이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Role of BTG2/TIS21/PC3 in differentiation
of myeloid leukemia cells and its regulation
under stress conditions
by
Muhammad Imran
Major in Molecular Medicine
Department of Biomedical Sciences
The Graduate School, Ajou University
Role of BTG2/TIS21/PC3 in differentiation
of myeloid leukemia cells and its regulation
under stress conditions
by
Muhammad Imran
A Dissertation Submitted to The Graduate School of
Ajou University in Partial Fulfillment of the Requirements
for the Degree of
Ph.D. in Biomedical Sciences
Supervised by
In Kyoung Lim M.D., Ph.D.
Major in Molecular Medicine
Department of Biomedical Sciences
The Graduate School, Ajou University
This certifies that the dissertation
of Muhammad Imran is approved.
SUPERVISORY COMMITTEE
The Graduate School, Ajou University
November 27
th, 2013
i
- ABSTRACT-
Role of BTG2/TIS21/PC3 in differentiation of myeloid leukemia
cells and its regulation under stress conditions
Mouse TIS21 (12-O-tetradecanoyl phorbol-13-acetate inducible sequence 21), an ortholog of human BTG2 (B-cell translocation gene 2) and rat PC3, is a tumor suppressor that belongs to antiproliferative gene family, and is implicated in a variety of biological processes. Deregulation of c-Myc transcription factor is common in leukemia and lymphomas; the tumors are highly proliferative and often blocked at an earlier phase than the terminal stage of differentiation. The interrelation and the functional interplay of these two different proteins are not defined yet. We have shown here that the tumor suppressor TIS21 negatively regulated c-Myc expression during all-trans-retinoic acid (ATRA)-induced differentiation that accelerated differentiation and reduced proliferation of acute promyelocytic leukemia HL-60 cells. TIS21 downregulated c-Myc mRNA and additionally decreased c-Myc protein stability by increasing its phosphorylation at S62 andT58 residues
via activation of Erk1/2 and inhibition of PI3K/Akt along with subsequent activation of
GSK-3β in response to low dose ATRA treatment. HL-60 cells treated with GSK-3β or proteosome inhibitors revealed marked accumulation of c-Myc both in the presence and absence of ATRA and TIS21, confirming ATRA plus TIS21 mediated c-Myc phosphorylation and its consequent degradation in proteosome. Immunoprecipitation assay revealed that TIS21 hindered interaction of p-Erk1/2 with Akt, thus directly regulating MAPK and Akt activities without interaction with c-Myc. These findings exhibit anticarcinogenic potential of TIS21 via downregulation of c-Myc expression during ATRA induced differentiation of HL-60 cells involving activation and deactivation of two major c-Myc regulators Erk1/2 and
ii
Akt, respectively.
Recently, we have reported transient induction of Btg2 expression in response to oxidative damage; however, the regulatory mechanism was not explored. In the present study we revealed F- as the upstream mediator involved in Btg2 transcription in response to cell stress challenges such as serum deprivation and oxidative stress i.e. H2O2, or
doxorubicin treatments. We observed close interrelation between generation of reactive oxygen species (ROS), enhanced IBα degradation, nuclear translocation of F- (p65/RelA) and the significant increase of Btg2 expression independent of p53 status. ChIP analysis revealed an enrichment of RelA (p65) bound to the B response element on Btg2 promoter in response to the cell stress challenges. Employing various inhibitors led to cytoplasmic accumulation of IBα, decreased p65 nuclear translocation along with significant reduction of Btg2 expression. Generation of ROS was the common event mediating F- activation and Btg2 transcription. Furthermore, PKC activation was also found to be a critical factor mediating ROS-mediated signals to NF-B pathway that culminate on Btg2 regulation, and specifically PKC- was responsible for this regulation under oxidative stress. Serum deprivation-associated ROS generation bypassed PKC activation, however, regulated NF-B-Btg2 cascade via MAPK activation. The present data imply that oxidative stress upregulates Btg2 expression via ROS-PKC-F or ROS-MAPK-F cascade, independent of p53 status that in turn could be involved in mediating various biological phenotypes depending on the cellular context.
NF-κB plays crucial roles in inflammation and immunity and its activation is an important event for macrophage differentiation both in vivo and in vitro. In contrast
iii
involvement. Here we report that NF-κB activation enhanced and switched ATRA induced-granulocytic programme to macrophages. Serum withdrawal and LPS treatment dampened IκBα expression via MAPK activation and ATRA treatment further corroborated this effect in HL-60 cells. The data revealed that NF-κB activation diverted ATRA induced granulocytic differentiation to macrophages as confirmed by microscopic examination and assessing the macrophages specific markers CD68 and MMP9 along with high level of Btg2. Employing various inhibitors attenuated NF-κB associated enhanced cells maturation and differentiation switch thus suggesting that NF-κB determines the lineage specificity of ATRA induced differentiation of myeloid leukemia cells. MAPK activation is important both for granulocytic and macrophage differentiation and the data revealed that MAPK- NF-κB signaling was an important event in differentiation switch. The study shows that NF-κB plays an important role in determining lineage specificity of ATRA induced differentiation of myeloid leukemia cells.
__________________________________________________________________________ ATRA, all-trans-retinoic acid; Btg2, B-Cell translocation gene 2; GSK3, glycogen synthase kinase 3; DCF, dichlorofluorescin diacetate; LPS, Lipopolysaccharide; NAC, N-acetyl cysteine; PC3, pheochromocytoma cell-3; ROS, reactive oxygen species; TIS21 (12-O-tetradecanoyl phorbol-13-acetate inducible sequence 21)
iv
TABLE OF CONTENTS
ABSTRACT ··· i
TABLE OF CONTENTS ··· iv
LIST OF FIGURES ··· viii
Part 1 I. INTRODUCTION ··· 1
II. MATERIALS AND METHODS ··· 7
1. Cell culture ··· 7
2. Adenoviral transduction of HL-60 cells with TIS21 gene ··· 7
3. Differentiation analyses ··· 7
4. RNA extraction, semiquantitative RT-PCR and real time PCR ··· 8
5. RNA Interference ··· 10
6. Immunoblot and immunoprecipitation analyses ··· 10
7. Statistical Analyses ··· 10
III. RESULTS ··· 11
A. ATRA-induces granulocytic differentiation of HL-60 cells ··· 11
B. ATRA upregulates Btg2 expression in HL-60 cells ··· 14
C. TIS21 enhances ATRA-induced differentiation of HL-60 cells ··· 16
D. TIS21 enhances ATRA-induced differentiation via down-regulation of c-Myc in HL-60 cell ··· 19
E. TIS21 decreases stability of c-Myc protein in response to ATRA treatment ··· 25
F. Transduction of shTIS21 abrogates TIS21 effect on c-Myc expression and HL-60 cells differentiation ··· 27
v
G. ATRA plus TIS21 increased Erk1/2 activity, but inhibited Akt with subsequent
GSK-3β activation ··· 29
H. TIS21 enhanced downregulation of c-Myc by activating GSK-3β ··· 36
I. TIS21 enhanced ATRA-mediated c-Myc degradation in the proteosome ··· 39
IV. DISCUSSION··· 42
V. CONCLUSION ··· 45
Part 2 I. INTRODUCTION ··· 46
II. MATERIALS AND METHODS ··· 50
1. Cell culture ··· 50
2. RNA extraction and semi quantitative RT-PCR ··· 50
3. Immunoblot analysis ··· 52
4. Cells fractionation ··· 52
5. Measurement of intracellular ROS level and cell cycle analysis ··· 52
6. Chromatin immunoprecipitation (ChIP) assay ··· 53
7. Cells Proliferation Assay ··· 53
8. Transfection of PKC- ··· 8. Statistical analysis ··· 54
III. RESULTS ··· 55
A. Btg2 is upregulated under serum deprivation ··· 55
B. Serum deprivation-induced reactive oxygen species generation upregulates Btg2 expression ··· 60
C. Serum deprivation induces NF-B activation ··· 63
vi
E. SP1 and DNA damage signals do not regulate Btg2 expression under serum
deprivation ··· 69
F. Exogenous H2O2 regulates Btg2 expression via NF-B activation in DLD-1 cells ···· 71
G. Doxorubicin induces Btg2 expression via ROS-NF-B pathway ··· 73
H. ROS regulate NF-B activation along with Btg2 expression via PKC activity ··· 76
I. PKC- regulates NF-B activity and Btg2 expression in ROS dependent manner ···· 81
J. ROS regulate NFB-Btg2 under serum deprivation via MAPK pathway activation ·· 83
K. Btg2 reduces cells proliferation expression under serum deprivation, H2O2 and Doxo treatment ··· 86
IV. DISCUSSION··· 89
V. CONCLUSION ··· 93
Part 3 I. INTRODUCTION ··· 94
II. MATERIALS AND METHODS ··· 97
1. Cell culture ··· 97
2. Differentiation analyses ··· 97
3. RNA extraction, semiquantitative RT-PCR and real time PCR ··· 97
4. Immunoblot and immunoprecipitation analyses ··· 98
5. Cells fractionation ··· 98
6. Measurement of intracellular ROS level ··· 99
7. Statistical Analyses ··· 99
III. RESULTS ··· 100
A. ATRA induces macrophage differentiation under reduced serum concentration ··· 100
vii
C. Inhibition of NF-B abrogates ATRA induced macrophage differentiation of
HL-60 cells··· 107
D. Activation of NF-B induces macrophage differentiation in response to ATRA Treatment ··· 109
E.ATRA enhances NF-B activation and induces macrophage differentiation via MAPK ··· 112
F. ATRA induced C/EBPα expression via MAPK is lower in cells under serum deprivation ··· 115
IV. DISCUSSION··· 118
V. CONCLUSION ··· 121
VI. REFERENCES ··· 122
viii
LIST OF FIGURES
Fig. 1. c-Myc structure and degradation Mechanism ··· 4
Fig. 2. ATRA induces granulocytic differentiation of HL-60 cells ··· 12
Fig. 3. ATRA upregulates Btg2 expression in HL-60 cells ··· 15
Fig. 4. TIS21-enhances ATRA induced differentiation of HL-60 cells ··· 17
Fig. 5. TIS21 enhances c-Myc degradation in response to ATRA ··· 21
Fig. 6. TIS21 decreases stability of c-Myc protein in response to ATRA ··· 26
Fig. 7. Knockdown of TIS21 gene abrogates the enhanced effects on c-Myc down-regulation and HL-60 cells differentiation ··· 28
Fig. 8. TIS21 enhances ATRA-mediated activation of Erk1/2, and inhibited PI3K/Akt pathway along with GSK-3β activation ··· 31
Fig. 9. TIS21-enhanced c-Myc degradation was regulated by GSK-3β activation ··· 37
Fig. 10. TIS21 enhances ATRA-mediated c-Myc degradation in the proteosome ··· 40
Fig.
