/PAS 도메인을 함유한 전사인자인 dCLOCK (dCLK) 과 CYCLE (CYC) 가 dperiod (dper)와 timeless (tim) 의 발현을 촉진하고 합성된 dPER 단백질과 TIM 단백질이 이합체로 결합하여 dCLK/CYC 의 전사활성을 억제하여 dper 와 tim 의 발현이 하루를 주기로 진동케 한다. 여기에는 dPER 에 의한 dCLK/CYC 전사활성의 억제가 필수적이다. 본 연구에서는 dPER 에서 dCLK/CYC 의 전사활성을 억제하는데 필요한 도메인으로 알려진 dCLK CYC inhibition domain (CCID) 에서 특히 종간 보존성이 매우 높은 아미노산 926-977 부분의 역할을 탐구하였다. dPER 로부터 아미노산 926-977 을 제거한 변이체인 dPER(Δ926-977) 은 초파리 세포라인 Schneider 2 (S2) 세포에서 dCLK/CYC 의 전사활성을 억제하지 못하였다. dPER(Δ926-977) 는 dCLK 과 결합하지 못하였는데, 이로부터 아미노산 926-977 이 dCLK 의 binding domain 으로 작용한다는 사실을 밝혀내었다. 더욱이 아미노산 949-977 만을 내부적으로 제거한 dPER(Δ949-977) 가 S2 세포에서 dCLK/CYC 의 전사활성을 억제하지 못하고 dCLK 과의 결합능력이 떨어지는 것으로 보아 아미노산 949-977 이 dCLK 과의 결합에 필요한 최소 도메인인 것을 확인하였다. 아미노산 926-977 의
in vivo 역할을 탐색하기 위하여 dPER(Δ926-977)를 발현시키는 초파리를 제작하였고, 이 초파리의 생체리듬을 분석하여 본 결과 주기가 길어지기는 하였으나 정상적인 생체리듬을 갖는 것을 알 수 있었다. dCLK 의 결합에 필수적인 아미노산 926-977 이 없는 dPER 가 in vivo 에서 생체시계 유전자의 발현이 진동하게 하는 feedback loop 작동에 정상적인 기능을 한다는 사실은 매우 흥미로우며 이에 대한 후속 연구가 진행되어야 할 것으로 보인다. 핵심어 : 초파리, 생체시계, dPERIOD, dCLOCK, 전사활성, 인산화
차 례
국문요약 ··· ⅰ 차례 ··· ⅲ 그림 차례 ··· ⅴ 표 차례 ··· ⅶ Ⅰ. 서론 ··· 1 Ⅱ. 재료 및 방법 ··· 8 A. Plasmid ··· 81. Site-direct mutagenesis 를 이용하여 만든 plasmid 들 ··· 8
2. Rapid and efficient PCR-based mutagenesis 를 이용하여 만든 plasmid 들 (Ko 와 Ma, 2005) ··· 8
3. Cloning 으로 만든 plasmid 들 ··· 8
4. 참고문헌을 통해 이미 사용되어 인용된 plasmid 들 ··· 8
B. Site-direct mutagenesis ··· 9
1. QuickChange site-directed mutagenesis kit ··· 9
2. Site-directed mutagenesis 를 위한 primer ··· 10
3. Rapid and efficient PCR-based mutagenesis method (Ko 와 Ma, 2005) ··· 11
4. PCR-based Rapid site directed mutagenesis 를 위한 Primer 들 ··· 13
C. Drosophila melanogaster Schneider line 2 (S2) 세포의 배양과 transfection ··· 15
D. 형질전환 초파리 (Transgenic Drosophila melanogaster) ··· 16
E. Locomotor activity assays ··· 17
F. Immunoblot analysis ··· 18
G. Immunoprecipitation (IP) ··· 21
H. Luciferase Reporter Assay 와 β-Galactosidase Enzyme Assay System ··· 23
Ⅲ. 결과 ··· 25 A. S2 세포에서 dPERΔ926-977 은 dCLK 의 전사활성을 억제하지 못한다 ··· 25 B. S2 cells 에 서 dPERΔ926-977 은 dCLK 과 결 합 하 지 못 한 다 ··· 28 C. dPERΔ926-977 은 DBT 에 의한 인산화에 결함을 나타내었다 ··· 32 D. 아미노산 949-977 이 dCLK 과의 결합에 필요한 최소 도메인이다 ··· 34
E. dPER 의 아미노산 949-977 를 Ser 과 Thr 을 모두 Ala 으로 바꾸어도 DBT 에 의한 인산화가 일어난다 ··· 38 F. dPERΔ926-977 을 발현하는 형질전환 초파리는 주기가 길어지기는 하였으나 정상적인 생체리듬을 가졌다 ··· 43 Ⅳ. 고찰 ··· 46 Ⅴ. 결론 ··· 48 참고문헌 ··· 50 ABSTRACT ··· 53
그림 차례
Fig. 1. The SCN sends signals throughout the body in response to light and dark ··· 5
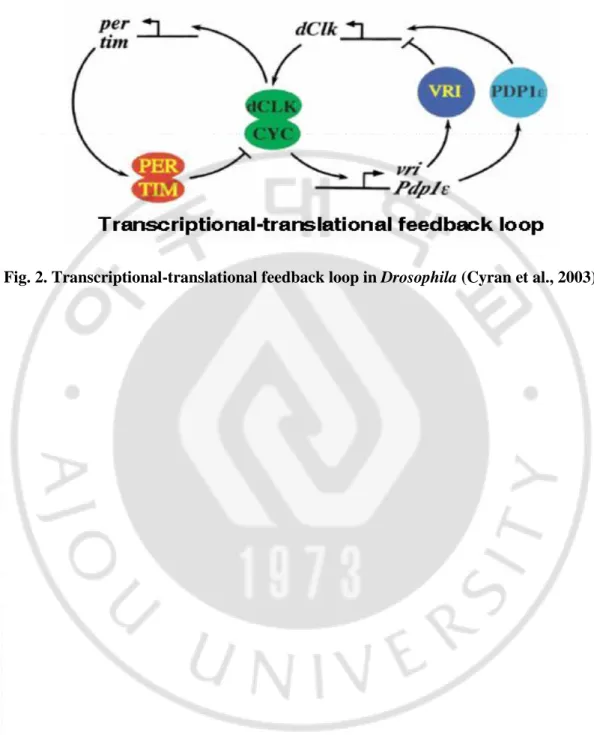
Fig. 2. Transcriptional-translational feedback loop in Drosophila ··· 6
Fig. 3. Sequence alignment of dPER protein from different species of Drosophila ··· 7
Fig. 4. Augmentataion of nuclear localization of dPERΔ926-977 could not restore the repressor activity of dPER(Δ926-977) ··· 27
Fig. 5. dPER(Δ926-977) could not directly bind to dCLK in S2 cells ··· 30
Fig. 6. DBT dependent phosphorylation of dPER(Δ926-977) is attenuated in S2 cells ··· 33
Fig. 7. dPER(Δ926-977) is a very weak repressor of dCLK-dependent transcription in S2 cells ··· 37
Fig. 8. Deletion of aa 949 to 977 from dPER strongly attenuates DBT-dependent hyperphosphorylation of dPER in S2 cells ··· 40
Fig. 9. dPER(Δ949-977) could not directly bind to dCLK in S2 cells ··· 41
Fig. 10. All 9 serines and threonines in dPER aa 949-977was switched to alanines ···· 42
Fig. 11. DBT-dependent phosphorylation and degradation of dPER(949-977_9A) occur in the similar extent as those of wild-type dPER in S2 cells ··· 44
표 차례
Table. 1. Locomotor activity rhythms of p{ dper(926-977)} flies and control p{dper(WT)} transgenic flies ··· 36
I. 서 론
지구상에 존재하는 모든 생명체는 지구의 자전으로 변화하는 환경을 미리 예측할 수 있다. 예를 들면 포유류의 경우 뇌의 시상하부에 위치한 SCN (Suprachiasmatic Nuclei) 이 낮과 밤을 예측하고 각 장기의 다양한 세포로 필요한 정보를 보내어 하루 중 대사, 체온, 호르몬 및 행동 등을 조절한다 (Fig. 1). 이로 인해 하루를 주기로 생명체의 여러 생리작용과 행동 및 대사 패턴을 변화시키는 생체리듬이 나타내게 된다. 생체시계와 질병과의 상관관계는 암, 수면장애, 여러 가지 형태의 우울증, 신경퇴행성 질환, 대사증후군, 약물중독 등 무수히 많은 예로부터 중요성이 입증되고 있다 (Bunney 와 Bunney, 2000; Cardinali, 2000; Fu 와 Lee, 2003; McClung, 2007; Rudic 등, 2004). 따라서 생체시계의 작용기전을 밝혀 생체리듬을 일으키는 원인과 비정상적 생체리듬이 발생함으로 인해 어떠한 질병이 나타나며, 예방과 치료를 위한 수단과 방법을 탐색하는데 중요한 단서가 될 것이다. 생체시계는 미생물에서부터 포유류까지 지구상의 모든 생물이 가지고 있는데 기원전부터 과학자들은 생물 내부에 자체적인 시계를 가지고 있을 것이라고 생각하고 있었으며, 이에 관한 연구는 초파리를 동물모델로 이용하면서 활발히 진행되었다. 생체시계의 핵심기전은 “interlocked transcriptional-translational feedback loop" 으로써 두 개의 loop 이 서로 맞물려 생체시계의 핵심 유전자들의 발현이 하루를 주기로 진동할 수 있도록 한다.초파리에서는 dperiod (dper) /timeless (tim) loop 과 dClock (dClk) loop 이 두 개의 feedback loops 을 이루고 있다 (Fig. 2). 이 때 dCLK/CYC 단백질은 이형이합체를 이루어 dper/tim 유전자의 promotor 부분의 E-box 에 붙어 전사를 촉진하고 dPER/TIM 단백질은 이형이합체를 이루어서 자신의 유전자를 만들게 하는 dCLK/CYC 단백질의 활성을 저해하는 feedback loop 를 만들게 된다. 또한 dCLK/CYC 단백질 이형이합체는 vrille (vri)과 PAR domain protein 1ε (PDP1ε) 유전자의 E-box 에 결합하여 이들의 발현을 촉진하는데, 이 때 VRILLE (VRI)은
dClk 의 발현을 억제하고, 반면 PDP1ε은 dClk 의 발현을 촉진하는 또 다른
feedback loop 를 이루게 된다 (Cyran 등, 2003)
생체시계의 “interlocked transcriptional-translational feedback loop”에서 dPER 와 TIM 의 이형이합체가 dCLK 과 CYC 이형이합체의 활성을 억제한다는 것은 잘 알려져 있다. 이때 dPER-TIM 이형이합체 단백질이 전사인자인 dCLK-CYC 이형이합체 단백질을 억제하기 위해 직접적인 결합이 필요할 것이며 (Lee 등, 1998; Lee 등, 1999; Sato 등, 2006), Chang 등은 dPER 가 단독으로도 dCLK-CYC 의 활성을 저해할 수 있음을 보고하여 dCLK-CYC 이합체의 활성을 저해하는 주요 단백질은 dPER 임을 밝혔다 (Chang 과 Reppert, 2003). Chang 등은 이 연구에서 dPER 에서 dCLK 의 활성을 억제하는데 필요한 최소 도메인을 탐색하였고, 그 결과 dPER 에서 아미노산 764-1034 가 내부적으로 제거되었을 때 dCLK 의 전사활성이 억제되지 않았고 이로부터 Chang 등은 dPER 의 아미노산 764-1034 를 dCLK:CYC Inhibition Domain (CCID)라 명명 하였다 (Chang 과 Reppert, 2003).
