저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Expr
essi
onandf
unct
i
onoft
heNLRP3
i
nf
l
ammasomei
nBehçet
'
sdi
sease
by
En Hyung Ki
m
Maj
ori
n Medi
ci
ne
Depar
t
mentofMedi
calsci
ences
Expr
essi
onandf
unct
i
onoft
heNLRP3i
nf
l
ammasome
i
nBehçet
'
sdi
sease
by
En Hyung Kim
A Dissertation Submitted to The Graduate School of Ajou University
in Partial Fulfillment of The Requirements for
The Degree of Ph.D. in Medical Sciences
Supervised by
Eun-So Lee, M.D., Ph.D.
Major in Medicine
Department of Medical Sciences
The Graduate School, Ajou University
This certifies that the dissertation
of En Hyung Kim is approved.
SUPERVISORY COMMITTEE
Sun Park
Dongsik Bang
Joo-Young Roh
Seonghyang Sohn
Eun-So Lee
The Graduate School, Ajou University
December18th, 2015
i
- ABSTRACT -
Expression and function of the NLRP3 inflammasome in
Behçet’s disease
Background: Behcet’s disease (BD) is a systemic inflammatory disease with manifestations
including recurrent oral and genital ulcerations, and vasculitis involving the skin, mucosa, joints, eyes, veins, arteries, nervous and gastrointestinal systems. BD is seen as a disease at the crossroad between autoimmune and autoinflammatory syndromes, possibly triggered by an aberrant response to infectious stimuli. The relevance of Gram negative bacteria-mediated oral inflammation with the increased expression of NACHT, LRR, and PYD domains-containing protein 3 (NLRP3), leading to systemic inflammation, prompted us to investigate the expression of NLRP3 inflammasome components and its link with IL-1β hypersecretion.
Purpose: The purpose of this study was to investigate whether NLRP3 inflammasome
expression is increased at the mRNA and protein levels in BD patients with skin manifestations. Also, we studied whether IL-1β production was mediated through NLRP3 inflammasome dependent pathway in BD.
Material and Methods: Peripheral blood mononuclear cells (PBMCs) from 15 active, 15
stable BD patients and 15 healthy volunteers (HC) were stimulated with lipopolysaccharide (LPS) and adenosine 5-triphosphate (ATP). The protein and mRNA expression of NLRP3 inflammasome components were analyzed by western blotting and quantitative real time PCR respectively. The in-situ expression of NLRP3 inflammasome components were
ii
analyzed by immunohistochemistry from biopsy samples of erythema nodosum (EN)-like lesions of 25 BD and 25 EN patients. Total IL-1β and mature IL-1β level were measured with ELISA
Results: When PBMCs were stimulated, the basal and LPS-induced expressions of NLRP3
inflammasome components were significantly increased at both mRNA and protein levels in BD patients compared to healthy controls. Also, increased expression of NLRP3 and ASC was observed in 25 BD skin lesions compared to 25 EN patients. Compatible with this, secretion of IL-1β by PBMCs stimulated with LPS alone or LPS plus ATP was increased in BD compared to healthy controls, which was suppressed by caspase-1 inhibitor.
Conclusion: Our findings suggest the possible link between increased IL-1β secretion and
increased expression of NLRP3 inflammasome components in BD patients with skin manifestations.