11
. Schematic representation of c-Myc degradation ··· 41Fig.
12
. The NF-B protein family members ··· 47Fig. 1
3
. Mechanisms of NF-Bactivation ···
48Fig. 14. Btg2 expression is up-regulated upon serum deprivation ··· 57
Fig. 15. Reactive oxygen species generated by serum deprivation regulates Btg2 expression ··· 61
Fig. 16. Serum deprivation induces NF-B activation via IBα degradation ··· 64
Fig. 17. NF-B regulates Btg2 expression under serum deprivation ··· 67
Fig. 18. SP1 and DNA damage signals do not regulate Btg2 expression under serum deprivation ··· 70
ix
Fig. 19. Exogenous H2O2 regulates Btg2 expression via NF-B activation in DLD-1 cells
··· 72
Fig. 20. Doxorubicin regulates Btg2 expression via ROS-NFB cascade ··· 74
Fig. 21. PKC mediate ROS-NFB-Btg2 cascade ··· 78
Fig. 22. PKC- isozyme regulates Doxo- and H2O2-induced Btg2 expression ··· 82
Fig.
23
. ROS regulate NFB-Btg2 under serum deprivation via MAPK pathway activation ··· 84Fig.24. Btg2 reduces cells proliferation of cells upon exposure to serum deprivation, H2O2 and Doxo ··· 87
Fig. 25. Schematic representation of the regulatory mechanism of Btg2 expression ··· 88
Fig. 26. Schematic representation of hematopoietic stem cells differentiation ··· 95
Fig.
27
. ATRA induces macrophage differentiation under reduced serum concentration ·· ··· 101Fig. 28. ATRA enhances NF-
B
activation of myeloid leukemia cells under serum deprivation ··· 105Fig. 29. Inhibition of NF-
B
abrogates ATRA induced macrophage differentiation of HL-60 cells··· 108Fig. 30. Activation of NF-B by LPS treatment induces macrophage differentiation in response to ATRA treatment ··· 110
Fig. 31. ATRA activates NF-
B
and induces macrophage differentiation via MAPK pathway regulation ··· 113Fig. 32. ATRA induced C/EBPα expression via MAPK is much lower in cells under serum deprivation ··· 116
x
Fig. 33. Schematic representation of the regulatory mechanism of ATRA enhanced NF-
B
activation and macrophage differentiation ··· 117 Table 1 ··· 9 Table 2 ··· 51- 1 -
Part-1
Role of Btg2/TIS21/PC3 in differentiation of myeloid leukemia cells
I. INTRODUCTION
A. Acute Myeloid Leukemia
Acute myeloid leukemia (AML), also known as acute myelogenous leukemia or acute nonlymphocytic leukemia (ANLL) is a heterogeneous group of hematologic malignancies characterized by a differentiation block in hematopoietic progenitor cells at the early stages of myelopoiesis and proliferation of immature blasts (Zpolat et al. 2007). The French-American-British (FAB, 1970) classification system divides AML into eight subtypes, M0 through to M7, based on the type of cell from which the leukemia developed and its degree of maturity. The WHO classification (2002) system divides AML into several broad groups on the basis of genetic abnormalities and immmunophenotypic features (James et al. 2002). Although the pathophysiology of AML has been studied extensively, however, the expanding pool of therapeutic approaches to the disease have not yet significantly improved the disease-free or overall survival associated with any form of AML except for acute promyelocytic leukemia (APL) (Ozpolat at el. 2008). All-trans retinoic acid (ATRA) serves as a useful pharmacological agent in chemoprevention and differentiation therapy of several types of cancers (Lotan et al. 1996, Altucci et al. 2001, Soprano et al. 2004) with particularly impressive results in the management of acute promyelocytic leukemia (Melnick et al. 1999). These pleiotropic activities are exerted primarily through the ability of ATRA to regulate gene expression and are mediated by the nuclear hormone receptors termed retinoic acid receptors (RARα, RARβ, RARγ, and their isoforms) and the RXRs (RXRα, RXRβ, and RXRγ and their isoforms). These receptors are ligand-activated transcription factors.
- 2 -
Liganded RAR-RXR heterodimers or RXR-RXR homodimers bind to retinoic acid response elements (RARE) and transduce the RA signal to the transcription machinery and the chromatin template to activate or repress the transcription of target genes (Eckhardt et al. 1994).
- 3 -
B. c-Myc
c-Myc is a nuclear protein which has been implicated in the control of normal cell growth as well as transformation and is implicated in a wide range of human cancers. Myc transcription factor is involved in cell cycle regulation, cells adhesion and migration, angiogenesis, cell growth and metabolism, apoptosis, genomic instability, transformation and blocks cellular differentiation (Bentley et al. 1986, Amati 2004). Unregulated c-myc expression is common in some leukemias and lymphomas; these tumor cells are highly proliferative and are often blocked at an earlier stage than the terminal stage of differentiation (Ben-Baruch et al. 1994). Down-regulation of c-myc mRNA expression is linked to growth and the differentiation arrest of hematopoietic cells whereas exogenous expression of c-myc impairs these phenomena (Dimberg et al. 2002). AML HL-60 cells, for example, have an extrachromosomally amplified myc gene that inhibits cells maturation. c-Myc, however, is downregulated during ATRA induced differentiation (Eckhardt et al. 1994). c-Myc expression is regulated through transcriptional, translational and posttranslational mechanisms. Regulation of transcriptional elongation of c-Myc mRNA during myeloid maturation has been well established (Bentley et al. 1986, Bentley et al. 1988, Eick et al. 1990). Posttranslational modifications and phosphorylation-dependant degradation processes are mediated in the Myc-binding domain I; Phosphorylation on S62 by Erk1/2, JNK or Cdk5 creates proliferative stimuli with increased stability of Myc protein, which deserves as a priming event for T58 phosphorylation by GSK3β (Welcker et al. 2008, Sears 2004, Noguchi et al. 1999) PP2A dephosphorylates p-cMycS62/T58 in Pin-1 dependent manner, and p-cMycT58 is then recognized by SCF-Fbw7 that is responsible for proteosome-mediated degradation (Arnold et al. 2008). However, this posttranslational regulation of c-Myc during ATRA induced differentiation of APL cells is not well studied.