여러 초파리 종으로부터 CCID domain 의 단백질 서열을 비교 분석해 보면 종간 보존성이 높은 부위 (Conserved region) 와 종간 보존성이 상대적으로 높지
않은 부위 (Non-conserved region) 로 나뉘는 것을 알 수 있다. 특히 종간 보존성이 높은 부위인 C3 와 C4 부위는 dPER 단백질의 기능에 중요한 역할을 할 것이라는 것을 암시한다 (Fig. 3). 아미노산 755-809 부위가 dPER 가 전사 억제제로서 작용하는데 중요한 역할을 하는 기전이 지도교수의 이전 연구에 의해 밝혀졌다. 그 내용을 간략하게 살펴보면 아미노산 755-809 가 없는 dPER(Δ755-809) 는 dCLK 의 활성을 억제하지 못하였으며, 행동학적 생체리듬을 갖지 못하였고, 하루 동안 정상적인 dPER 에 비해 양이 증가하며, dPER 단백질의 인산화에 따른 분해가 거의 없어 진동 없이 일정한 것을 알 수 있었다. 이 때, dPER(Δ755-809) 가 인산화 되지 못하는 이유는 아미노산 755-809 부위가 DBT 와 결합하는 부위이기 때문이라는 사실을 in vitro 와 in vivo 실험에서 알 수 있었으며 (data not shown), dPER 단백질에 의해 dCLK 이 주로 인산화에 의한 전사 후 조절이 많은 기여를 할 것이라는 가능성이 제시됨으로써 dPER 가 dCLK-CYC 의 전사활성을 억제함에 있어 직접적인 결합뿐만 아니라 또 다른 매개 단백질을 통해 보다 간접적인 방법으로 억제할 것이라는 가설이 제시되었다 (Kim 등, 2007)
따라서 본 연구에서는 생체시계의 “interlocked transcriptional-translational feedback loop”에서 dPER 가 dCLK 의 활성을 억제하는데 필요한 CCID 중 종간
보존성이 높아 중요성이 높은 아미노산 926-977 도메인의 기능을
분자적·생화학적 관점에서 탐구하고자 한다. 또한 최근 우리 연구실에서는 luciferase-reporter assay 를 통해 앞서 연구된 dPER CCID 에 존재하는 아미노산 768-842 외에도 아미노산 926-977 역시 제거하게 되면 전사 억제제로서의 활성을 잃어버리게 된다는 연구결과를 얻게 되었는데 이것은 아미노산 926-977 역시
CCID 의 활성에 매우 중요한 기능을 담당하고 있다는 것을 알 수 있으며, dCLK 과의 상호작용과 관련된 메커니즘을 in vitro 와 in vivo, 그리고 행동학적 실험을 통해 분자적·생화학적 관점에서 탐구하고자 하였다.
Fig. 1. The SCN sends signals throughout the body in response to light and dark. (http://learn.genetics.utah.edu/content/begin/dna/clockgenes/)
Ⅱ. 재료 및 방법
A. Plasmid
1. Site-direct mutagenesis 를 이용하여 만든 plasmid 들
13.2dper+(Δ926-977)-HA10His (Lee 등, 1998), dper(Δ926-977),
pAct-dper(Δ926-977)-V5, pAct-dper(Δ926-948), pAct-dper(Δ926-948)-V5,
pAct-dper(Δ949-977), pAct-dper(Δ949-977)-V5, pAct-dper(Δ978-999)-V5
2. Rapid and efficient PCR-based mutagenesis 를 이용하여 만든
plasmid 들 (Ko 와 Ma, 2005)
pAct-dper(949-977_9A), pAct-dper(949-977_9A)-V5
3. Cloning 으로 만든 plasmid 들
pAct-dper(1-1224)-NLS, pAct-dper(Δ926-977)-NLS
4. 참고문헌을 통해 이미 사용되어 인용된 plasmid 들
(A) pMT-dbt-V5, pAct-dper(1-1224)와 pAct-dper(1-1224)-V5 (Ceriani 등, 2002; Kim 과 Edery, 2006; Kim 등, 2007; Ko 등, 2002)
(B) pAct-dper(Δ755-809)-V5 (Kim 등, 2007)
(D) pMTB-HAdclk (Kim 과 Edery, 2006; Ko 등, 2002; Lee 등, 1999) (E) dperE-Luc, pAct5.1-LacZ-V5/His (Darlington 등, 1998)
B. Site-direct mutagenesis
1. QuickChange site-directed mutagenesis kit (Stratagene Co.,
La Jolla, CA, U.S.A)
QuickChange site-directed mutagenesis kit (Stratagene Co., La Jolla, CA, U.S.A) 을 사용하여 site directed mutagenesis 를 수행하였다. 이 때 PfuTurbo DNA polymerase (Stratagene Co., La Jolla, CA, U.S.A) 를 이용하였으며, 각 plasmid 에 대한 PCR 반응조건은 PfuTurbo DNA polymerase (Stratagene Co., La Jolla, CA, U.S.A) 가 제시하는 사용방법으로 설정하였고 PCR 반응 혼합액 구성요소는 100 ng 의 pSP72-dper, 5 ㎕ 의 10X cloned Pfu reaction buffer, 2 ㎕ 의 10 mM dNTP, 각각 150 ng 의 primer (F) (R), 2.5 units 의 PfuTurbo DNA polymerase (Stratagene Co., La Jolla, CA, U.S.A), 그리고 멸균수로 최종반응 양을 50 ㎕ 로 맞추어 조제하였다. PCR 은 핵산증폭기 (MJ Research, Inc., Waltham, MA, U.S.A) 를 이용하였다. PCR 산물 중 mutation 되지 않은 주형 DNA 를 제거하기 위해 제한효소 DpnⅠ 1 ㎕ 를 처리한 후 핵산증폭기를 이용하여 37℃ 에서 1 시간 반응한 후, XL10-Gold Ultracompetent cells (Stratagene Co., La Jolla, CA, U.S.A) 에 42℃ 로 30 초간 heatshock 을 주어 transformation 하였다. 재조합 plasmid 를 가진 colony 임을 확인하기 위해 다시 핵산증폭기를 사용하여 colony PCR 을 수행하였다. Colony 현탁액은 20 ㎕ 의 멸균수에 단일 colony 를 넣고 잘 섞어 준비하였다. Colony PCR 을 위한 반응혼합액은 10 ㎕ 의 colony 현탁액, 2.5 ㎕ 의 10X PCR buffer, 1 ㎕ 의 10 mM
dNTP, 1 ㎕ 의 rTaq (Takara bio Inc., Otsu, Shiga, Japan) 그리고 최종 반응량을 25 ㎕ 로 만들기 위해 8.5 ㎕ 의 멸균수를 넣어 조제하였다. 반응조건은 95℃ 2 분간 최초 세포막 파괴 및 DNA 를 denature 하고, 95℃ 30 초, 55℃ 30 초, 72℃ 60 초 세 가지 온도를 순서대로 denature, annealing, elongation 30 회 반복 반응하였고, 72℃ 10 분간 최종 신장반응 하였다. Colony PCR 산물을 KpnⅠ과 BamHⅠ으로 자른 후 2% agarose gel 에서 전기영동 하여 예상 크기와 근접한 colony 를 DNA-spin plasmid DNA purification kit (Intron Biotechnology Inc., Seongnam, Gyeonggi-do, Korea) 으로 mini prep 한 후 DNA 염기서열 분석 (Bionics Co., Seoul, Korea) 으로 돌연변이 된 plasmid 들을 최종 확인하였다. 최종 확인된 dperΔ 들을 KpnⅠ, BamHⅠ과 XhoⅠ을 이용하여 pSP72 vector 에서 기존에 연결되어 있던 Casper vector 또는 pAc5/V5-His vector 로 다시 잘라 옮겨 붙인 후 JM109 competent cells (Promega Co., Madison, WI, U.S.A) 에 42℃ 47 초간 heatshock 을 주어 transformation 하였다. 원하는 plasmid 를 가진 colony 를 확인하기 위해 KpnⅠ과 BamHⅠ으로 자른 후 전기영동 하여 확인하였고, Maxi prep kit (Qiagen Co., Hilden, Germany) 으로 DNA 를 다량 확보하였다.