iii
TABLE OF CONTENTS
ABSTRACT --- i
TABLE OF CONTENTS --- iii
LIST OF FIGURES --- v
LIST OF TABLES --- vii
I. INTRODUCTION --- 1
II. MATERIALS AND METHODS --- 8
A. Patients and samples --- 8
B. Methods --- 8
1. Cell culture and stimulation of PBMCs --- 8
2. Quantitative real time PCR --- 9
3. Western blot --- 10 4. Immunohistochemistry --- 11 5. Image analysis --- 13 6. ELISA --- 14 7. Statistical analysis --- 14 III. RESULTS --- 15 A. Subject characteristics --- 15
B. Western blot analysis of NLRP3 inflammasome components before stimulation --- 17 C. mRNA expression levels of NLRP3 inflammasome components
iv
before stimulation --- 19
D. Immunohistochemical analysis of NLRP3 and ASC in erythema nodosum-like skin lesions of Behçet’s disease patients --- 21
E. Western blot analysis of NLRP3 inflammasome components in stimulated PBMCs from healthy control --- 25
F. Western blot analysis of NLRP3 inflammasome components in stimulated PBMCs from Behçet’s disease patients --- 27
G. mRNA expression levels of NLRP3 inflammasome components in stimulated PBMCs from Behçet’s disease patients --- 29
H. Cytokine level from stimulated PBMCs of Behçet’s disease patients --- 31
I. IL-1β level from stimulated PBMCs of Behçet’s disease patients --- 35
J. mRNA expression levels of Il-1β in stimulated PBMCs from Behçet’s disease patients --- 38
K. Expression of NLRP3 inflammasome components from stimulated PBMCs of recurrent apthous ulcer patients --- 40
IV. DISCUSSION --- 43
V. CONCLUSION --- 48
REFERENCES --- 49
v
LIST OF FIGURES
Fig. 1. Schematic representation of the nucleotide oligomerization domain-like receptors --- 4 Fig. 2. Activation of the NLRP3 inflammasome --- 6 Fig. 3. The protein expression of NLRP3, ASC, caspase-1 is increased in Behçet’s
disease before stimulation --- 18 Fig. 4. The mRNA expression of NLRP3, ASC, caspase-1 is increased in Behçet’s
disease before stimulation --- 20 Fig. 5. Immunohistochemical staining of erythema nodosum-like lesions --- 23 Fig. 6. NLRP3 and ASC expression in erythema nodosum-like skin lesions
of Behçet’s disease patients --- 24 Fig. 7. Representative western blot analysis and quantitation of NLRP3, ASC, and
caspase-1 from cell lysates of freshly isolated PBMCs --- 26 Fig. 8. The induced protein expression of NLRP3, ASC and caspase-1 is
increased in Behçet’s disease --- 28 Fig. 9. The induced mRNA expression of NLRP3, ASC and caspase-1 is
increased in Behçet’s disease --- 30 Fig. 10. The time and dose dependent release of IL-1β secretion from PBMCs
of Behçet’s disease --- 32 Fig. 11. TNF-α and IL-18 secretion from PBMCs of Behçet’s disease --- 34 Fig. 12. Total and mature IL-1β secretion is increased in PBMCs following
vi
NLRP3 activation --- 37 Fig. 13. IL-1β mRNA level is increased in PBMCs following NLRP3
activation --- 39 Fig. 14. The induced expression of NLRP3, ASC, caspase-1 and IL-1β is
increased in Behçet’s disease compared to healthy control or recurrent apthous ulcer patients ---41
vii
LIST OF TABLES
Table 1. Antibodies and their working dilutions --- 13 Table 2. Characteristics of Behçet’s disease patients and healthy controls --- 16
1
INTRODUCTION
ETIOLOGY OF BEHÇET’S DISEASE
Behçet’s disease (BD) was described as a systemic chronic inflammatory disorder characterized by recurrent oral aphtha, genital ulcers, uveitis, and skin lesions (Chajek and Fainaru, 1975; Sakane et al., 1999; Al-Mutawa and Hegab, 2004). The disease was later recognized to affect a number of systems and organs and was associated with arthritis (Mason and Barnes, 1969), thrombophlebitis (Cucuob et al., 2000), mucocutaneous (Alpsoy, 2007), muscle (Afifi et al., 1980) and neurological (Farah et al., 1998) problems, gastrointestinal (Ebert, 2008), renal (Duarte et al., 1998), cardiovascular (Atzeni et al., 2005), and pulmonary (Erkan et al., 2001) symptoms. The etiology and pathogenesis of BD have remained unclear, however both genetic factors and environment factors play a role in the pathogenesis of this disease. A complex genetic background leading to a pro-inflammatory, innate immune-system-derived activation perpetuated by adaptive immune responses against environmental and auto-antigens is accepted to be the hallmark of BD (Direskeneli et al., 2001).