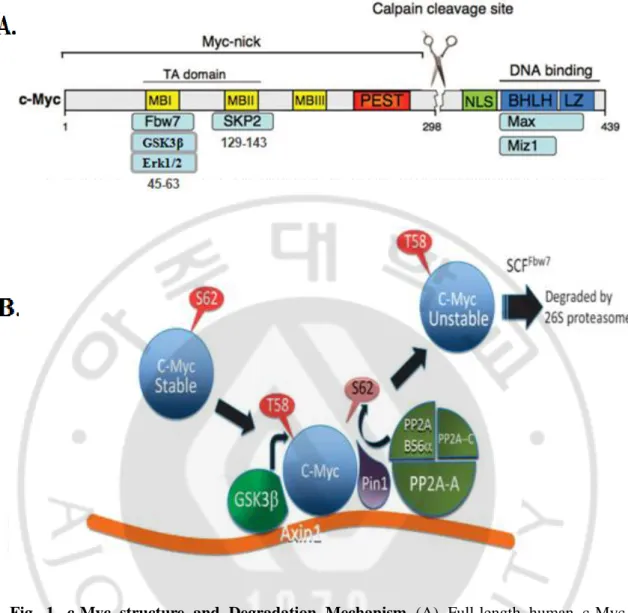
- 4 -
Fig. 1. c-Myc structure and Degradation Mechanism (A) Full-length human c-Myc
protein and its binding partners. The scissors indicate the major calpain cleavage site within human c-Myc. Myc boxes (MB) and Basic Helix Loop Helix Leucine Zipper (BHLH LZ) domains are highly conserved among Myc family members. NLS, nuclear localization sequence; PEST, region containing amino acids often found in unstable proteins (Modified from Conacci-Sorrell et al. Cell Cycle 10 (2011) 604-610) (B) Model showing sequential steps in c-Myc degradation and the role of Axin1 as a scaffold in this process. (Adopted from Arnold et al. EMBO J. 28 (2009) 500-512).
- 5 -
C. BTG2/TIS21/PC3
TIS21 (12-O-tetradecanoyl phorbol-13-acetate (TPA)-inducible sequences 21) in mouse, identified as a primary-response gene in mouse 3T3 fibroblasts treated with 12-O-tetradecanoyl phorbol-13-acetate (TPA), forskolin, and epidermal growth factor and is a member of antiproliferative (APRO) gene family ((Lim et al., 1987, Fletcher et al., 1991) along with BTG1 (Rouault et al., 1996), BTG3 (Guehenneux et al., 1997), and Tob (Matsuda et al., 1996) genes. PC3 (pheochromocytoma cell-3) found in rat PC12 cells and BTG2 (B-cell translocation gene 2) in humans (Bradbury et al., 1991 Rouault et al., 1996, Young-Bin et al., 2008, Yang et al., 2008, Melamed et al., 2002) are TIS21-homologous genes. Expression of TIS21 BTG2/PC3 is constitutively high in thymus, lung alveolar epithelium, spleen, lymph node, proximal tubule of kidney, and basal cell layer of prostate acini (Melamed et al., 2002). Moreover, the expression is much less in tumor than in the surrounding tissues and related to carcinogenesis in thymus, prostate acini, renal proximal tubule, and liver, suggesting its role as a tumor suppressor (Lim et al., 1995, Struckmann et al., 2004, Ficazzola et al., 2003, Park et al., 2008).
TIS21 BTG2/PC3 has been implicated in the regulation of a variety of biological processes (Duriez et al., 2004, Lim, 2006). For instance, TIS21 can act as: a transcriptional co-regulator (Canzoniere et al., 2004, Farioli-Vecchioli et al., 2009); an anti-apoptotic factor in neurogenesis (Tirone et al., 2001, Konrad et al., 2005); a key mediator of the stage specific expansion of developing thymocyte (Oswald et al., 2006); a negative regulator of hematopoietic progenitor expansion (Ryo et al., 2005); a tumor suppressor gene in both mouse and human [Lim et al., 1995, Elmore et al., 2004, Guardavaccaro et al., 2000); a pan cell cycle modulator (Lim et al., 1998, Hong et al., 2005, Park et al., 2004); induction of
- 6 -
cellular senescence and a developmental regulator (Sugimoto et al., 2005, Sakaguchi et al., 2001, Corrente et al., 2002). Furthermore, TIS21 BTG2/PC3 enhances differentiation in neuronal cells (Cho et al., 2004) by inhibiting cell cycle progression as well as contributing towards the activation of proneural genes in neural progenitor cells (Canzoniere et al., 2004). Several studies strongly suggest the involvement of BTG2 in the stress-mediated phenotypes [Fiedler et al., 1998, Savkovic et al., 2004, Slevin et al., 2009). Depending on the type of cytotoxic and genotoxic stresses, BTG2 expression is regulated by p53, NF-κB, SP1 or other transcription factors, and the binding sites for various transcription factors including SP1, GATA1, CREB, p53 and NF-κB are located within the BTG2 promoter [Rouault et al., 1996, Duriez et al., 2002).
D. Purpose of this study
TIS21 BTG2/PC3 enhances differentiation in neuronal cells by inhibiting cell cycle progression as well as contributing towards the activation of proneural genes in neural progenitor cells. Contrary to neuronal cells, however, very little is known about the role of TIS21 in the differentiation of hematopoietic cells. In the current study, we attempted to explore the possible involvement of TIS21 during retinoic acid induced differentiation of HL-60 cells, with regard to c-Myc down-regulation. Our results revealed
- 7 -
II. MATERIALS AND METHODS
A. MATERIALS
Antibodies against pc-Myc, pAkt, Akt, pErk1/2, Erk1/2, pGSK3β, GSK3β, pJNK, JNK, PARP and actin were purchased from Cell Signaling; anti-CD11b was from Abcam Laboratories; c-Myc, Pin1, PP2A, Skp2, p21, cyclin-A, cyclin-D1, cyclin-E, Bcl2, GAPDH and α-tubulin were from Santa Cruz Biotechnology and p27 from Epitomics. ATRA, LY294002, U0126, LLnL, lithium chloride, cycloheximide, and 1,5-dimethyl-1,5-diazaundecamethylene polymethobromide (polybrene) were purchased from Sigma Chemicals
B. METHODS
1. Cells culture
HL-60 and NB4 cells were cultured in RPM1 1640 (Sigma, St. Louis USA) supplemented with 10% heat inactivated FBS and 1% penicillin and streptomycin at 37 °C under 5% CO2 in air.
2. Adenoviral transduction of HL-60 cells with TIS21 gene
Ad-TIS21 virus prepared in our laboratory has been described in our previous reports Lim et al. 2008, Park et al. 2008). Infection of Ad-TIS21 virus was performed in the presence of 10 μg/ml polybrene as described previously (Clark et al. 1999). Transduction of Ad-LacZ was employed as control.
3. Differentiation analyses
To monitor granulocytic differentiation, HL-60 cells (2×105 cells/ml) were treated with ATRA in the presence and absence of Ad-TIS21. Differentiation of HL-60 was
- 8 -
evaluated by measuring CD11b and CD38 expression using FACS analysis with PE-conjugated anti-CD11b and anti-CD38 antibodies or anti-mouse IgG1κ (BD Biosciences). Expressions of CD11b and CD38 were compared to that of control. Morphologic changes were evaluated by May-Grunwald Giemsa stain (Sigma-Aldrich, Germany) according to the manufacturer’s instruction. For proliferation ATRA-treated HL-60 cells (2x105/ml) were counted microscopically at various time points using a hemocytometer both in the presence and absence of TIS21. Cell cycle profile was determined by flow cytometry according to the manufacturer’s instruction (BD Biosciences).
4. RNA extraction, semiquantitative RT-PCR and real time PCR
RNA was extracted with Trizol Kit (Invitrogen) from HL-60 cells treated with ATRA (0.1-1.0 μM) in the presence or absence of Ad-TIS21. One μg of total cellular RNAs were used for reverse transcription of cDNAs and gene of interest was amplified by semiquantitative RT-PCR or real-time PCR. Real-time PCR was carried out for c-Myc gene expression with PowerSYBR Green PCR Master Mix (Applied Biosystems) using the following conditions; Initial activation at 95 oC for 5 min, followed by 40 cycles of 95oC for 15 sec and 60oC for 1 min, and the fluorescence was determined at the melting temperature of the product for 20 sec (ABI PRISM 7000 Sequence Detection System). The 2-ΔΔCT was used to analyze the data. Relative gene expression values were normalized to GAPDH. Primer sequences are described in Table 1.
- 9 -
Table 1. List of PCR Primers
RNA Direction Sequence
c-myc (Real Time) Forward Reverse
5 -TGTCCCACGGGACCTACTAC-3
5 -CCTCATGCTCACACATGTCC-3
Btg2 (Real Time) Forward Reverse5 -TCCCTGGAGAAGAGCTACCA-3
5 -AGGAAGGAAGGCTGGAAGAG-3
GAPDH (Real Time) Forward Reverse5 -TGGGCTACACTGAGCACCAG-3
5 -GGGTGTCGCTGTTGAAGTCA-3
Btg2 (RT-PCR) Forward Reverse5 -CCT GGG CAG AGA GTG AAA AG-3
5 -CCT TCC ATC CTA ACC CCA AT-3
TIS21 (RT-PCR) Forward Reverse
5 -ATGAGCCACGGGAAGAG-3
5 - CGGGGCCTCCTCATAC -3
GAPDH (RT-PCR) Forward Reverse5 -CCATGGAGAAGGCTGGGG-3
5 -CAAAGTTGTCATGGATGACC-3
- 10 -
5. RNA Interference
Lentiviral vectors targeting TIS21 gene (shTIS21: CCGGCTATCGCTGTATCCGTATCAACTCGAGTTGATACGGATACAGCGATAGTTT TTG) were purchased from Open Biosystems (Huntsville, AL.). Lentiviral particles were produced according to the manufacturer’s instruction; 293TN cells were transiently transfected with shTIS21 or viral vector along with envelope plasmid pVSV-G and pδ8.9 packaging plasmid. Culture media containing viralparticles were filtered (0.45μm) 48h after the transfection and the supernatants were used for further experiments.