2. Site-directed mutagenesis 를 위한 primer
(A) dperΔ926-977(F)
5‟-TCC ACG CCA GGA CCC ACG CCC ACC AGT CCC -3‟ (B) dperΔ926-977(R)
(C) dperΔ926-948(F)
5‟-TCC ACG CCA GGA CCC GTT CAC TCC ACG CAC-3‟ (D) dperΔ926-948(R)
5‟-GTG CGT GGA GTG AAC GGG TCC TGG CGT GGA-3‟ (E) dperΔ949-977(F)
5‟-GGC ATC ACA CCG CCC ACG CCC ACC AGT CCC-3‟ (F) dperΔ949-977(R)
5‟-GGG ACT GGT GGG CGT GGG CGG TGT GAT GCC-3‟
3. Rapid and efficient PCR-based mutagenesis method (Ko 와
Ma, 2005)
pAct-dper-V5 내부에 일부 인산화되는 아미노산인 Ser 과 Thr 을 Ala 으로 치환하기 위해 해당 염기서열을 Rapid and efficient PCR-based mutagenesis 방법을 사용하였다. (Ko 와 Ma, 2005). 이 때 PfuTurbo DNA polymerase (Stratagene Co., La Jolla, CA, U.S.A) 를 이용하였으며, primer 는 Ⅱ형 제한효소 BsmBⅠ염기를 포함하며 치환하고자 하는 특정부위 염기서열을 바꿔서 제작 한 후 함께 해당
주형 DNA 와 함께 BamHⅠ염기서열 부위부터 XbaⅠ염기서열 부위까지
핵산증폭기 (MJ Research, Inc., Waltham, MA, U.S.A) 를 이용하여 PCR 을 수행하였다. 이후 BsmBⅠ으로 55℃에서 8 시간 동안 자른 후 T4 DNA ligase (Takara bio Inc., Otsu, Shiga, Japan) 를 이용하여 단편을 붙였다. PCR 을 이용하여 붙인 단편의 양을 증폭시켜 PCR 산물의 양을 증가시켰다. PCRquck-spinTM
product purification kit (Intron Biotechnology Inc., Seongnam, Gyeonggi-do, Korea) 을 이용하여 PCR 산물을 정제한 후, 제한효소 BamHⅠ과 XbaⅠ로 37℃에서 8 시간 동안 자른 후 0.8% agarose gel 에서 전기영동 하여 제한효소 처리한 DNA 만을 잘라서 QIAquick Gel extraction kit (Qiagen Co.,
Hilden, Germany
) 을 이용하여 회수하였다. pSP72-dper 또한 BamHⅠ과 XbaⅠ으로 자른 단편을 37℃에서 8 시간 동안 자른 후 0.8% agarose gel 에서 전기영동 하여 vector 단편을 QIAquick Gel extraction kit (Qiagen Co.,Hilden, Germany
) 을 이용하여 회수하였다. 회수한 pSP72 vector 단편과 dper mutagenesis 단편을 T4 DNA ligase (Takara bio Inc., Otsu, Shiga, Japan) 를 이용하여 단편을 붙인 뒤 JM109 (Promega Co., Madison, WI, U.S.A) 에 transformation 하여 colony 를 확인하였다. Colony 를 DNA-spin plasmid DNA purification kit (Intron Biotechnology Inc., Seongnam, Gyeonggi-do, Korea) 으로 mini prep 한 후 plasmid 일부를 BamHⅠ과 XbaⅠ으로 잘라서 원하는 DNA 라는 것이 확인되면 KpnⅠ과 XbaⅠ, ScaⅠ으로 자른 다음 전기영동한 후 QIAquick Gel extraction kit (Qiagen Co.,Hilden, Germany
)으로 dper 단편을 획득하였다. 또한 pAct-dper-V5 를 KpnⅠ과 XbaⅠ으로 잘라서 동일한 방법으로 pAc5/V5-His vector 단편을 획득한 후 dper 단편과 T4 DNA ligase (Takara bio Inc., Otsu, Shiga, Japan) 를 이용하여 단편을 붙인 뒤 JM109 (Promega Co., Madison, WI, U.S.A) 에 transformation 하여 colony 를 확인하였다. Colony 를 DNA-spin plasmid DNA purification kit (Intron Biotechnology Inc., Seongnam, Gyeonggi-do, Korea) 으로 mini prep 한 후 plasmid 일부를 BamHⅠ과 XbaⅠ으로 잘라서 원하는 DNA 라는 것이 확인되면 QIAGEN plasmid midi kit (Qiagen Co.,Hilden, Germany
) 으로 Midi prep 을 하여 plasmid DNA 를 다량 확보하였다.4. PCR-based Rapid site directed mutagenesis
를 위한
Primer 들
(A) dper951.952A_BsmBⅠ(F)
5‟TAG ATG CGT CTC T GCC GCG CAC ACG GCC ATG GCC CAG-3‟ (B) dper951.952A_BsmBⅠ(R)
5‟- TAG ATG CGT CTC TC GGC GTG AAC GGG CGG TGT GAT-3‟ (C) dper954A_BsmBⅠ(F)
5‟- TAG ATG CGT CTC T GCG GCC ATG GCC CAG AGC AGC-3‟ (D) dper954A_BsmBⅠ(R)
5‟- TAG ATG CGT CTC TC CGC GTG CGT GGA GTG AAC GGG-3‟ (E) dper959,960A_BsmBⅠ(F)
5‟- TAG ATG CGT CTC T GCC GCC TTC TCC TCC GCC GGC C-3‟ (F) dper959,960A_BsmBⅠ(R)
5‟- TAG ATG CGT CTC TC GGC CTG GGC CAT GGC CGT GTG-3‟ (G) dper962,963A_BsmBⅠ(F)
5‟- TAG ATG CGT CTC T GCC GCC GCC GGC CTC TTC CCG ACC-3‟ (H) dper962,963A_BsmBⅠ(R)
5‟- TAG ATG CGT CTC TC GGC GAA GCT GCT CTG GGC CAT-3‟ (I) dper976A_BsmBⅠ(F)
5‟-TAG ATG CGT CTC T GCC TTG ACG CCC ACC AGT CCC-3‟ (J) dper976A_BsmBⅠ(R)
5‟-TAG ATG CGT CTC TA GGC GGC GGG GAT GTA GTA GAA-3‟ (K) dper978A_BsmBⅠ(F)
5‟-TAG ATG CGT CTC T GCG CCC ACC AGT CCA CAG CGC-3‟ (L) dper978A_BsmBⅠ(R)
5‟-TAG ATG CGT CTC TG CGC CAA GGA GGC GGG GAT GTA-3‟ (M) dper951.952,954A_BsmBⅠ(R)
5‟-TAG ATG CGT CTC TC CGC GTG CGC GGC GTG AAC GGG-3‟ (N) dper954,959,960A_BsmBⅠ(F)
5‟-TAG ATG CGT CTC T GCG GCC ATG GCC CAG GCC GCC-3‟ (O) dper959,960,962A_BsmBⅠ(R)
5‟-TAG ATG CGT CTC TC GGC GAA GGC GGC CTG GGC CAT-3‟ (P) dper962,963,969A_BsmBⅠ(F)
5‟-TAG ATG CGT CTC T GCC GCC GCC GGC CTC TTC CCG GCC-3‟ (Q) dper981A_BsmBⅠ(F)`
5‟-TAG ATG CGT CTC T GCT CCC ACG CGC TCT CCC CGG-3‟ (R) dper981A_BsmBⅠ(R)
5‟-TAG ATG CGT CTC TG AGC GGT GGG CGT CAA GGA GGC-3‟ (S) dper976,978,981A_BsmBⅠ(F)
5‟-TAG ATG CGT CTC T GCC TTG GCG CCC ACC GCT CCC-3‟ (T) dper976,978A_BsmBⅠ(F)
5‟-TAG ATG CGT CTC T GCC TTG GCG CCC ACC AGT CCC-3‟ (U) dper976,978,981A_BsmBⅠ(F)
5‟-TAG ATG CGT CTC T GCC TTG GCG CCC ACC GCT CCC-3‟ (V) dper_BamHⅠ(F)
5‟-ATG GAT CCG GAA CAG GCG TAG CAG-3‟ (W) dper_XbaⅠ(R)
C. Drosophila melanogaster Schneider line 2 (S2) 세포의 배양과
transfection
Drosophila melanogaster Schneider line 2
(
S2) 세포는 10% Fetal Bovine Serum, Certified (FBS) (Invitrogen Co., Carlsbad, CA, U.S.A) 와 0.5% penicillin streptomycine (Sigma-Aldrich Co., Louis, MO, U.S.A) 이 포함된 Schneider‟s Drosophila medium (Invitrogen Co., Carlsbad, CA, U.S.A) 를 사용하였고 23℃ 항온기에서 배양하였다. Transfection 은 Effectene Transfection reagent (Qiagen Co.,Hilden, Germany
) 를 사용하였고 제조사가 제시한 실험방법에 따라 실험하였다. 각 transfection 은 0.6 ㎍ 의 각각 다른 dper 를 포함한 plasmid 와 0.2 ㎍ 의 pMT-dbt-V5/His 또는 0.4 ㎍ 의 pMT-HAdclk 을 사용하였다. pMT-dbt-V5/His 와 pMT-HAdclk 은 transfection 후 최종 500 uM CuSO4 를 넣어 induction 함으로써 특정시간 동안D. 형질전환 초파리 (Transgenic Drosophila melanogaster)
우리가 새롭게 만든 형질전환 초파리들은 변이된 dper 를 가지고 있다. 즉 13.2per+(Δ926-977)-HA10His 를 발현하는 초파리를 만들었다. 우리는 13.2kb dper genomic 단편의 C-말단에 HA epitope 와 다수의 histidine 잔기 (10XHis) 가 표지 되어 있는 이전에 그 특성이 기술되어 있는 vector (13.2per+
-HAHis) 를 이용하였다 (Lee 등, 1998). 이 plasmid 의 BamHⅠ-KpnⅠ단편을 pSP72 vector (Promega Co., Madison, WI, U.S.A) 속에 subclone 하였다. 이것은 QuickChange Mutagenesis Kit (Stratagene Co., La Jolla, CA, U.S.A) 를 사용하여 site-direct mutagenesis 를 수행하기 위한 주형으로 사용하였다. 변이된 dper 영역은 DNA sequencing analysis 를 통해 확인하였고, 상응하는 단편을 13.2per+-HAHis plasmid 의 제자리에 되돌려놓았다. 이렇게 만든 plasmid 를 13.2per+
(Δ926-977)-HA10His 라 명명한다. 이 plasmid 를 초파리 배아에 주입하여 형질전환 초파리를 제작하였다. w1118 배아에 plasmid 를 주입하는 것은 BestGene Inc (BestGene Inc., Chino Hills, CA, U.S.A.)에 의뢰하여 수행하였다. w1118per+ (wper+이라 함) 와 w1118
per0 (wper0 이라 함) 의 배경에 야생형 또는 변이된 dper 이식유전자를 발현하는 형질전환 초파리는 이미 논문에서 기술하고 있다 (Kim 등, 1997) .