BD has various aspects that deserve to be evaluated as ‘autoimmune’. Some effective treatments in BD such as azathioprine and cylophosphamide are classical immunosuppressives, and cyclosporine A is a T-cell inhibitor (Direskeneli, 2006). However, BD does not have the classical clinical features of autoimmunity such as anti-nuclear antibody (ANA) positivity, female dominance and association with other autoimmune diseases such as Sjogren’s syndrome (Yazici, 1997). Also, in contrast to classical
2
autoimmune disorders, no significant high-titer autoantibodies or antigen-specific T-cells are present (Direskeneli, 2006). A recently introduced concept to BD is ‘autoinflammation’ (Direskeneli et al., 2006). Autoinflammatory diseases are described as a group of inherited disorders characterized by episodes of seemingly unprovoked recurrent inflammatory attacks of innate nature, mainly by neutrophils (Stojanov et al., 2005). The inflammatory process, which under normal conditions is self-limiting, becomes continuous and chronic inflammatory diseases develop subsequently. Behcet’s disease, with some of its clinical features such as recurrent non-scarring mucocutaneous lesions and non-deforming arthritis, and enhanced inflammatory response with the overexpression of pro-inflammatory cytokines, is described to be in this spectrum (Gul et al., 2005). Although there are clinical and inflammatory response similarities between autoinflammatory disorders and BD, it is difficult to describe BD as either an autoimmune or an autoinflammatory disorder. An infectious agent is possibly required to trigger the innate-derived inflammation, but unlike classical autoinflammatory disorders, an adaptive response might also be sustained through ‘bacterial persistance’ or autoantigen activated antigen-presenting cells (Direskeneli et al., 2006).
THE NUCLEOTIDE OLIGOMERIZATION DOMAIN-LIKE RECEPTORS (NLR)
The innate immune system relies on its capacity to rapidly detect invading pathogenic microbes as foreign and to eliminate them. To accomplish this, cells of the innate immune system rely on several types of pattern recognition receptors (PRR) to detect both microbes and host cells that are damaged by infection. These include the Toll-like receptors
3
(TLRs), the C-type lectin receptors (CLRs), the RIG-like helicases (RLRs), cytosolic DNA sensors and members of the nucleotide oligomerization domain-like receptor (NLR) family (Bryant and Fitzgerald, 2009). The discovery of TLRs provided a class of membrane receptors that sense extracellular microbes and trigger antipathogen signaling cascades (Medzhitov and Janeway, 2002). However, due to their superficial localization in the cell, they could not account for detection of the intracellular pathogens or the internal danger signals of host cells. More recently, intracellular microbial sensors have been identified, including NLRs (Martinon et al., 2009).
Structurally and functionally, NLRs are divided into subfamilies. Structurally, NLRs are multidomain proteins with a tripartite architecture containing a C-terminal region characterized by a series of leucine rich repeat (LRR), a central nucleotide domain termed the NACHT (also called NOD) domain, and an N-terminal effector domain (Martinon et al., 2009). The LRR is a widespread structural motif of 20–30 amino acids with a characteristic pattern rich in the hydrophobic amino acid leucine (Bella et al., 2008). LRR domains are formed by tandem repeats of a structural unit consisting of a β strand and an α helix, and are organized in such a way that all the β strands and the helices are parallel to the same axis, resulting in a nonglobular, horseshoe-shaped molecule with the curved β parallel sheet lining the inner circumference and the α helices lining the outer circumference (Bella et al., 2008). The LRR domain has been implicated in ligand sensing and autoregulation of NLRs. The NACHT domain belong to STAND family of NTPases (Leipe et al., 2004). The crucial step in NLR activation lies in the oligomerization of the NACHT domain, thereby forming active, high molecular weight complexes that characterize inflammasomes (Martinon and Tschopp,
4
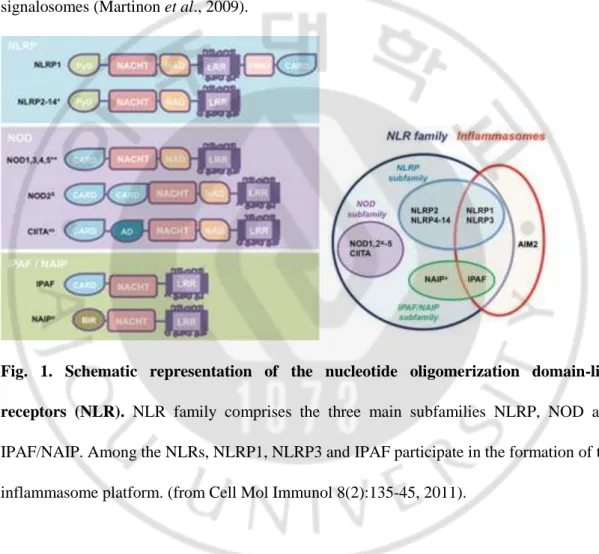
2004). The vast majority of NLRs harbor a death-fold domain at the N terminus, which is either a CARD or a pyrin domain (PYD) (Martinon et al., 2009). NLR subfamilies differ in their N-terminal effector domains, which mediate signal transduction to downstream targets, leading to activation of inflammatory caspases by inflammasomes or NF-κB by NOD signalosomes (Martinon et al., 2009).