6. Immunoblot and immunoprecipitation analyses
HL-60 cells were harvested and lysed in buffer containing 50 mM Tris, pH 7.4, 150 mM NaCl, 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride, 1 μg/ml aprotinin, 1 μg/ml leupeptin, 1 μg/ml pepstatin, 1 mM NaF, 1 mM sodium orthovanadate, 0.25% sodium deoxycholate, and 1 % NP40. After centrifugation at 12,000 x g for 15 min, supernatants were analyzed by PAGE and immunoblotting on polyvinylidine difluoride membrane (Millipore). To monitor the interaction of proteins, cells were lysed in buffer with 50 mM Tris-HCl, 150 mM NaCl, 1 % NP40, 0.50 % sodium deoxycholate and protease inhibitors, and subjected to immunoprecipitation and immunoblotting.
7. Statistical Analyses
All of the data was presented as mean ± SD. Each experiment was performed in triplicates. Statistical differences were analyzed by Student’s t-test or ANOVA test and p values <0.05 were regarded as statistically significant.
- 11 -
III. RESULTS
A. ATRA-induces granulocytic differentiation of HL-60 cells
To study the role of TIS21 in the granulocytic differentiation of HL-60
cells we first
examined
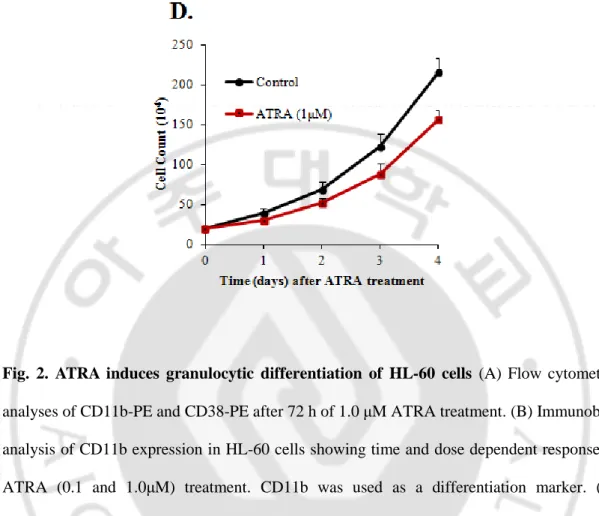
the surface expression of CD11b and CD38, markers of myeloid differentiation, in HL-60 cells in response to ATRA treatment. CD11b is a general differentiation marker expressed by many cell lineages while CD38 is granulocyte specific. HL-60 cells were treated with 1 μM ATRA for 72 h, and analyzed by fluorescence-activated cell sorting with anti-CD11b and anti CD38 antibodies (Fig. 2A). Flow cytometric data for surface markers expression showed that exposure of HL-60 cells to ATRA profoundly induced differentiation of these cells. Differentiation was also assessed in a time and dose dependent manner by Western blotting using CD11b as a marker. CD11b expression induced at 12 h for 1μM while it was detectable at 24 h for 0.1μM and increased regularly time dependently (Fig. 2B). We also assessed morphologic changes in the cells by May-Grünwald-Giemsa (MGG) staining. ATRA-treated cells underwent morphological changes characteristic of granulocytes with segmented nuclei, which revealed that the untreated HL-60 cells were predominantly promyelocytes with characteristic cytoplasmic granules, large nuclei, and prominent nucleoli (Fig. 2C). Moreover, ATRA treatment decreased cells proliferation (Fig. 2D).- 13 -
Fig. 2. ATRA induces granulocytic differentiation of HL-60 cells (A) Flow cytometric
analyses of CD11b-PE and CD38-PE after 72 h of 1.0 μM ATRA treatment. (B) Immunoblot analysis of CD11b expression in HL-60 cells showing time and dose dependent response to ATRA (0.1 and 1.0μM) treatment. CD11b was used as a differentiation marker. (C) Morphological changes of HL-60 cells during the granulocytic differentiation were analyzed by May-Grurwald-Giemsa (MGG) staining after 72 h of 1.0 μM ATRA treatment. (D) ATRA decreases proliferation of HL-60. HL-60 cells were treated with 1.0 μM ATRA and cells proliferation was determined by hemocytometry.
- 14 -
B. ATRA upregulates Btg2 expression in HL-60 Cells
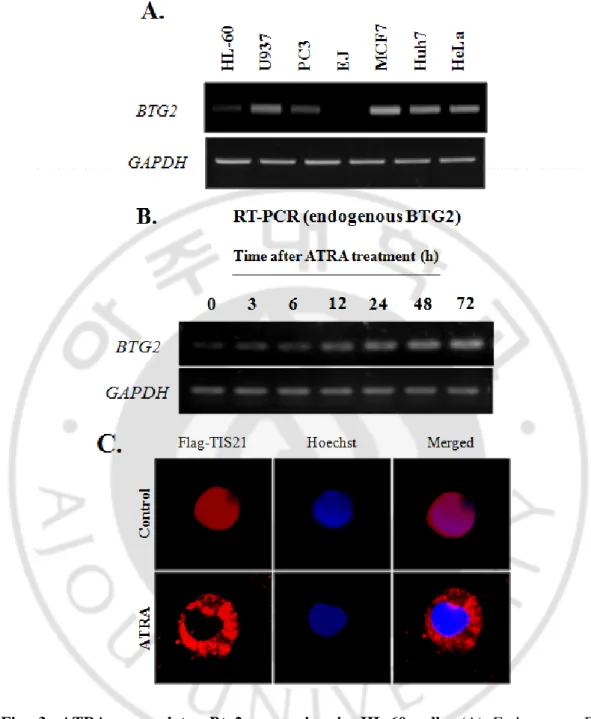
To study the role of TIS21 in the granulocytic differentiation of HL-60 cells we first examined the expression level of endogenous Btg2 in HL-60 cells by RT-PCR in comparison with a set of other cell lines. HL-60 cells were found to express low basal level of Btg2 higher than U937 and EJ cells while lower than HeLa and Huh7 cells (Fig. 3A). We next evaluated the effect of ATRA on Btg2 expression level. ATRA (1μM) treatment to HL-60 cells upregulated Btg2 expression in time dependent manner (Fig. 3B). This increased expression of Btg2 suggests that it might play a role in the differentiation process of myeloid cells. Moreover, ATRA treatment translocates Btg2 from nucleus to cytoplasm thus indicating a functional role for Btg2 in the differentiation event.
- 15 -
Fig. 3. ATRA upregulates Btg2 expression in HL-60 cells. (A) Endogenous Btg2
expression in various cancer cells in response to ATRA treatment. Btg2 expression was determined by RT-PCR. HL-60 cells express relatively low level of Btg2. (B) HL-60 cells were treated with ATRA (1.0μM) or DMSO for the indicated times, and Btg2 expression was evaluated by RT-PCR. GAPDH was used as loading control. (C) Btg2 translocation to cytoplasm after 24 h of ATRA treatment in HL-60 cells.
- 16 -
C. TIS21 enhances ATRA-induced differentiation of HL-60 cells
In order to find out the effect of TIS21on ATRA induced differentiation, HL-60 cells were infected with Ad-TIS21 or Ad-LacZ in time and manner. The effect of Ad-TIS21 on ATRA-induced differentiation in HL-60 cells was then evaluated by measuring the differentiation marker, CD11b and CD38 by FACS analysis. The expressions of surface markers, CD11b and CD38, were significantly enhanced by ectopic expression of TIS21 (Fig. 4A). TSI21 is an antiproliferative gene and thus reduces cells proliferation. The interactive effect between low dose ATRA and TIS21 significantly inhibited proliferation of HL-60 cells compared to TIS21 or ATRA alone treatment as well as the control from 1 day after ATRA treatment (Fig. 4B). The results thus demonstrate that TIS21 significantly enhanced differentiation of HL-60 cells to granulocytes. Interestingly we found that though TIS21 can induce differentiation in neuronal cells, but in APL cells it contributes only towards ATRA induced differentiation and enhances its effect, as TIS21 alone failed to increase expression of differentiation markers.
- 18 -
Fig. 4. TIS21-enhances ATRA induced differentiation of HL-60 cells. (A) HL-60 cells
were infected with adenovirus (200 moi) carrying either TIS21 or LacZ for 6 h. At 24h of posttransfection cells were treated with ATRA (0.1μM) or DMSO (for 24 and 48 h) and then subjected to FACS analysis for CD11b and CD38 expressions. (B) To assess the regulation of cells proliferation, HL-60 cells were treated with ATRA (0.1μM) after transduction of
TIS21 (200 moi) or LacZ (200 moi), and then cells proliferation was measured every 24 h
after ATRA treatment. Significant growth inhibition in the combined treatment of ATRA plus TIS21 was observed. The data presented as mean±SD of three independent experiments.