E. Locomotor activity assays
초파리 각각의 locomotor activity 는 Drosophila Activity System (DAM) 을 이용하여 측정하였다 (Waltham MA) (Hemblen-Coyle 등, 1992). 측정을 위해 초파리는 어린 수컷 성체를 사용하였다. 25℃ 항온기에서 12 시간간격으로 빛과 어둠 속에 총 4 일간 노출하였고 (12:12LD, zeitgeber time zero [ZT0] 는 light phase 가 시작되는 시점으로 정의를 내린다.), Constant-darkness condition (DD)을 12:12LD 이후 연속해서 7 일간 유지하였다. 각 초파리의 Locomotor activity 는 Dr. F. Rouyer. (France)로부터 받은 FaasX software (MacOSX 를 위한 Fly Activity Analysis Suite) 를 이용하여 분석하였다. Period 는 초파리 각각을 chi-square periodogram analysis 하여, 각 유전형의 그룹별 평균을 산출하여 그 값을 획득하였다. Power 는 DD 동안 rhythm 의 strength 의 정도이다. 각각의 초파리들은 power 가 10 이상의 값을 가지고 width 값 (periodogram 95% confidence line 보다 30 분 위에 peak 된 수를 표시) 이 2 이상의 rhythm 을 나타낸다.
F. Immunoblot analysis
전기영동용 Gel 은 6% polyacrylamide gel (Resolving gel mix; 1 ㎖ 의 29.6% polyacrylamide + 0.4% Bis, 1.94 ㎖ 의 DW, 2 ㎖ 의 1 M Tris-Hcl pH 8.8, 50 ㎕ 의 10% SDS, 7.5 ㎕ 의 25% APS, 5 ㎕ 의 TEMED) (Stacking gel ; 375 ㎕ 의 29.2% Achrylamide + 0.8% Bis, 1.85 ㎖ DW, 250 ㎕ 의 1 M Tris-Hcl pH 6.8, 25 ㎕ 의 10% SDS, 7.5 ㎕ 의 25% APS, 5 ㎕ 의 TEMED)을 제작하여 사용했다. 시료의 전 처리를 위해 S2 세포를 거두어들인 뒤 PBS 로 한 번 씻어준 후 150 ㎕ 의 Modified RIPA (50 mM Tris-Hcl pH 7.5, 1% NP-40, 0.25% deoxycholate, 50mM NaCl, 5% glycerol,) 에 신선한 1 mM EDTA, compete EDTA-free protease inhibitor cocktail (Roche Diagnostics Co., Mannheim, Germany), PhosSTOP phosphatase inhibitor cocktail (Roche Diagnostics Co., Mannheim, Germany) 을 추가하여 세포를 용해하였다. 용해방법은 얼음 위에서 5 분마다 10 초씩 세 번 vortexing 하여 총 15 분간 용해한 후 4℃에서 13000 rpm 으로 15 분간 원심 분리하여 상층액만을 회수하였다. 회수한 각 세포용해액은 ELIZA leader (BioTex Inc., Houston, TX, U.S.A) 를 이용하여 595 nm 로 농도를 측정하여 OD 3 으로 정량한 후 단백질을 변성시키기 위해 NuPAGE LDS sample buffer (Invitrogen Co., Carlsbad, CA, U.S.A) 와 50 mM DTT 을 추가하여 95℃ 이상의 온도에서 5 분간 끓인 후 spindown 하여 sample 들을 loading 하였다. 준비된 sample 들은 SDS-PAGE 전기영동법 (Sodium Dodecyl Sulfate – Polyacrylamide gel Electrophoresis, Laemmli 법) 을 수행하였다. Mini-PROTEAN Tetra Cell (BIO-RAD Laboratories, Inc., Hercures, CA, U.S.A) 을 이용하였고, 전기영동용 buffer (1 L ; 3.025 g Tris, 14.4 g Glycine, DW, 1% SDS) 를 채운 뒤 전기영동 하였다. Stacking gel 에서는 130 V 로 Resolving gel 에서는 190 V 로 설정
하였고 PRO-STAIN Prestained protein marker (Intron Biotechnology Inc., Seongnam, Gyeonggi-do, Korea) 을 함께 loading 하여 marker 의 115 kDa 밴드가 gel 의 가장 하단에 위치할 때까지 전기영동 하였다. 전기영동 완료된 gel 은 resolving gel 만을 잘라낸 후 transfer buffer 에 충분히 적신 Gel Blot Paper (Whatman Inc., Springfield Mill, Kent, U.K) 와 PROTRAN BA 85 NITROCELLULOSE (Whatman GmbH Inc., Hahnestr, Dassel, Germany) 를 함께 semi-dry blot sandwich 를 만들어 Trans-Blot SD semi-dry electrophoretic transfer cell (BIO-RAD Laboratories Inc., Hercures, CA, U.S.A) 에 함께 장착한 다음 240 ㎃ 에서 45 분간 Transfer 하였고, Transfer 된 nitrocellulose transfer membrane 은 0.1% Ponseau S (0.1% Ponceau S, 5% acetic acid) 로 확인하였다. Orbital shaker 위에서 5% Blocking 용액 (5% 탈지분유, TBST, 0.05% Thimerosol) 에 넣어 상온에서 30 분간 blocking 하였고, 1 차 항체가 포함된 5% Blocking 용액에 넣어 냉장실에서 하루 동안 반응시킨 뒤, 2 차 항체가 포함된 5% Blocking 용액에 넣어 상온에서 두 시간 동안 반응하였다. 이 때 Anti-V5 (Invitrogen Co., Carlsbad, CA, U.S.A) 는 1:20000, Anti-HA (3F10) (Roche Diagnostics Co., Mannheim, Germany) 은 1:4000 으로 희석하여 사용하였고, Goat anti-mouse IgG (H+L), horseradish peroxidase (Molecular Probes Inc., Eugene, OR, U.S.A) 는 1:10000, Goat anti-rat IgG (H+L), horseradish peroxidase (Molecular Probes Inc., Eugene, OR, U.S.A) 는 1:5000 으로 희석하여 사용하였다. 항체 반응이 끝난 뒤에는 항상 15 분 간격으로 4 번 TBST (4 M NaCl, 1 M Tris-HCl pH 7.5, 0.05% Tween-20, DW) 로 세척하였다. 항체반응이 끝나면 Immobilon Western chemiluminescent HRP substrate (Millipore Co., Billerica, MA, U.S.A)를 nitrocellulose transfer membrane 에 적절히 묻힌
후 CP-BU NEW medical X-ray film blue (Agfa HealthyCare Co., Soptestraat, Mortsel, Belgium) 에 적정시간 동안 노출시켜 현상하였다.