Fig. 1. Schematic representation of the nucleotide oligomerization domain-like receptors (NLR). NLR family comprises the three main subfamilies NLRP, NOD and
IPAF/NAIP. Among the NLRs, NLRP1, NLRP3 and IPAF participate in the formation of the inflammasome platform. (from Cell Mol Immunol 8(2):135-45, 2011).
NLRs are directly or indirectly activated by pathogen associated molecular pattern (PAMPs)–containing molecules in the intracellular environment and damage-associated molecular patterns (DAMPs) in the extracellular space, such as a high concentration of adenosine triphosphate. Once activated, most proteins in the NLR family including NLRC4,
5
NLRP3, and NLRP6 provide a scaffold for assembly of a multiprotein complex called the inflammasome (Yoneyama and Fujita, 2007; Takaoka and Taniguchi, 2008).
NLRP3 INFLAMMASOME
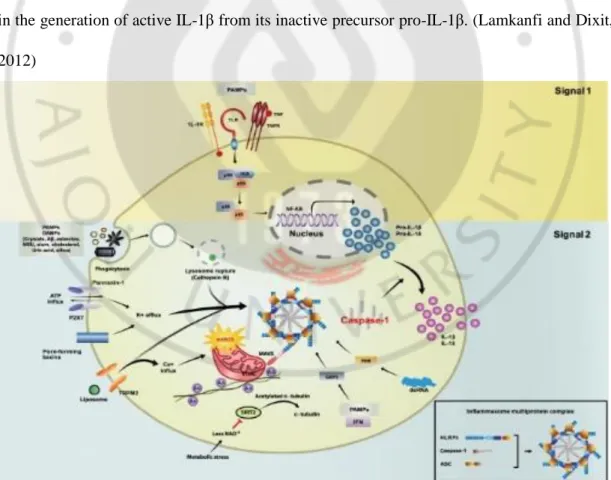
The inflammasome links the sensing of microbial products and metabolic stress to the proteolytic activation of the proinflammatory cytokines IL-1β and IL-18 (Martinon et al., 2009). One of the best-characterized NLRs is NACHT, LRR, and PYD domains-containing protein 3 (NLRP3), also known as cryopyrin and NALP3. While all inflammasomes recognize certain pathogens, it is the distinctive feature of NLRP3 inflammasome to be activated by many and diverse stimuli making NLRP3 the most versatile, and importantly also the most clinically implicated inflammasome. This molecule mediates inflammasome activation in response to several bacterial ligands, including lipopolysaccharide (LPS), peptidoglycan, bacterial and viral RNA, danger signals such as extracellular ATP and uric acid crystals (monosodium urate) (Kanneganti et al., 2006; Mariathasan et al., 2006; Martinon et al., 2006). NLRP3 may recognize a “messenger molecule” that is generated inside the cell in response to the insult that these agonists may cause.