- 19 -
D. TIS21 enhances ATRA-induced differentiation via down-regulation of c-Myc in HL-60 cells
c-Myc deregulation is common in some leukemias and lymphomas; the tumors are highly proliferative and are often blocked at an earlier phase than the terminal stage of differentiation. HL-60 cells constitutively express c-Myc due to their characteristic extrachromosomally amplified c-myc gene. c-Myc is thus implicated in the differentiation block of HL-60 leukemic cells, so we attempted to establish the correlation between c-Myc down-regulation and differentiation of HL-60 cells in TIS21 dependent manner. The down-regulation of c-Mycis a key event in the process of terminal differentiation and growtharrest of acute promyelocytic leukemia cells. It is previously reported that ATRA-induced differentiationcells is associated with an early down-regulationof c-Myc. To investigate the effect of TIS21 gene on Myc expression in ATRA sensitive HL-60 cells, regulation of c-Myc expression was first examined. c-c-Myc level was reduced slightly until 72 h by 0.1 μM ATRA treatment as opposed to the robust decrease by 1.0 μM ATRA both at protein and mRNA levels (Fig. 5A, 5B). To investigate the TIS21 dependent c-Myc regulation and the effect of TIS21 on the ATRA-induced differentiation, HL-60 cells were treated with low dose ATRA (0.1 μM) in the presence and absence of Ad-TIS21 infection, and then harvested 48 h after ATRA treatment. ATRA alone minimally induced c-Myc protein downregulation and CD11b expression, however, combined treatment of ATRA with TIS21 significantly induced CD11b expression and downregulated c-Myc protein dose dependently (Fig. 5C), as opposed to no effect by TIS21 alone (Fig. 5D). The effect of TIS21 on c-Myc regulation was further clarified time dependently. When ectopic expression of TIS21 gene was achieved by infection of Ad-TIS21, expression of c-Myc protein was significantly reduced in 24 hours of 0.1 M ATRA (Fig. 5E), suggesting the enhanced effect of 0.1 μM ATRA on the reduction of
- 20 -
c-Myc expression by TIS21 gene. To clarify the immunoblot observation, relative values of c-Myc protein expression were calculated by densitometry. TIS21 reduced c-Myc protein expression over 80% of the control (Fig. 5E), whereas c-Myc mRNA expression was reduced to more than 50% of the control 24 h after ATRA treatment (Fig. 5F). Moreover, the expression level of exogenous TIS21 gene was independent of ATRA treatment (Fig. 5G). The data shows that TIS21 alone does not affect c-Myc expression, however, enhances ATRA-induced down-regulation of c-Myc in HL-60 cells and strongly suggest a possible regulation of c-Myc expression at the posttranscriptional level.
- 22 -
- 24 -
Fig. 5. TIS21 enhances c-Myc degradation in response to ATRA. (A) HL-60 cells were
treated with ATRA (0.1 and 1.0μM) or DMSO for the indicated times, and the change of c-Myc expression was evaluated by Western blot analysis. GAPDH was used as loading control. (B, C) HL-60 cells were infected with adenovirus carrying either TIS21 or LacZ in dose dependent manner (100 and 200 moi) for 6 h, and then cultured until 72 h until analysis without ATRA treatment (B) or with ATRA (0.1μM) treatment for 48 h (C). CD11b and c-Myc expressions were determined by Western blotting. α-Tubulin was used as a loading control. (D) HL-60 cells were infected for 6 h with adenoviruses carrying either TIS21 or LacZ gene (200 moi) as a transduction control and treated with low dose ATRA (0.1M) after 24 h of adenoviral transduction. Cells were harvested at the indicated times. Anti-Flag antibody was used to detect TIS21 expression and α-tubulin as a loading control. Graph represents densitometric analysis for c-Myc protein expression (E) TIS21 transduced cells were treated with ATRA (0.1M) for the indicated times and subjected to quantitative PCR analysis. The expression was normalized to GAPDH. The data represents mean ± SD of three independent measurements. (F) To assess the efficacy of transduced TIS21 gene expression in HL-60 cells, RT-PCR analysis was performed using the above cells at 12 h and 24 h after ATRA treatment.
- 25 -
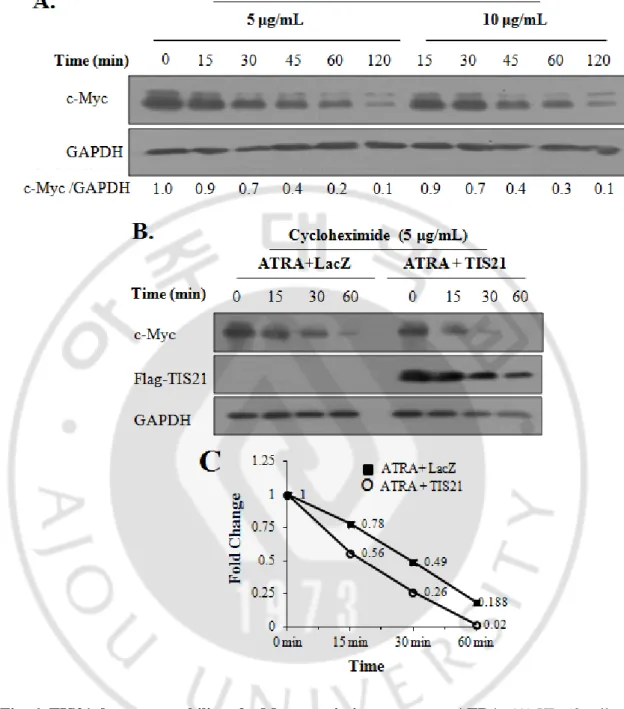
E. TIS21 decreases stability of c-Myc protein in response to low dose ATRA
To address the possibility of TIS21-dependent regulation of c-Myc protein expression, c-Myc protein turnover was first evaluated by using two different concentrations of cycloheximide. As shown in Fig. 6A, more than 50% of c-Myc protein was reduced within 45 min. To examine the effect of TIS21 on c-Myc degradation, TIS21 transduced HL-60 cells were pretreated with ATRA for 12 h and then treated with cycloheximide for 1 hour and c-Myc protein level was assessed at various time points. Expression of TIS21 accelerated degradation of c-Myc protein compared with that of LacZ expression (Fig. 6B). To further confirm the effect of TIS21 gene on c-Myc degradation, the immunoblot findings were analyzed by densitometry based on the expressions of GAPDH, and the effect of TIS21 on acceleration of c-Myc protein degradation was quantified (Fig. 6C).
- 26 -
Fig. 6. TIS21 decreases stability of c-Myc protein in response to ATRA. (A) HL-60 cells
were treated with cycloheximide (5 and 10 g/mL) to inhibit new protein synthesis and degradation of Myc protein was determined by Western blotting. (B) TIS21 gene was transduced to HL-60 cells by adenovirus infection for 6 h, followed by ATRA (0.1M) and cycloheximide (5 g/mL) treatment to the cells at 24 h and 36 h after viral transduction, respectively. Time indicates incubation after cycloheximide treatment. (C) c-Myc protein shown in Figure 2B was quantified by Image J.
- 27 -
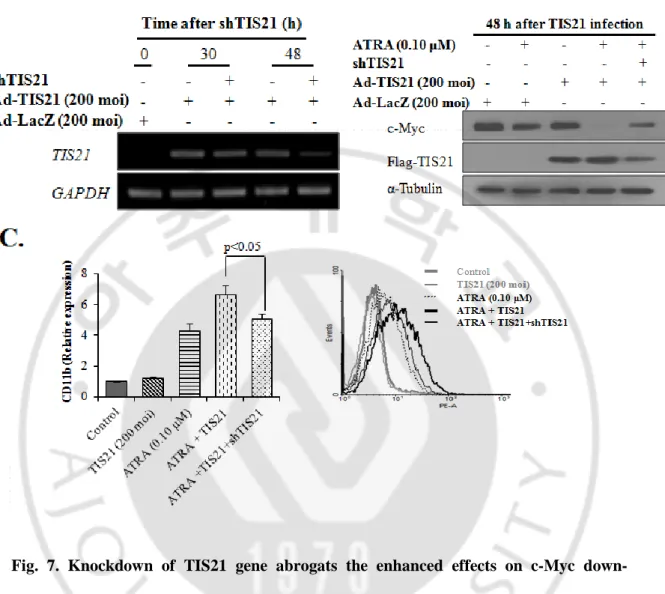
F. Transduction of shTIS21 abrogates TIS21 effect on c-Myc expression and HL-60 cells differentiation
To confirm the effect of TIS21 on c-Myc downregulation and differentiation of HL-60 cells, transduction of shRNA against TIS21 was employed by lentiviral vector. The efficiency and specificity of TIS21 knockdown was demonstrated by RT-PCR analysis (Fig. 7A). To allow down-regulation of target gene and its effect on c-Myc expression, cells were incubated with shTIS21 before transduction of Ad-TIS21. Twenty four hours after Ad-TIS21 transduction, cells were treated with ATRA (0.1M) for another 24 h and then subjected to Western blot and FACS analyses. Knockdown of TIS21 expression significantly diminished the synergistic effect of ATRA plus TIS21 on the degradation of c-Myc protein (Fig. 7B) and the differentiation of HL-60 cells (Fig. 7C), indicating the enhanced effect of TIS21 gene on HL-60 cells differentiation by downregulating c-Myc expression.