G. Immunoprecipitation (IP)
pMT-HAdclk 0.4 ㎍ 과 함께 pAct-dper-V5/His 또는 pAct-dper(Δ)-V5/His series 0.6 ㎍ 을 S2 세포에 Effectene Transfection reagent (Qiagen Co.,
Hilden, Germany
) 을 사용하여 제조사의 설명에 따라 transient transfection 하였다. 36 시간 동안 배양한 뒤, 최종 500 uM CuSO4 를 넣어 dCLK 의 발현을 유도한 후 24 시간 동안배양하였다. 세포를 수확하고 PBS 로 한 번 씻어준 후 Modified RIPA (50 mM Tris-Hcl pH 7.5, 1% NP-40, 0.25% deoxycholate, 50 mM NaCl, 5% glycerol,) 에 신선한 0.5 mM PMSF, 2.5 mM NaF, 1 mM Na2VO3, compete EDTA-free protease inhibitor cocktail (Roche Diagnostics Co., Mannheim, Germany) 를 추가한 lysis buffer 를 이용하여 세포를 용해하였다. 용해 방법은 얼음 위에서 5 분마다 10 초씩 세 번 vortexing 하여 총 15 분간 용해한 후 13000 rpm 으로 15 분간 4℃ 원심 분리하여 상층액만을 회수하였다. 회수한 세포용해액은 ELIZA leader (BioTex Inc., Houston, TX, U.S.A) 를 이용하여 59 5nm 에서 O.D 값을 측정한 후 단백질 양을 동일하게 맞추고 각각 dPER 에 대한 Anti-V5 항체 (Invitrogen Co., Carlsbad, CA, U.S.A) 또는 dCLK 에 대한 Anti-HA (12CA5) 항체 (Roche Diagnostics Co., Mannheim, Germany) 를 2 ㎕ 씩 넣어준 다음 4℃ 에서 세 시간 동안 lotation 하였다. 그 동안 GammaBind G Sepharose (GE Healthcare Bio-Sciences AB., Björkgatan, Uppsala, Sweden) bead 를 사용할 만큼 미리 준비하여 세포용해시 사용했던 동일한 buffer (Modified-RIPA) 로 약하게 세 번 세척하였다. 세 시간이 경과하면 각 sample 마다 준비한 bead 를 25 ㎕ 씩 넣어준 후 다시 4℃ 에서 한 시간 동안 lotation 하였다. Sample 들은 4℃ 에서 2 분 동안 원심 분리한 후 상층액은 따로 모아두고, bead 를 다시 4℃ 에서 5 분간 lotation 하여 buffer (Modified-RIPA) 로 세척하는 과정을 세
번 반복한 후 buffer (Modified-RIPA) 를 최대한 제거한다. 1.2 X Sample buffer 를 각 sample 마다 30 ㎕ 씩 넣고 95℃ 에서 5 분간 끓인 후 15 ㎕ 씩 loading 하여 immunoblotting 을 수행하였다. Immunoblotting 에는 6% polyacrylamide gel 을 사용하였고, IP 수행 시 Anti-V5 (Invitrogen Co., Carlsbad, CA, U.S.A) 로 dPER 를 검출한 sample 에는 dCLK 을 찾기 위해 1 차 항체로 Anti-HA (3F10) (Roche Diagnostics Co., Mannheim, Germany) 을 사용했고 Anti-HA (12A5) (Roche Diagnostics Co., Mannheim, Germany) 로 dCLK 을 검출한 sample 에는 dPER 를 찾기 위해 Anti-V5 (Invitrogen Co., Carlsbad, CA, U.S.A) 항체를 사용하였다. 또한 원래의 세포용해액은 dCLK 은 O.D 6, dPER 는 O.D 2 로 동량하여 함께 loading 하여 줌으로써 세포 내 존재하는 각 단백질 양과 dPER 또는 dCLK 의 각각의 결합도를 비교하였다.
H. Luciferase Reporter Assay 와 β-Galactosidase Enzyme Assay
System
Assay 를 위해서 먼저 세포용해액을 준비해야 한다. 24 well plate 에 S2 세포를 배양한 후 dperE-Luc, pAct5.1-LacZ-V5/His, pMTB-HAdclk-V5/His 와
pAct-dper-V5/His 또는 pAct-dper(mutant)-V5/His series plasmid 를 Effectene Transfection
reagent (Qiagen Co.,
Hilden, Germany
) 을 사용하여 제조사의 설명에 따라서 transfection 하였고, 모든 sample 의 transfection 효율을 동일화 시키기 위해 pMTB 로 보정하였으며 오차범위를 줄이기 위해 triplecation 하였다. 36 시간 후 최종 500 mM CuSO4 로 induction 한 다음 12 시간 후 well 을 PBS 로 두 번 세척하였다. 5X Reporter Lysis Buffer (RLB) (Promega Co., Madison, WI, U.S.A) 를 1 X 로 희석하여 각 well 의 표면을 충분히 덮을 수 있을 정도로 150 ㎕ 를 분주하였고, 4℃ shaker 로 강하게 흔들면서 20 분 가량 incubation 하면서 세포를 최대한 용해시켰다. 세포용해액은 4℃ 에서 13000 rpm 으로 5 분간 원심분리하여 상층액만을 회수하였다. Luciferase reporter assay 를 위해서 상온에서 Luciferase Assay Reagent (LAR) (Promega Co., Madison, WI, U.S.A) 를 녹인 다음 1.5 ㎖ microcentrifuge tube 에 100 ㎕ 의 LAR 와 세포용해액을 넣어 잘 섞어서 delay 는 3 초, integration time 은 15 초, replication 은 1 로 설정된 Luminometer (Turner Designs Inc., Sunnyvale, CA, U.S.A) 에서 측정하였다. 이 때 세포용해액 중 dper 가 없는 dClk 을 2 ~ 10 ㎕ 을 넣어 측정하였을 경우 측정값이 약 3000 이 나올 때 적절하다.β-Galactosidase Enzyme Assay 를 위해서 96 well plate 에 각 sample 20 ㎕, 1X RLB 30 ㎕ 와 Assay 2 X Buffer (2 mM MgCl2, 100 mM 2-mercaptoethanol, 1.33 mg/㎖
ONPG) 50 ㎕ 를 넣고 37℃ 에서 노란색으로 변할 때까지 약 30 분 정도 반응시킨 후 150 ㎕ 의 1 M Sodium Carbornate 를 넣어 반응을 중단시키고 즉시 ELIZA leader (BioTex Inc., Houston, TX, U.S.A) 를 이용해 420 nm 의 파장으로 측정하였다. β-Galactosidase Enzyme Assay 값을 이용하여 Luciferase Reporter Assay 값을 보정하여 graph 를 작성하였다.
Ⅲ. 결 과
A. S2 세포에서 dPER(Δ926-977) 은 dCLK
의 전사활성을
억제하지 못한다.
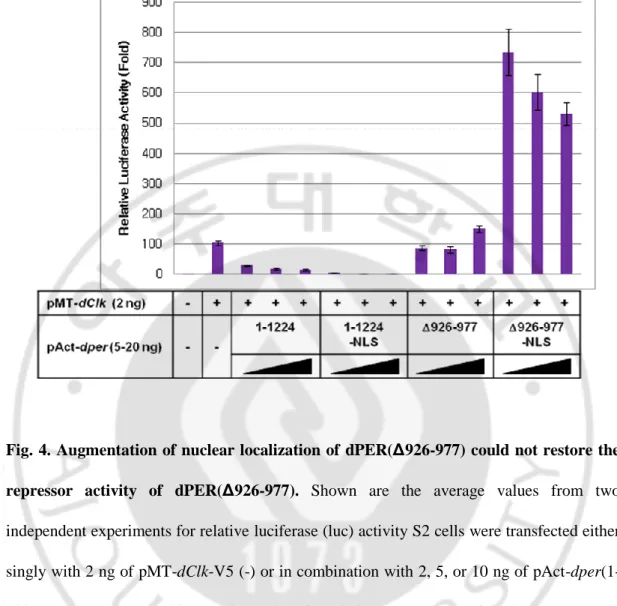
최근 우리 연구실에서는, dPER 의 단백질에서 전사 억제제로서의 활성에 중요한 부위인 CCID (dCLK:CYC Inhibition Domain) 를 luciferase-reporter assay 를 통해 앞서 연구된 dPER CCID 에 존재하는 아미노산 926-977 을 제거하게 되면 전사 억제제로서의 활성을 잃어버리게 된다는 연구결과를 얻게 되었다 (Data not shown). dPER(Δ926-977) 단백질이 전사 억제제로서의 활성을 잃는 것이 이 단백질의 세포 내 분포가 변하였기 때문일 가능성을 먼저 조사해보았다. NLS (Nucleus Localization Signal) sequence 를 dper 와, dper(Δ926-977) 에 각각 붙여 핵 내로의 이동을 인위적으로 원활하게 한 후 이들의 전사 억제제로서의 활성을 luciferase-reporter assay 로 측정하였다 (Fig. 4). dPER 는 농도에 따라 dCLK 의 전사활성을 억제하는 반면 dPER(∆926-977) 은 농도가 증가하여도 dCLK 의 전사활성을 억제하지 못하는 것을 확인할 수 있었다. 이 때 dPER-NLS 가 동일한 농도의 dPER 에 비해 dCLK 의 전사활성을 더욱 억제하는 것으로 보아 NLS sequence 의 첨가로 dPER 의 핵 내로의 이동이 촉진되어 전사 억제제로서의 활성이 증가 하였슴을 확인할 수 있었다. 반면 dper(∆926-977) 의 경우에는 NLS sequence 를 첨가 하였슴에도 dCLK 의 전사가 여전히 억제되지 않음을 알 수 있었으며 오히려 dCLK 의 전사활성이 증가하는 현상을 관찰할 수 있었다. 따라서 dPER(∆926-977) 이 dCLK 의 전사활성을 억제하지 못하는 것은 적어도
dPER(∆926-977) 가 세포질에서 핵으로 이동하지 못하기 때문에 일어나는 것은 아니라는 것을 나타낸다. dPER(∆926-977) 의 양이 증가함에 따라 dCLK 의 전사활성이 오히려 증가하고 그 효과가 NLS sequence 를 첨가하였을 때 증폭되는 것으로 보아 아미노산 926-977 을 dPER 로부터 제거하였을 때 dominant negative 하게 작용하는 것으로 생각된다.
Fig. 4. Augmentation of nuclear localization of dPER(Δ926-977) could not restore the repressor activity of dPER(Δ926-977). Shown are the average values from two independent experiments for relative luciferase (luc) activity S2 cells were transfected either singly with 2 ng of pMT-dClk-V5 (-) or in combination with 2, 5, or 10 ng of pAct-dper(1-1224) or pAct-dper(1-pAct-dper(1-1224)-NLS or pAct-dper(Δ926-977) or pAct-dper(Δ926-977)-NLS. Cells were incubated with 500 uM CuSO4 (final in the media) to induce ectopic expression
of dCLK at 36 h after transfection and harvested 24 h later. Luc activity in the absence of transfecting pMT-dClk-V5 was set to 1, and all other values were normalized.
B. dPER(Δ926-977)
은 DBT
에 의한 인산화에 결함을
나타내었다.