The inflammasome cooperates with the TLR pathways to mediate a rapid response to pathogens (Dowds et al., 2004). The activation of the NLRP3 inflammasome requires two separate signals (Fig. 2). Signal 1 occurs when IL-1R, TNF or a TLR ligand binds its cognate receptor resulting in the translocation of NF-κB into the nucleus where expression of Nlrp3,
pro-IL1β, and Il-18 are induced. Signal 2 involves the assembly of NLRP3 inflammasome
6
processing and secretion of cytokines IL-1β and IL-18 (Kim and Jo, 2013). NLRP3 is the central component of the NLRP3 inflammasome; in its inactive state the LRR domain of NLRP3 is thought to self-associate, preventing interaction with CARDINAL or ASC. NLRP3 activation by agonists, such as ATP, MDP, uric acid crystals, bacterial messenger RNA, or skin irritants, unfolds the NLRP3 molecule, enabling the assembly of the inflammasome components CARDINAL, ASC, and pro-caspase-1 through homotypic interactions between their respective pyrin (PYD) and CARD domains; CARDINAL interacts with the NAD domain of NLRP3 through its FIIND domain. The oligomerization of inflammasome complexes induces cleavage of pro-caspase-1 to its active form, resulting in the generation of active IL-1β from its inactive precursor pro-IL-1β. (Lamkanfi and Dixit, 2012)
7
Peripheral blood mononuclear cells (PBMCs), particularly monocytes, which are known to be high producers of IL-1β, are reported to express NLRP3 mRNA (Kummer et al., 2007), and this was highly induced by bacterial LPS (Sutterwala et al., 2006). The fundamental involvement of inflammasome complexes in inflammatory responses has been emphasized by the fact that mutations in the NLRP3 gene are associated strongly with autoinflammatory conditions, such as rheumatoid arthritis, Muckle–Wells syndrome, Crohn’s disease, familial cold autoinflammatory syndrome (Hoffman et al., 2001; Hull et al., 2003). Several inflammatory disorders have been suggested to be IL-1β associated based on reports indicating favorable outcome after IL-1β blockade with anakinra in the disease course (Mitroulis et al., 2010). However, the role of NLRP3 inflammasome in the pathogenesis of these diseases has not been elucidated yet.
The potential involvement of NLRP3 in the pathogenesis of more common inflammatory disorders prompted us to investigate its role in BD. There are scant data on inflammasome expression in BD. Skin responses to urate crystals are described in BD (Gogus et al., 2005). Urate crystals are recently shown to activate NLRP3 inflammasome, causing the activation of caspase-1 complex and leading to the release of IL-1β (Martinon et
al., 2006).
In this study, we investigated whether NLRP3 inflammasome expression is increased in BD patients with skin manifestations. Also, we studied whether NLRP3 inflammasome expression and cytokine release was altered after stimulation in BD patients.
8
II. MATERIALS AND METHODS
A. Patients and samples
The patient population consisted of 15 active and 15 stable BD patients, who presented themselves for the first time or were monitored at the Department of Dermatology, Ajou University Hospital. BD patients met the Diagnostic criteria of the BD Research Committee of Japan. The active group patients had at least one of the BD symptoms despite the treatment and inactive group patients were in well-controlled states by taking anti-inflammatory medication. The control groups consisted of 15-newly diagnosed recurrent apthous ulcer patients without any other evident disease and 15 healthy volunteers as disease and healthy control groups, respectively. The informed consent was obtained from patients prior to enrolling them into the study. This study was approved by the Institutional Review Board (IRB no.: AJIRB-GN3-07-098, AJIRB-GGEN-GEN-10-119).
B. Methods
1. Cell culture and stimulation of PBMCs
Venous blood was sampled in sodium citrate-containing cell preparation tubes (Vacutainer CPT; BD Biosciences, San Jose, CA). Peripheral blood mononuclear cells (PBMCs) were separated by Ficoll Hypaque density gradients (Ficoll paqueTIM plus,
9
StemCell Technologies, Vancouver, BC, Canada). Cells from the interphase were collected and washed twice with phosphate buffered saline (PBS; Sigma, ST. Louis, MO) with 2% heat-inactivated fetal bovine serum (FBS; Gibco-BRL, Grand Island, NY).
PBMCs (1x106 cell/ml) were resuspended in culture medium (RPMI 1640 medium supplemented with 2 mM L-glutamine, 100 U/㎖ penicillin and 100 ㎍/ml streptomycin) with 10% FBS. The cells were incubated with either 100 uL of culture medium (negative control) or various concentrations of lipopolysaccharide (LPS; Sigma-Aldrich, St Louis, MO) as described in the figure legends. The supernatants were collected and stored at -70°C until assayed. Cells were harvested at various times, 1, 4 and 24 hr, after the incubation at 37℃ and 5% CO2. To investigate the effect of LPS followed by the second stimulus of ATP on cytokine production, PBMCs were initially stimulated for 4 hours with LPS. After 4 hours, RPMI containing various concentrations of adenosine 5-triphosphate (ATP; Sigma-Aldrich) was added to the cells for another 15, 30 and 60 minutes as described in the figure legends (LPS/ATP). In separate experiments, 20 uM zYVAD(Ome)-FMK an irreversible caspase-1 inhibitor (CaspI; Enzo life science, PlymouthMeeting, PA) was added.