- 28 -
Fig. 7. Knockdown of TIS21 gene abrogats the enhanced effects on c-Myc down-regulation and HL-60 cells differentiation. (A) TIS21 was knocked down by lentivirus
infection carrying shTIS21 sequences. The efficiency and specificity of TIS21 gene knockdown was demonstrated by RT-PCR. GAPDH was used as loading control. (B) HL-60 cells were infected with lentivirus for 12 h followed by adenoviral transduction of TIS21 gene; At 24 h of TIS21 transduction cells were treated with ATRA (0.1μM) and then subjected to Western blotting. (C) Cells were transduced as described in (B), treated with ATRA (0.1μM) for 48 h and then subjected to FACS analysis for CD11b expression.
- 29 -
G. ATRA plus TIS21 increases Erk1/2 activity, but inhibits Akt with subsequent GSK-3β activation
To investigate the mechanism regulating ATRA-induced c-Myc expression along with differentiation of HL-60 cells in TIS21 dependent manner, MAPK and Akt pathways were evaluated by using inhibitors treatments; Inhibition of MEK by U0126 significantly downregulated ATRA-induced CD11b expression, whereas inhibition of PI3K by LY294002 markedly increased CD11b expression, suggesting reciprocal regulation of ATRA-induced differentiation of HL-60 cells (Fig. 8A). ATRA treatment activates MAPK pathway while inhibits Akt pathway as depicted by an increase and decrease of Erk1/2 and Akt phosphorylation respectively (Fig. 8B). Indeed, both pathways were found to regulate c-Myc expression as their inhibitions significantly decrease c-Myc expression (Fig. 8C, D). Treatment of cells with ATRA plus TIS21 led to activation of Erk1/2, but inhibition of Akt along with subsequent GSK-3activation. Regulation of the pathways was accompanied with downregulation of c-Myc protein and increase of pc-MycT58/S62 (Fig.). It is worth to mention that JNK is also involved in phosphorylation of c-Myc at serine 62 residues (Noguchi et al., 1999), however, does not contribute towards ATRA- induced granulocytic differentiation (Yen et al. 1999) and importantly its activation was not regulated by TIS21 in ATRA dependent manner. Furthermore, PP2A, Pin1 and ubiquitin ligase Skp2 that participate in c-Myc ubiquitination and proteosomal degradation were not affected by treatment with ATRA plus TIS21 (Fig. 8E). Moreover, TIS21 was not physically associated with c-Myc protein, indicating no direct regulation by TIS21 (Fig. 8F). To evaluate whether TIS21 is directly involved in the regulation of MEK/Erk1/2 and PI3K/Akt pathways, coimmunoprecipitation assays were performed; p-Erk1/2 interacted with Akt and TIS21 independent of ATRA treatment (Fig. 8G), however, treatment of HL-60 cells with ATRA
- 30 -
plus TIS21 inhibited Akt binding to p-Erk1/2. Moreover, there was no direct interaction between Akt and TIS21 (Fig. 8H), indicating the TIS21-mediated regulation of Akt and p-Erk1/2 interaction at the downstream of ATRA.
- 35 -
Fig. 8. TIS21 enhances ATRA-mediated activation of Erk1/2, and inhibited PI3K/Akt pathway along with GSK-3β activation. (A) Immunoblot analyses revealing activation of
MAPK and inhibition of Akt phosphorylation by ATRA treatment in time dependent manner. α-Tubulin was used as a loading control. (B) HL-60 cells were treated with U0126 (10 M) or LY294002 (10 M) for 2 h followed by ATRA treatment for 48 h. FACS analysis was performed to assess CD11b expression. (C, D) HL-60 cells were treated with UO126, an inhibitor of MEK, and LY294002, inhibitor of PI3K for 2 h and Erk1/2 and c-Myc (on S62 residue) phosphorylation and the reciprocal activation of p-Akt and p-Erk1/2 were measured time dependently. (E) HL-60 cells were infected for 6 h with adenoviruses (200 moi) carrying either TIS21 or LacZ gene as a transduction control and treated with low dose ATRA (0.1M) after 24 h of adenoviral transduction. Cells were harvested after 24 h of ATRA treatment and immunoblot analyses was performed. α-Tubulin was used as a loading control. (F, G) To investigate the effect of TIS21 on the interaction of p-Erk1/2 and Akt in response to ATRA treatment, immunoprecipitation was performed with anti-pErk1/2 (F) and anti-Akt antibodies both in the presence and absence of TIS21. (G) Immunoprecipitation with anti-Myc antibody and their immunoblot findings; TIS21 does not interact with c-Myc, although the amount of c-Myc was significantly decreased in the ATRA plus TIS21.
- 36 -
H. TIS21 enhanced downregulation of c-Myc by activating GSK-3β
GSK-3β is the primary kinase responsible for T58 phosphorylation and targeting c-Myc for degradationand pAkt is an upstream negative regulator of GSK-3β via phosphorylation at S9 residue, we also attempted to determine the GSK3β-pS9 status in TIS21 infected cells. To elucidate a role of GSK-3β in c-Myc degradation, HL-60 cells were treated with GSK-3β inhibitor, LiCl, in dose (~20 mM) and time (~6 hour) dependent manner, which led significant accumulation of c-Myc protein along with inhibition of pc-MycT58/S62 as opposed to GSK3β phosphorylation (Fig. 9A). We next infected cells with Ad-TIS21 (200 moi) or Ad-LacZ (200 moi) followed by treatment with ATRA (0.1μM) for 18 h, and then LiCl (20 mM) for another 6 h. Interestingly the ratio of pc-MycT58/S62 to c-Myc was markedly increased by treatment with ATRA and TIS21, whereas it was completely abolished by LiCl treatment (Fig. 9B). Moreover, ATRA-induced c-Myc degradation was significantly increased by TIS21 expression without LiCl treatment. Nevertheless, ATRA and TIS21 treatment still maintained pc-MycT58/S62 expression even in the presence of LiCl treatment, indicating that TIS21 significantly enhanced GSK-3β activity in HL-60 cells in response to ATRA and lead to phosphorylation and degradation of c-Myc protein. Treatment of HL-60 cells with LiCl revealed significant induction of c-Myc transcription by real-time PCR, in addition to GSK3Sphosphorylation (Fig. 9C). The data strongly support regulation of GSK-3β in TIS21 plus ATRA dependent manner, as it is the primary kinase responsible for c-Myc phosphorylation on T58 residue and targeting for its degradation.
- 38 -
Fig. 9. TIS21-enhanced c-Myc degradation was regulated by GSK-3β activation. (A)
GSK3β was inhibited with LiCl in dose and time dependent manner and expressions of pGSK3βS9
, pGSK3β, c-Myc and pc-MycT58/S62 were determined by Western blotting. (B) HL-60 cells were infected with TIS21 (200 moi) or LacZ control (200 moi) for 6 h and then treated with ATRA (0.1μM) at 24 h of post transduction;18 h after ATRA treatment cells were treated with LiCl (20 mM) for 6 h, harvested and subjected to Western blotting. α-Tubulin was used as a loading control. Significant inactivation of GSK-3β by phosphorylation at its S9 residue, led to the loss of pc-MycT58/S62 with marked accumulation
of c-Myc protein. (C) Transcriptional upregulation of c-Myc mRNA and protein level by treatment of HL-60 cells with 20 mM LiCl for 6 h, when examined by real time-PCR and immunoblot analyses, respectively
- 39 -
I. TIS21 enhanced ATRA-mediated c-Myc degradation in the proteosome
To further confirm the specific mechanism underlying ATRA-mediated TIS21 effects on c-Myc regulation, we investigated whether c-Myc degradation was prevented by inhibiting proteosome activity or not; Treatment of HL-60 cells with LLnL, a proteosome inhibitor, significantly accumulated c-Myc protein in concentration dependent manner (Fig. 10A), and the inhibition of proteosome activity by 20 μM LLnL with and without TIS21 and ATRA treatment significantly abrogated the TIS21 effects on ATRA-mediated c-Myc degradation with dramatic stabilization of c-Myc protein (Fig. 10B). However, our data revealed the partial recovery of c-Myc protein by LLnL (20 μM) treatment, because TIS21 expression could reduce c-Myc mRNA level via GSK-3β activation (Fig. 9E) in the presence of ATRA. It was confirmed by increase of c-Myc mRNA and protein expression by inhibiting GSK3β by LiCl treatment (Fig. 10B).