이전의 연구 결과 dPER(Δ755-809) 는 DBT 와 결합하지 못하여 DBT 에 의한 인산화가 일어나지 못하며 또한 dCLK 의 전사활성을 억제하지 못하였다. (Kim 등, 2007). 이에 Kim 등은 아미노산 755-809 를 dPER DBT binding domain (dPDBD)라 명명하였다. 아미노산 926-977 이 dPDBD 와 매우 인접하여 있으므로 dPER(Δ926-977) 이 전사 억제제로서의 활성을 잃은 것이 DBT 와의 결합에 문제가 생겼기 때문인지를 조사하기 위하여 먼저 DBT 에 의한 dPER(Δ926-977) 의 인산화 정도를 조사하여 보았다. 그래서 각각의 pAct-dper(1-1224)-V5, 또는 V5, 또는 pAct-dper(Δ755-809)-V5, 또는 dper(Δ768-842)-V5, 또는 dper(Δ843-925)-dper(Δ768-842)-V5, 또는 dper(Δ926-977)-dper(Δ768-842)-V5, 또는
pAct-dper(Δ978-999)-V5 와 pMT-dbt-V5 를 cotransfection 후 시간에 따라 dPER 가
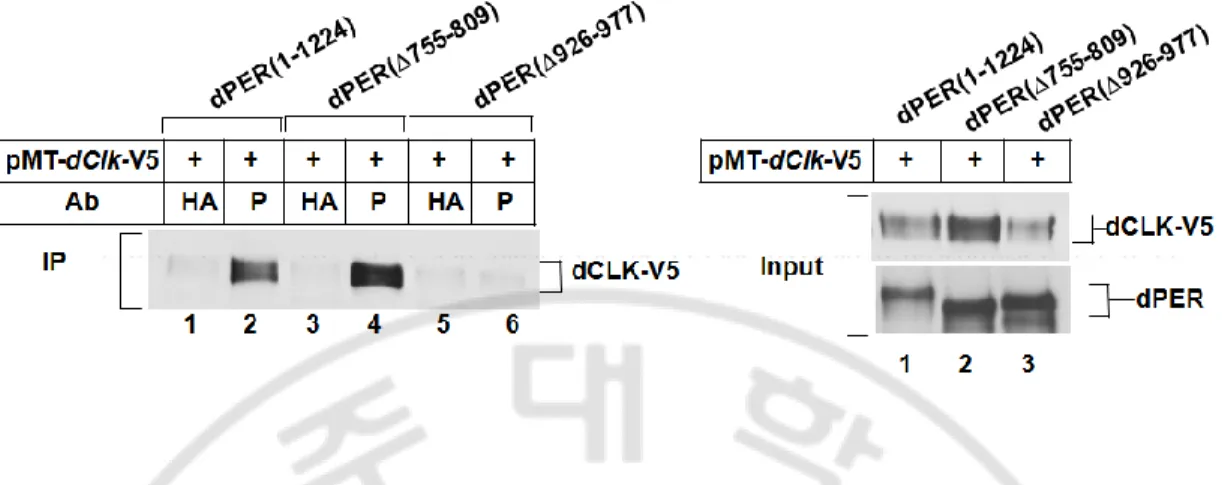
인산화되는 정도를 비교하여 보았다. 그 결과 dPER(Δ843-925) 와 dPER(Δ978-999) 는 DBT 를 induction 후 비교하였을 때 wild-type 의 dPER 와 유사한 정도로 시간에 따라 hyperphosphorylation 되고 분해됨을 확인할 수 있었다. 그러나 dPER(Δ926-977) 은 dPDBD 를 포함하는 아미노산 부위가 결핍된 dPER(Δ768-842) 와 유사하게 DBT 에 의한 hyperphosphorylation 과 이에 따른 단백질의 분해가 더디게 일어나는 것을 확인하였다 (Fig. 5A). 이것은 dPER(Δ926-977) 은 DBT 에 의한 인산화에 심각한 결함을 가지는 것을 알 수 있었다. 그렇다면 이러한 결함이 DBT 와의 interaction 의 결함 때문은 아닌가를 조사하기 위하여 immunoprecipitation assay 를 수행하였다. 이때 CCID 내에 초파리 종간 보존성을
기준으로 내부적으로 제거한 네 가지의 dPER 변이체를 함께 조사하였다 (Fig. 5B). dPDBD 가 결핍된 dPER(Δ755-809) 과 dPDBD 를 포함하며 CCID 내에 종간 보존성이 높은 아미노산 768-842 를 제거한 dPER(Δ768-842) 는 wild-type 의 dPER 에 비해 DBT 와의 결합이 매우 감소되어 있는 것을 확인할 수 있었다. 반면에 CCID 에서 초파리 내 종간 보존성이 떨어지는 부위인 아미노산 843-925 가 결핍된 dPER 는 DBT 와의 결합이 오히려 증가하였으며 아미노산 978-999 가 결핍된 dPER 는 wild-type 과 유사한 정도로 DBT 와 결합하는 것이 확인되었다. wild-type 의 dPER 에 비해 오히려 dPER(Δ926-977) 은 dPER(Δ843-925) 와 유사한 정도로 DBT 와의 결합이 증가하는 양상을 나타내었다. 흥미롭게도 IP 에 사용한 input lysate 를 비교해 본 결과 dPER wild-type 과 dPER(Δ843-925), dPER(Δ978-999) 는 dPER 의 leve0l 이 감소한 것을 관찰할 수 있는 반면 dPDBD 가 결핍된 dPER(Δ755-809) 과 dPER(Δ768-842), 그리고 dPER(Δ926-977) 은 단백질의 양이 pMT-dbt-V5 를 transfection 하지 않은 조건과 유사한 수준으로 유지되는 것을 관찰할 수 있었다. dPER(Δ926-977) 은 DBT 와의 결합이 정상적임에도 불구하고 DBT 에 의한 인산화에 심각한 결함을 나타내었다.
Fig. 5. DBT dependent phosphorylation of dPER(Δ926-977) is attenuated in S2 cells. S2 cells were transfected with 600 ng of nontagged (B) versions of dper-containing plasmids (as indicated on bottom of panel). (A and B) There presense (+) or absence (-) of 200 ng of pMT-dbt-V5 is indicated. Exogenous DBT was induced 36 h after transfection by adding to the media 500 uM CuSO4 (final). Cells were harvested 12 h (B) later. (B) Extracts were
prepared and either subjected to immunoprecipitaion (IP) or analyzed directly (input). Immune complexes were recovered with anti-V5 (added specific antibody) antibody. Immunoblots were probed with antibodies against anti-dPER(GP339) (to detect dPER), or anti-V5 (to detect DBT), as indicated. (A) Cells were harvested at the indicated times, and extracts were analyzed by immunoblotting in the presence of anti-V5 antibodies.
C. S2 cells 에서 dPER(Δ926-977) 은 dCLK 과 결합하지 못한다.
dPER 단백질은 핵 내에서 dCLK 의 전사활성을 억제할 때에 dCLK 단백질과 complex 를 이루는 것이 잘 알려져 있다 (Lee 등, 1998; Lee 등, 1999). dPER(Δ977) 이 dCLK 의 전사활성을 억제하지 못하는 이유가 아미노산 926-977 의 제거에 의해 dCLK 과 결합하지 못하기 때문은 아닌지 조사해보기 위해 dCLK 과 dPER 또는 dPER(Δ926-977) 와 immunoprecipitation assay 를 수행하였다 (Fig. 6). dPER 와 dPDBD (Drosophila PER DBT Binding Domain) (아미노산 755-809) 가 결핍되어 dCLK 의 전사활성을 억제하지 못하는 dPER(Δ755-809) 는 dCLK 과 함께 immunoprecipitation 이 되는 반면에 (lane 2 and 4), dPER(Δ926-977) 은 전혀 immunoprecipitation 이 되지 않는 것을 알 수 있다 (lane 6). 이러한 결과는 아미노산 926-977 이 dPER 에서 dCLK binding domain 으로 작용했을 것이라는 사실을 나타낸다.
Fig. 6. dPER(Δ926-977) could not directly bind to dCLK in S2 cells. S2 cells were transfected with 600 ng of pAct-dper(1-1224) or pAct-dper(Δ755-809) or pAct-dper (Δ926-977) in combination with 400 ng of pMT-dClk-V5. At 36 h after transfection, cells were incubated in media containing 500 uM CuSO4 (final) to induce ectopic expression of target
proteins and harvested 24 h later. Extracts were prepared and either subjected to immunoprecipitaion (IP) or analyzed directly (input). Immune complexes were recovered with anti-dPER (GP339) (added specific antibody) or anti-HA (12CA5) (added nonspecific antibody) antibodies. Immunoblots were probed with antibodies against anti-V5 (to detect dCLK), or anti-dPER (GP339) (to detect dPER), as indicated.