2. Quantitative real time PCR
Total RNA was extracted using Trizol® (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Reverse transcription of RNA was performed using dNTP and oligo (dT) primers (Invitrogen, Carlsbad, CA, USA) and using Superscript™ III
10
(Invitrogen, Carlsbad, CA, USA) following the recommendations of the manufacturer. Real-time PCR analysis was performed using primers and internal probes for NLRP1 (Hs00248187_m1), NLRP3 (Hs00918082_m1), ASC (Hs00203118_m1), Caspase-1 (Hs00354836_m1), IL-1β (Hs01555410_m1) and GAPDH (Hs99999905_m1), which were purchased as Assays on Demand primer-probe (Applied biosystems, Seoul, Korea). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the reference gene to normalize the RNA expression levels. Real-time PCR was performed on ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster, CA, USA) using TaqMan® Gene Expression Assays (Applied Biosystems, Foster, CA, USA) according to the manufacturer’s instructions. Subsequently, a Ct value was obtained for each sample. Quantification was done using the 2−ΔΔCt method. The ΔCt values were calculated for each gene of interest as follows: ∆Ctsample = (Ctsample − CtGAPDH) Relative mRNA levels were calculated by the expression 2–∆∆Ct, where ∆∆Ct= (∆Ctsample − ∆Ctcalibration). The calibrator sample (∆Ct calibration) was assigned from the HC group.
3. Western blot
Freshly isolated PBMCs and stimulated PBMCs were harvested and lysed in RIPA buffer (Sigma–Aldrich, St. Louis, MO, USA) containing protease inhibitors. Cell extracts were run on Bolt™ 4−12% Bis−Tris Plus Gel (life technologies, Carlsbad, CA, USA) and transferred onto polyvinylidene difluoride membranes (Merck Milipore, Darmstadt, Germany). Following transfer, the membrane was blocked overnight at room temperature
11
with PBS/0.2% Tween-20/5% skim milk. Blots were incubated with primary antibodies Anti-NLRP3, anti-apoptosis-associated speck-like protein containing a CARD (ASC), and anti-caspase-1 antibodies (Abcam, Cambridge, MA). The membranes were incubated in a solution containing an appropriate secondary Ab (either anti-rabbit IgG or anti-mouse IgG Ab) linked to horseradish peroxidase (Invitrogen). Bands were visualized with Immobilon Western Chemiluminescent HRP Substrate (Merck Milipore, Darmstadt, Germany).
Western blots were quantified using ImageJ analysis. The images were set to grayscale 8-type bit and a rectangular box was drawn to enclose a single lane. The box was then selected as the first lane and the same process was repeated for the remaining of the lanes. The relative density of the contents of each lane was then plotted as histograms and a straight line was drawn underneath where the peak ends to subtract any background. The wand tool was then used to get the measurements of each profile. Bands were normalized to β-actin by dividing the average intensity of the band by the average intensity of the β-actin band from the same sample labeled on the same gel.
4. Immunohistochemistry
Six mm punch skin biopsies of erythema nodosum (EN)-like lesions of 25 BD and 25 EN patients were performed. Formalin-fixed and paraffin-embedded tissues of EN-like lesions in BD were cut (3-μm thickness) and mounted onto slides. Specimens were deparaffinated and endogenous peroxidase activity was blocked by 3% H2O2 in methanol for
12
15 minutes at room temperature. After rinsing in phosphate-buffered saline (PBS) for 10min, the nonspecific binding sites were blocked by blocking solution for 10 minutes at room temperature. To determine the cells infiltrating EN-like lesions, a panel of antibodies directed against mononuclear cell antigens using an indirect avidin-biotin immunoperoxidase technique. The antibodies used are listed on Table 1. The degree of inflammatory cell infiltration, which also showed positivity to NLRP3 in serial section, were graded on a semi-quantitative scale of 0-4: 0, absent; 1, minimal; 2, mild; 3, moderate; 4, marked staining. All slides were evaluated by three examiners in a blinded fashion and the average score was calculated for each section.