- 40 -
Fig. 10. TIS21 enhances ATRA-mediated c-Myc degradation in the proteosome. (A)
Proteasomal degradation of c-Myc protein was confirmed by treatment of HL-60 cells with LLnL, a proteasome inhibitor, in time and dose dependent manner; significant accumulation of c-Myc protein was observed. (B) HL-60 cells were transduced with TIS21 followed by ATRA as described in and then with LLnL (20 μM) for 6 h. Treatment with LLnL partially abolished the effect of TIS21 on the regulation of c-Myc degradation, indicating that TIS21 enhanced proteosomal degradation of c-Myc protein in the presence of ATRA.
- 41 -
Fig. 11. Schematic representation of ATRA plus TIS21 mediated c-Myc degradation.
The combination of ATRA and TIS21 activates Erk1/2 leading to c-Myc phosphorylation at S62, followed by phosphorylation at T58 by activated GSK3β. Dephosphorylation at S62 by PP2A leads to proteosomal degradation of c-Myc accompanied by a decrease in cells proliferation and enhance differentiation of HL-60 cells.
- 42 -
V. Discussion
The present study highlights a role of TIS21 in hematopoietic malignancy and for the first time shows its involvement in ATRA-induced in myeloid leukemia cells differentiation via c-Myc downregulation. The role of c-myc is highlighted by the finding that the abrogation of its expression induces growth arrest and terminal differentiation in leukemia cells5. Although it has been known that expression of c-Myc is significantly down-regulated both at protein and mRNA levels by ATRA treatment in HL-60 cells along with their differentiation into granulocytes, however, previous reports describe its mechanism as the elongation block of c-Myc mRNA in response to ATRA, and so far there is no report describing the synergism between ATRA and TIS21, which leads toward degradation of c-Myc protein along with HL-60 cells differentiation. Transduction of TIS21 significantly enhanced HL-60 cells differentiation and downregulation of c-Myc protein much more than that of mRNA changes in response to ATRA (Fig. 4, Fig.5). We observed that ATRA plus TIS21 decreased half life of c-Myc protein in the presence of cycloheximide (Fig. 6). The findings strongly suggest the presence of a mechanism different from the reported transcriptional regulation of c-myc mRNA elongation after ATRA treatment [Bentley et al. 1986; Bentley et al. 1988; Eick 1990]. In the present study, the molecular mechanism was suggested as the phosphorylation of c-Myc on S62 and T58 residues by pERK1/2 and GSK-3β, respectively, via activation of MAPK and inhibition of PI3K/AKT pathways by TIS21 gene, resulting in enhanced degradation of c-Myc protein along with differentiation of HL-60 cells (Fig. 8). It has been reported that ERK1/2 controls c-Myc accumulation by phosphorylation-mediated stabilization at S62, whereas subsequent T58 phosphorylation by GSK-3β is required for c-Myc degradation [Noguchi et al. 1999; Sears 2004; Welcker et al. 2008]. Recently, the 3 unique c-myc knock-in mice with either wt-c-Myc or c-Myc-T58A or c-Myc-S62A
- 43 -
phosphorylation mutant provide in vivo evidence of c-Myc's oncogenic activity regulated by phosphorylation by the two pathways [Wang et al. 2011]. Thus, MAPK pathway controls differentiation via regulating specific proteins. In contrast, PI3K/Akt pathway negatively regulates differentiation through multiple mechanisms including translation of c-Myc, translational inhibition of PDCD4, DAP5/P97, the important regulators of both granulocytic and monocytic differentiation of APL cells [Ozpolat et al. 2007; Ozpolat et al. 2008]. PI3K/Akt signaling also contributes to c-Myc stability by leading to the inhibition of GSK-3β3. Mechanistically, GSK-3β-mediated c-Myc phosphorylation potentiates PP2A-mediated dephosphorylation of S62 residue and also allows recognition by E3 ubiquitin ligase complex, SCF-Fbw7 [Arnold et al. 2008].It is worth noting that GSK-3β also decreases c-Myc transcription by increasing β-catenin degradation35
. Thus the regulation of c-Myc is achieved both at RNA and protein levels, suggesting multiple mechanisms which regulate c-Myc expression in cancer cells.
ATRA is an inducer of endogenous BTG2 as shown in Fig. 3B and the finding has been well supported by the previous reports [Passeri et al. 2006; Donato et al. 2007]. ATRA-induced Btg2 expression does not require de novo protein synthesis, rather Btg2 comprises a direct target for ATRA-induced, RAR-mediated, transcriptional signaling. A Functional RARE consisting of two direct hexameric repeats is present in the Btg2 promoter which binds RAR-RXR heterodimer for its transcriptional regulation. Therefore, activity of BTG2/TIS21 is dependent on the presence of ATRA in the process of c-Myc protein degradation as well as HL60 differentiation. Based on our previous report21, activation of Erk1/2 is a prerequisite for TIS21 phosphorylation on S147 residue that is recognized by Pin-1, indeed, ATRA (1.0 M) induced Erk1/2 phosphorylation, but inhibited Akt (Fig. 8). Interaction of TIS21 and Pin-1 allows TIS21 as a pan-cell cycle inhibitor in U937 cells and
- 44 -
induces growth arrest either in G1/S or G2/M phase [Lim et al. 1998; Guardavaccaro et al. 2000; Tirone 2001; Canzoniere et al. 2004; Ryu et al 2004] therefore, it is highly possible that potential regulation of Pin-1 activity by TIS21 might be effective on the differentiation of HL-60 cells and the TIS21-enhanced degradation of c-Myc protein in the presence of ATRA. It has been reported that Pin-1 also enhances ligand independent response of AML cells to retinoic acid [Gianni et al 2009].
It is important to mention that we directly assessed the activity of TIS21 towards c-Myc, p-Erk1/2 and Akt by examining their association. The overexpressed TIS21 did not show any association with c-Myc (Fig. 8F), however, p-Erk1/2 interacted both with TIS21 and Akt (Fig. 8G, 8H) but no interaction between Akt and TIS21 was observed (Fig. 8H). Moreover, interaction of TIS21 and p-Erk1/2 was increased by ATRA treatment, whereas interaction of p-Erk1/2 with Akt was repressed along with reduced Akt phosphorylation and GSK3β activation. Earlier reports describe that in leukemia cells Akt inhibits Erk1/2 phosphorylation while MAPK pathway positively regulates Akt activation via direct interaction. TIS21 decreases AKT-pErk1/2 physical association thus mediating Akt-MAPK reciprocal regulation in ATRA dependent manner. Moreover, ATRA translocates endogenous Btg2 from the nucleus to cytoplasm and additionally increases the interaction between pErk1/2 and TIS21 thus explaining the importance of ATRA in this process. The data strongly support the regulation of c-Myc degradation by TIS21 via activation of p-Erk1/2 and inhibition of Akt activity. The phenomenon is well accordant with the previous reports that TIS21 regulates homeostasis of bone marrow stem cells in response to estradiol treatment by regulating Akt/mTOR pathway through interaction of TIS21 with p-ERK1/2, but not Akt [Kim et al. 2008], and Akt negatively regulates Erk1/2 phosphorylation in response to vitamin D3 treatment [wang et al. 2009]. Moreover, deprivation of EGF in renal
- 45 -
tubular cells inhibits Erk1/2 activity but upregulates Akt [ Sinha et al. 2008], and hPEBP4-regulated Akt activation occurs at the downstream of Erk1/2 deactivation in prostate cancer cells [Li et al. 2007]. Therefore, activation of MAPK and inhibition of PI3K/Akt signaling by ATRA plus TIS21 significantly enhanced low dose ATRA-induced differentiation of HL-60 cells and apparently provide a mechanism leading towards the degradation of c-Myc (Fig. 11).
V. Conclusion
c-Myc promotes tumor cell proliferation and invasiveness and the tumors with c-myc amplification have poor prognosis [Yasojima et al. 2001]. The present highlights the
important of TSI21/BTG2/PC3 in myeloid leukemia cells differentiation via c-Myc downregulation. TIS21 enhanced MAPK activation and Akt inhibition decreased c-Myc expression both at transcriptional and posttranslational level and increased the differentiation process in ATRA dependent manner. Combination of TIS21 with ATRA might contribute to the therapeutic applications for a subset of AML.
- 46 -
PART –II
Regulation of Btg2
/TIS21/PC3expression under stress conditions
I. INTRODUCTION
A. NF-κB
NF-κB comprises a family of transcription factors that plays crucial roles in inflammation and immunity. The mammalian NF-κB family consists of five different related members that bind as homo or heterodimers to the target DNA sequences. These include: RelA/p65; p105 and its processing product, p50; p100 and its processing product, p52; c-Rel; and RelB. All of these family members have a Rel-homology domain (RHD) that is essential for DNA binding and dimerization. In resting state, NF-κB is sequestered in cytoplasm by IκBα inhibitory proteins, predominantly IκBα. Various mechanisms regulate the nuclear translocation and DNA binding activity of various NF-κB homo- and hetero- dimmers through canonical or non-canonical mechanisms. The basic scheme of NF-κB signaling consists of a series of positive and negative regulatory elements. Inducing stimuli trigger IKK activation leading to phosphorylation, ubiquitination, and degradation of IκB proteins. Released NF-κB dimers are further activated through various posttranslational modifications and translocate to the nucleus where they bind to specific DNA sequences and promote transcription of target genes. In its most basic form, therefore, the pathway consists of receptor and receptor proximal signaling adaptor molecules; the IKK complex; IκB proteins; and NF-κB dimmers. NF-κB regulates the expression of a diverse array of genes that are involved in regulating cell growth, differentiation, development, and apoptosis [Hatada et al. 1992; Hayden et al. 2004; Hayden et al. 2008; Vallabhapurapu et al. 2009; Morgan et al. 2011].