D.
dPER(Δ926-977) 을 발현하는 형질전환 초파리는 주기가
길어지기는 하였으나 정상적인 생체리듬을 가졌다.
dPER 의 아미노산 926-977 의 in vivo 에서의 역할을 탐색하기 위하여 아미노산 926-977 부위가 제거된 dPER 를 발현하는 초파리를 제작하였다. 13.2Kb 의 genomic dper[13.2(dper+)]와 HA epitope tag 그리고 10His 를 포함하는 plasmid 에서 아미노산 926-977 을 제거하여 microinjection 에 사용하였다. dPER(Δ926-977)을 발현하는 독립적인 두 개의 형질전환 초파리 라인을 확립하였고 dper0 background 의 초파리와 교배 후 이를 p{dper(926-977)} 초파리라 명명하였다. 정상적인 dper 를 발현하는 초파리를 역시 dper0 background 의 초파리와 교배 후 이를 p{dper(WT)} 라 명명하고 대조군으로 사용하였다. 생체리듬은 DAM (Drosophila Activity Monitoring) system 을 이용하여 분석하였다 (Table. 1). 그 결과 p{dper(WT)} 초파리는 약 23.2 시간의 주기를 가지고 rhythmic 한 반면 p{dper(926-977)} 초파리는 약 26.7 시간의 주기를 가지며 rhythmic 한 것을 알 수 있었다 (Table. 1). 온도가 변하더라도 그 주기가 일정하게 유지되는 것이 “temperature compensation” 이라고 잘 알려진 생체리듬의 독특한 특성이다. p{dper(WT)} 의 생체리듬은 온도가 18℃ 로 낮아지거나 29℃ 로 올라가더라도 일정하게 약 24 시간의 주기를 유지하는 반면에 p{dper( 926-977)} 의 생체리듬은 온도가 올라감에 따라 길어지는 것을 확인할 수 있었다 (Table. 1). 이는 dPER(Δ926-977) 단백질의 생화학적 결함이 온도가 올라가면서 심해진다는 것을 말해주고 있다. 그렇다면 생체리듬 주기가 길어진 p{dper( 926-977)} 형질전환 초파리의 생체 내에서 dPER 는 어떠한 변화가 있는지 조사하기
위해 p{dper(926-977)} 형질전환 초파리를 12:12LD cycle 에 entrainment 시킨 후 4 시간 간격으로 초파리를 수확하여, 머리로부터 단백질 액을 얻어 western blot 을 수행 하였다 (Fig. 7). 그 결과 p{dper(926-977)} 형질전환 초파리는 S2 세포에서 DBT 가 있음에도 불구하고 매우 더디게 degradation 되던 in vitro 결과와 유사한 양상이 관찰되었다. 즉 dper+를 발현하는 dper0 ;;dper-wt,M16 초파리의 경우 ZT20, ZT23.8 에 인산화 결과 dPER 의 mobility 가 느려지는 것은 볼 수 있으나 dPER(Δ926-977) 을 발현하는 형질전환 초파리 라인에서는 mobility 가 느린 isoform 이 관찰되지 않는 것을 알 수 있다. 또한 ZT20 에서 dper-wt 과 dperΔ926-977 의 밴드 intensity 를 비교해 보면 (lane 6 와 8) dperΔ926-dperΔ926-977 의 level 이 약 1.5-2 배 정도 더 높은 것을 알 수 있다. p{dper(926-977)} 초파리에서 TIM 단백질의 molecular rhythm 의 변화도 dPER 의 그것과 유사하였다. 즉 p{dper(926-977)} 초파리에서 TIM 단백질의 양은 약 1.5 ~ 2 배 정도 높았고 peak time 이 p{dper(WT)} 초파리에 비해서 늦었다.
TABLE. 1. Locomotor activity rhythms of p{dper(926-977)} flies and control
p{dper(WT)} transgenic fliesa
Genotypeb Temp (C) Period±SEMc Power±SEMd % Rhythmice Num Totalf
w1118 18 23±0.75 41.8 50 32 wper01;; p{dper(WT)}(M16) 18 23.4±0.42 34.6 29 31 wper01; p{dper(926-977)}(M1) 18 24.3±0.16 36.8 27.6 29 wper01;; p{dper(926-977)}(F3) 18 24.1±0.17 60.6 93.5 31 perS 18 19.2±0.11 49.3 38.5 26 perL 18 27.5±0.38 33.9 21.9 32 w1118 25 23.3±0.08 74.9 73.3 16 wper01;; p{dper(WT)}(M16) 25 23.3±0.05 141.7 100 32 wper01; p{dper(926-977)}(M1) 25 26.2±0.22 69.4 68.8 16 wper01;; p{dper(926-977)}(F3) 25 26.2±0.15 100.1 93.1 31 perS 25 19.7±0.81 108 74.2 32 perL 25 28.1±0.11 116.8 89.7 32 wper01;; p{dper(WT)}(M16) 29 23.3±0.05 106.8 100 32 wper01; p{dper(926-977)}(M1) 29 28±0.17 61.3 66.7 30 wper01;; p{dper(926-977)}(F3) 29 27.5±0.07 122.4 100 32 perS 29 18.7±0.05 74 70 30 perL 29 29.9±0.08 98.9 96.9 32 a
Flies were kept they indicated temperature and exposed to 4 days of 12:12LD followed by 7 days of DD.
b
Independent transgenic lines are designated by the numbers in parenthesis.
c
Period is Indicate period of circadian rhythm.
d
Power is a measure of the strength or amplitude of the rhythm in arbitrary units.
e
Percentage of flies showing locomotor rhythms with power value of ≥10 and a width value of ≥2.
f
Fig. 7. Molecular rhythm of dPER and TIM in p{dper(926-977)} flies. Adult flies of each genotype (indicated above the panel; WT, wper01;; p{dper(WT)}(M16); 926-977,
wper01;; p{dper(926-977)}(F3)) were collected at the indicated ZT or CT. Head extracts were prepared and analyzed by immunoblotting. Anti-HA (3F10) or anti-TIM (TR-3) antibodies were used to visualize dPER or TIM, respectively.
E. 아미노산 949-977
이 dCLK 과의 결합에 필요한 최소
도메인이다.
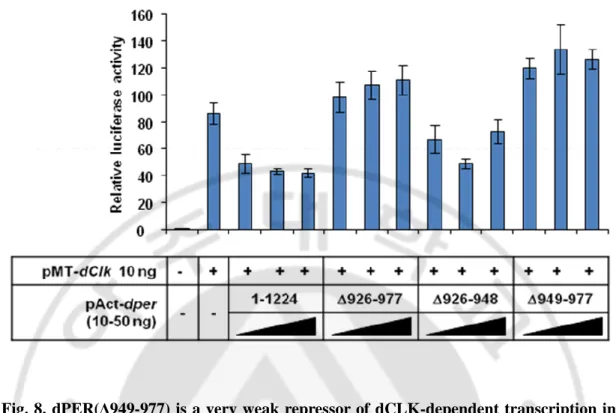
아미노산 926-977 부위에서 기능적으로 중요한 부위를 더욱 좁혀보기 위하여 927-977 부위를 두 개의 부위로 나누어 각각 제거한 dPER 변이체를 제작하였다. 아미노산 926-948 이 제거된 dPER(Δ926-948) 과 아미노산 949-977 이 제거된 dPER(Δ949-977) 를 제작하였고, 이들이 dCLK 의 전사 억제제로서의 활성이 dPER(Δ926-977)과 동일한 양상을 보이는지 조사하기 위해 앞서 이용하였던 동일한 방법으로 luciferase-reporter assay 를 수행하였다 (Fig. 8). 그 결과 dPER(Δ926-948) 은 dPER 와 유사하게 정상적으로 dCLK 의 전사를 억제하며, dPER(Δ949-977) 은 dPERΔ926-977 과 유사하게 dCLK 의 전사를 억제하지 못한다. 또한 dPER(Δ926-948) 과 dPER(Δ949-977) 의 DBT 에 의한 인산화 양상을 조사하였다 (Fig. 9). dPER 의 인산화 정도를 살펴본 결과 dPER(Δ926-948) 은 dPER 처럼 시간이 지남에 따라 정상적으로 인산화가 점차 많이 되는 것을 볼 수 있지만 dPER(Δ949-977) 은 인산화가 매우 느려지는 dPER(Δ926-977) 과 동일한 인산화 양상을 나타내었다. 그래서 dPER(Δ949-977) 이 dPER(Δ926-977) 처럼 dCLK 의 전사 억제제로서의 활성을 가지지 못하고 DBT 에 의한 인산화에서 결함을 나타낸다면 dCLK 과 interaction 에는 이상이 없는지 조사하기 위해 immunoprecipitation 을 수행 하였다 (Fig. 10). 그 결과 dPER(Δ926-948) 은 dPER 와 유사하게 정상적으로 dCLK 과 결합하며, dPER(Δ949-977) 은 dPER(Δ926-977) 과 유사하게 dCLK 과 결합하지 못한다. 이상의 결과들로부터 dPER 의 아미노산 926-977 도메인 중 아미노산 949-977 부위가 전사억제제로서의 활성과 dCLK 과의 interaction, DBT 에 의한 인산화 기능을 나타내기 위해 필수적인 부위임을 알 수 있다.
Fig. 8. dPER(Δ949-977) is a very weak repressor of dCLK-dependent transcription in S2 cells. Shown are the average values from two independent experiments for relative luc activity. S2 cells were transfected either singly with 10 ng of pMT-dClk-V5 (-) or in combination with 10, 20, or 50 ng of dper(1-1224) or dper(Δ926-977) or
pAct-dper(Δ926-948) or pAct-dper(Δ949-977). Cells were incubated with 500 uM CuSO4 (final
in the media) to induce ectopic expression of dCLK at 36 h after transfection and harvested 24 h later. Luc activity in the absence of transfecting pMT-dClk-V5 was set to 1, and all other valus were normalized.
Fig. 9. Deletion of aa 949 to 977 from dPER strongly attenuates DBT-dependent hyperphosphorylation of dPER in S2 cells. S2 cells were transfected with 600 ng of V5-tagged versions of dper-containing plasmids. The presence (+) of 200 ng of pMT-dbt-V5 is indicated. Exogenous DBT was induced 36 h after transfection by adding to the media 500
M CuSO4. Cells were harvested at the indicated times, and extracts were analyzed by
Fig. 10. dPER(Δ949-977) could not directly bind to dCLK in S2 cells. S2 cells were transfected with 600 ng of pAct-dper(1-1224) or pAct-dper (Δ926-977) or pAct-dper(Δ926-948) or pAct-dper(Δ949-977) in combination with 400 ng of pMT-dClk-V5. At 36 h after transfection, cells were incubated in media containing 500 uM CuSO4 (final) to induce
ectopic expression of target proteins and harvested 24 h later. Extracts were prepared and either subjected to immunoprecipitaion (IP) or analyzed directly (input). Immune complexes were recovered with anti-dPER(GP339) (added specific antibody) or anti-HA (added nonspecific antibody) antibodies. Immunoblots were probed with antibodies against anti-V5 (to detect dCLK), or anti-dPER (to detect dCLK), as indicated.
F. dPER 의 아미노산 949-977 를 Ser 과 Thr 을 모두 Ala 으로
바꾸어도 DBT 에 의한 인산화가 일어난다.
dPER(Δ949-977) 이 전사 억제제로서의 활성을 가지지 못하는데 이 단백질이 DBT 에 의해 인산화가 원활히 일어나지 못하는 것이 기여하는 바가 있는가를 확인하기 위하여 먼저 아미노산 949-977 부위에 DBT 에 의해 인산화가 되는 아미노산 잔기가 있는가를 확인하고자 하였다. 이를 위하여 dPER 의 아미노산 949-977 중 Ser/Thr 을 Rapid and efficient PCR-based mutagenesis method (Ko 등, 2007) 를 이용하여 모두 Ala 으로 변형시켰다. 이를 pAct-dper(949-977_9A)-V5 라 명명 하였다 (Fig. 11). 제작한 pAct-dper(949-pAct-dper(949-977_9A)-V5 를pMT-dbt-V5 와 S2 cell 에 cotransfection 하여 western blot 을 수행한 결과 (Fig. 12) 이
부위의 모든 Ser/Thr 을 Ala 으로 치환하였을 때에는 dPER(1-1224)에 비해 dPER(949-977_9A) 가 약간 더 인산화되는 것으로 보이지만 dPER(1-1224)와 매우 유사하게 정상적으로 인산화 되는 것을 볼 수 있었다. 적어도 아미노산 949-977 에는 DBT 에 의해 인산화 되는 잔기가 존재하지 않을 것임을 암시한다. 그러나 아미노산 949-977 제거에 의해 이 부위에 인접한 Ser/Thr 잔기의 인산화에 영향을 미쳤을 가능성을 배제할 수는 없다. 아미노산 949-977 의 부위는 아니지만 이미 보고 (Chiu 등, 2008) 한 바에 의하면 Ser976, Thr978, Ser981 이 S2
세포에서 DBT 에 의해 인산화되는 잔기임이 알려져 있다. 따라서 아미노산 949-977 의 제거에 의해 Ser976, Thr978, Ser981의 인산화에 영향을 미쳐 dPER(Δ949-977)
Fig. 11. All 9 serines and threonines in dPER aa 949-977was switched to alanines. (A) Shown is dPER amino acid sequence from 949 to 977. *, Ser and Thr residues that were mutated to Ala to generate the dPER(949-977_9A) version of dPER. (B) 9 Ser/Thr were mutated to Ala by using „A rapid and efficient PCR-based mutagenesis method‟ as indicated in materials and methods. Shown is the sequence alignment of wild-type dPER and 9A mutated version of dPER.
Fig. 12. DBT-dependent phosphorylation and degradation of dPER(949-977_9A) occur in the similar extent as those of wild-type dPER in S2 cells. S2 cells were transfected with 600 ng of V5-tagged versions of dper-containing plasmids. The presence (+) of 200 ng of pMT-dbt-V5 is indicated. Exogenous DBT was induced 36 h after transfection by adding to the media 500 uM CuSO4 (final). Cells were harvested at the indicated times, and extracts
were analyzed by immunoblotting in the presence of anti-V5 antibodies to visualize dPER (A and B). dPER(1-1224) and dPER(949-977_9A) extracts were loaded different density (1 : 2) after dPER(949-977_9A) to compare side by side with dPER(1-1224) (B).
Ⅳ. 고 찰
모든 생물은 생체시계를 가지며 이것을 이용하여 하루 중 생체리듬을 조절한다. 그 중에서도 분자생물학적 유전학적 연구가치가 뛰어나며 인간과 상동성이 높은 초파리 동물모델 생체시계의 transcriptional feedback loop 중에서 중요한 역할을 수행하는 dPER 의 분자생물학적 생화학적 역할을 규명하는 것은 생체시계 연구분야에서 매우 중요한 연구이다. dPER 의 서열 중 종간 보존성이 매우 높은 C4 (아미노산 926-977) 부위의 역할과 그러한 역할을 하는 보다 정확한 부위를 찾고자 하였다. 그 결과 우리는 아미노산 926-977 중에서도 중요한 역할을 하는 부위를 아미노산 949-977 로 줄일 수 있었고 이 부위가 제거되었을 경우 dPER 가 가지는 고유의 역할인 전사 억제제 역할을 잃어버리는 것을 확인할 수 있었으며, DBT 에 의한 인산화가 정상적으로 이루어지지 않았고, dCLK 과 결합하지 못하는 것을 발견하였다. 이 때 DBT 에 의한 인산화가 이루어진다고 알려진 Ser 과 Thr 을 Ala 으로 치환하여 dPER(949-977) 중에서도 DBT 에 의해 인산화되는 부위를 찾고자 하였으나 dPER(1-1224) 보다 약간 더 인산화 되는 듯한 패턴을 보였지만 wild-type 과 유사한 인산화 패턴을 보인다는 것은 적어도 아미노산 949-977 에는 DBT 에 의해 인산화되는 아미노산이 존재하지 않는다는 것을 의미한다.
한 편 in vitro 뿐만 아니라 in vivo 결과에서도 중요한 의미를 가진다. dper+
gene 을 발현하는 형질전환 초파리는 약 23.2 시간의 주기를 가지며 rhythmic 한 반면 dPER(Δ926-977) 을 발현하는 형질전환 초파리의 독립적인 라인에서 모두
주기가 약 3.7 ~ 4.3 시간 길어졌지만 정상적인 생체리듬을 가지는 것을 알 수 있었다. 초파리 세포배양에서 확인한 바에 의하면, dPER(Δ926-977) 는 dCLK 과 직접적으로 결합할 수 있는 능력을 잃어버려 dCLK 의 전사활성을 억제하지 못하였다. DBT 와 결합하는 도메인이 없는 dPER(ΔdPDBD) 를 발현하는 형질전환 초파리가 행동학적으로 생체리듬을 전혀 나타내지 못하는 반면, dPER(Δ926-977)을 발현하는 형질전환 초파리는 비록 주기가 길어지기는 하였으나 정상적인 리듬을 갖는 다는 사실은 매우 흥미롭다. 이는 dPER(Δ926-977) 단백질의 분자적ㆍ생화학적 결함이 in vivo 에서는 다를 수 있다는 것을 시사하는 것으로 이에 대한 연구는 앞으로 더욱 진행되어야 할 것이다. 이러한 dPER 의 아미노산 926-977 의 역할을 현재 기술한 논문에서 전사활성, 단백질의 변화, 그리고 행동학적 측면에서 관찰하였는데 앞으로의 또 다른 접근의 연구가 필요할 것이라고 생각된다. 그래서 우리는 첫 째, 형질전환 초파리 생체 내 RNA level 을 확인할 것이다. 둘 째, dCLK 과 결합하지 못하는데도 불구하고 dPER(Δ926-977) 은 초파리 생체 내에서 생체리듬의 주기가 길어지긴 했지만 정상적인 생체리듬을 가진다는 것은 dPER(Δ926-977) 과 dCLK 을 간접적으로 도와주는 단백질이 있을 수 있을 가능성이 있다. 예측되는 후보 단백질로는 dPER 와 이형이합체를 이루는 TIM 과 우리가 아직 알지 못하는 또 다른 factor 가 관여하고 있을 가능성을 조사할 것이다. 현재 진행 중에 있는 이러한 연구들이 결실을 맺게 된다면 생체시계 연구분야의 생화학적 분자생물학적 기전을 이해하는데 매우 중요한 역할을 할 것이라 생각된다.
Ⅴ. 결 론
모든 생명체는 생체시계를 가지고 있으며, 생체시계의 핵심기전은 “interlocked transcriptional feedback loop” 으로서 두 개의 loop 이 서로 맞물려 생체시계의 핵심 유전자들의 발현이 하루를 주기로 진동할 수 있도록 한다. 초파리의 경우 종간 단백질의 서열을 비교하였을 때 dPER 의 CCID (dCLK:CYC
Inhibition Domain) 내에서 아미노산 768-842 와 아미노산 926-977 이 잘 보존되어
있었다.
앞서 발표된 논문에서 아미노산 768-842 의 역할이 밝혀졌고 (Kim 등 2007), 우리는 아미노산 926-977 의 역할을 밝혀내기 위해 연구하였다. dPER 에서 아미노산 926-977 이 제거된 plasmid 를 만들었고, 이 것을 이용하여 S2 세포에서 luciferase assay 를 통해 조사해 본 결과 전사활성을 억제하지 못했다. dPER(Δ926-977) 이 전사활성을 억제하지 못한다는 것은 아미노산 926-977 이 제거됨으로 인해 dPER 가 핵 속으로 이동하지 못해서 일어나는 현장이 아니라는 것을 NLS 서열을 붙여 다시 한 번 luciferase assay 를 수행함으로써 알 수 있었다. 또한 전사억제 활성을 가지지 못하는 이유를 dPER(Δ926-977) 과 dCLK 의 결합력을 조사하기 위해 IP 를 수행한 결과 dCLK 과 결합하지 못했으며 S2 세포에서 DBT 에 의한 인산화를 조사해본 결과 hyperphosphorylation 이 매우 더디게 나타나며 분해되지 못한다. dCLK 과 결합하지 못하고 전사억제 활성을 가지지 못하게 하며 인산화되지 못하는 dPER(Δ926-977) 부위를 좀 더 좁히기 위해 dPER(Δ926-948), dPER(Δ949-977) 로 두 부분으로 나누어서 동일한 실험을