- 47 -
Fig. 12. The NF-κB protein family members. The number of amino acids in each protein is
indicated on the right. Inhibitory events and phosphorylation and ubiquitination sites on p100, p105, proteins that mediate proteasomal degradation are indicated with red Ps and Us, respectively. RHD, Rel homology domain; TAD, transactivation domain; LZ, leucine zipper domain; GRR, glycine-rich region; and DD, death domain (Adopted from Hayden, et al. Cell
- 48 -
Fig. 13. Mechanisms of NF-κB activation. NF-κB is activated via two distinct mechanisms;
Canonical and Noncanonical, in response to a variety of signals. Canonical mechanism invol ves IκBα degradation, while noncanonical mechanism is independent of IκBα. (Modified fro m Hayden et al. Cell 132 (2008) 344-362).
- 49 -
E. Purpose of this study
Btg2 transcription is regulated by p53. However, almost half of the cancers have abnormal p53 status (deleted or mutated) but still Btg2 expression is induced when cells are exposed to various stress challenges. How this p53 independent Btg2 regulation is achieved? Moreover, Btg2 is a primary response and stress response gene but what early signals and stress-mediated signals regulate Btg2 expression is unknown. These questions have been addressed in this project and here we have found NF-κ B as a strong inducer of Btg2 transcription when cells are exposed to oxidative stresses. Importantly this regulatory mechanism is independent of p53 expression and does not require de novo protein synthesis.
- 50 -
I. MATERIALS AND METHODS
A. MATERIALS
Antibodies against NFB RelA (p65) and lamin B1 were purchased from Cell Signaling; GF-109203X and antibodies against Btg2, IBα, SP1, and NF for Chip assay (109X) and α-tubulin were from Santa Cruz Biotechnology (Dallas, TX); PARP and anti-γH2AX were obtained from Abcam Laboratories (San Francisco, CA). LLnL, N-acetyl-L-cysteine (NAC), propidium iodide, RNase A, and polybrene were from Sigma Chemicals; Bay11-7082 and Lactacystin from Cayman Chemical (Ann Arbor, MI), and CM-H2DCF-DA was purchased
from Life Technologies-Invitrogen.
B. METHODS
1. Cells culture
Myelocytic leukemia cells (HL-60, NB4, THP-1 and U937) and anchorage-dependent colon cancer cell line DLD-1 were cultured in RPM1 1640 (Sigma, St. Louis, MO), while H9c2 and NIH3T3 cells were maintained in DMEM, supplemented with 10% heat inactivated fetal bovine serum (FBS), 1% penicillin and streptomycin, at 37 °C under 5% CO2 in air.
2. RNA extraction and semiquantitative RT-PCR
Total cellular RNAs were extracted with RNAiso Plus (Takara, Dalian), and cDNAs were synthesized with 1.0 μg of RNA using the reverse transcription kit (Invitrogen) according to the manufacturer’s instruction. Btg1, Btg2, Btg3, Tob1, Tob2, and GAPDH genes were amplified from the cDNAs by semiquantitative RT-PCR with the primer sequences shown in the Table 1. GAPDH was used as endogenous control for genes expression.
- 51 -
Table 2. List of PCR Primers
RNA Direction Sequence
Btg1 Forward Reverse 5/- CCGTCGGGGGCCGCGC-3/ 5/-CCGTGTGCTGCCTGTC -3/ Btg2 Forward Reverse 5/-CCTGGGCAGAGAGTGAAAAG-3/ 5/-CCTTCCATCCTAACCCCAAT-3/ Btg3 Forward Reverse 5/-CCGGACGAGGCTCGCTG -3/ 5/-CACAGGCTTTCAGGACATC -3/ Btg4 Forward Reverse 5/- GTTTCTCTTTCTGATCTAGC-3/ 5/-GTGCCAGTGACTTCTGTA -3/ Tob1 Forward Reverse 5/-TCTGTCCTCTGTCCTCAGC -3/ 5/-TGGGAAGCTTATTGTACAA -3/ Tob2 Forward Reverse 5/-CGTCGTCGCCGCTAGT -3/ 5/-AAGTAGGAGATGATGAAGTT -3/ GAPDH Forward Reverse 5/-CCATGGAGAAGGCTGGGG-3/ 5/-CAAAGTTGTCATGGATGACC-3/
- 52 -
3. Immunoblot analysis
Cells were harvested after washing twice with ice-cold phosphate buffered saline (PBS) and lysed with cell lysis buffer (50 mM Tris, pH 7.4, 150 mM NaCl, 1.0 mM EDTA, 1.0 mM phenylmethylsulfonyl fluoride (PMSF), 1.0 μg/ml aprotinin, 1.0 μg/ml leupeptin, 1.0 μg/ml pepstatin, 1.0 mM NaF, 1.0 mM sodium orthovanadate, 0.25% sodium deoxycholate, and 1.0 % Nonidet P-40). The proteins were separated by SDS-PAGE and transferred to PVDF membrane. Specific proteins were visualized using Western blotting detection reagent kit Ab SignalTM (AbClon, Seoul, Republic of Korea).
4. Cells fractionation
Cells were harvested after washing twice with ice cold PBS, and then lysed in 250 μl of TD buffer [25 mM Tris (base) pH 8.0, 2.0 mM MgCl2, 0.25% v/v nonident P-40, 0.5 mM
dithiothreitol (DTT), 1.0 mM PMSF] for 5 min at room temperature. The lysates were centrifuged at 1000 g for 20 min to pellet unlysed nuclei, and the supernatant (cytoplasmic fraction) was collected. The pellet was suspended in 125 μl of BL buffer [10 mM Tris (base) pH 8.0, 0.4 M LiCl, 0.5 mM DTT, 1 mM PMSF] for 5 min, sonicated, and centrifuged at 12,000 g for 20 min to pellet the cellular debris. Protein concentration of the samples was assessed by BioRad protein assay kit and analyzed by immunoblotting as described.
5. Measurement of intracellular ROS level and cell cycle analysis
Cells were cultured in media with or without serum, and stained with H2DCF-DA (20
µM) for 30 min. They were washed twice with PBS and subjected to fluorescence activated cell sorting on a FACScan flow cytometer to measure ROS level according to the manufacturer’s instruction (BD Biosciences). For cell cycle analysis HL-60 cells were exposed to serum deprivation, or Doxo, while DLD-1 cells were treated with H2O2 for 24 h.
- 53 -
were incubated for 30 min with 500 μg/ml RNase A and 50 μg/ml propidium iodide in PBS. Cell cycle profile was determined by flow cytometry according to the manufacturer’s instruction (BD Biosciences).
6. Chromatin immunoprecipitation (ChIP) assay
ChIP assay was performed as previously described [Kawakubo et al. 2004]. Briefly, cells were cross-linked with 1% formaldehyde for 10 min and then quenched using 0.125 M glycine. Nuclear fraction was isolated and lysed in SDS lysis buffer [50 mM Tris-HCl (pH 8.0)/10 mM EDTA/1%SDS] with sonication. Chromatin was immunoprecipitated overnight with anti-p65 antibody at 4 °C. Antibody complexes were incubated with protein G-beads for 2 h at 4 °C, washed four times, and eluted with 0.1% SDS/0.1 M NaHCO3. The eluates
were reverse cross-linked at 65 °C for 5 h. Following proteinase K treatment, DNA was extracted with phenol/cholroform/propanol and precipitated with ethanol-sodium acetate. Immunoprecipitated DNA was subjected to semi-quantitative PCR and analyzed by gel electrophoresis. Input templates were purified from 10% of the original lysates. Primers used to amplify BTG2 promoter were 5/-GTTCTTAGCACTGACGACAG-3/ (forward) and 5/ AAAACAGCGTTACCCTGGCC-3/ (reverse).
7. Cells Proliferation Assay
To ascertain the effect of Btg2 on cell growth and cell death, adenovirus transduction carrying Btg2 gene (Btg2) was carried out in DLD-1 and HL-60 cells. Preparation of Ad-Btg2 has been described in our previous reports [Kim et al. 2008; Lim et al. 2008]. For HL-60 cells, Ad-Btg2, infection was performed with 10 μg/mL polybrene (1,5-dimethyl-1,5-diazaundecamethylene polymethobromide). Transduction of Ad-LacZ was employed as a control. The cells infected with Ad-Btg2 or Ad-LacZ were counted microscopically at