To examine the expression of NLRP3 inflammasome components of EN-like lesions and EN, the above specimens were incubated with polyclonal antibodies against NLRP3 and ASC for 30 min at room temperature (Table 1). Next, HRP polymer (Thermo scientific, Fremont, CA, USA) was applied and incubated for 30 minutes at room temperature. After washing in PBS for 10min, bound antibodies were visualized by incubation with AEC chromogen system (Thermo scientific, Fremont, CA, USA). Slides were counterstained with hematoxylin. Negative controls were isotype matched. The image was analyzed using Image Pro Plus Version 4.5 (Media Cybertics Co., MD, U.S.A.)
13
Table 1. Antibodies and their working dilutions
Specificity/ Antibody
Mono / Poly Source Working
Dilution
NLRP3 Mouse polyclonal Alexis Biochemicals, San diego, CA, 1:50 ASC Rabbit polyclonal Lifespan bioscience, Seattle, WA 1:100 CD3 Mouse polyclonal Novocastra, Newcastle, UK 1:200
CD20 Mouse polyclonal Dako, Denmark 1:300
CD68 Mouse polyclonal Novocastra, Newcastle, UK 1:80 MPO Rabbit polyclonal NeoMarker, Fremount, CA 1:600
5. Image analysis
Image signals were recorded on a personal computer and evaluated using Image Pro Plus Version 4.5 (Media Cybernetics Co., Silver Spring, MD, USA). For each staining, we established a standard for antibody (Ab) positivity and applied the same standards to the samples. The stained area per total area was measured and the ratio of positive Ab area to the septal and lobular panniculitis area was calculated. Each measurement was evaluated under constant magnification (×200). As inflammation presents mainly in and around the septa, fat lobules were left out when calculating the area. The image analysis was performed on a representative area of each specimen and repeated three times by three examiners and the
14
mean was used for evaluation. Finally the data was expressed as fold difference between EN-like lesion and EN.
6. ELISA
Total IL-1β, mature IL-1β, TNF-α and IL-18 level in the supernatants was measured with a commercial ELISA kit from R&D Systems according to the manufacturer's protocols.
7. Statistical analysis
The data are presented as mean ± standard deviation (S.D). Data were analyzed by one-way analysis of variance followed by the Scheffé test for overall multiple comparisons among HC, stable BD, active BD. Student's t-test was used for comparison between two groups. SPSS 17.0 (SPSS Inc., Chicago, IL) was used. A p-value < 0.05 was considered to indicate statistical significance.
15
III. RESULTS
A. Subject characteristics
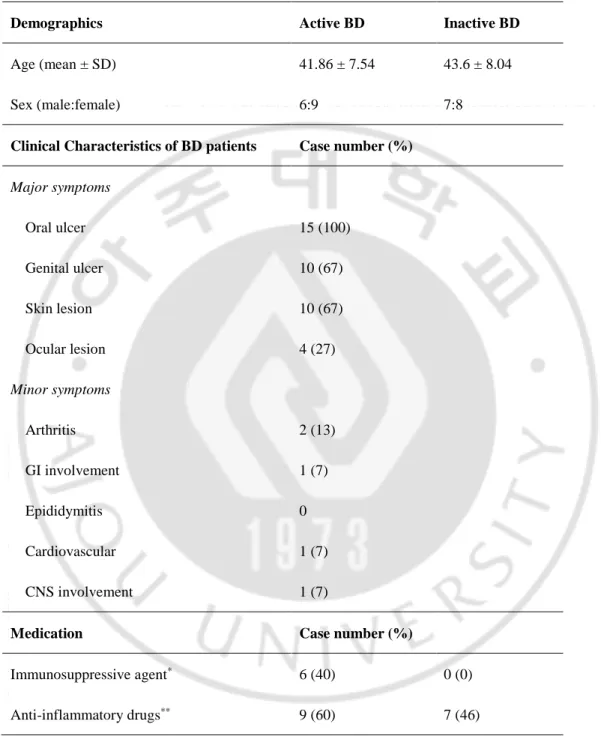
A total of 60 subjects entered the study (15 active BD patients, 15 stable BD patients, 15 RAU patients, and 15 of healthy controls). Baseline clinical findings of BD patients are summarized in Table 2. Active BD patient was defined as those who had more than one active clinical symptom such as oral ulcer, genital ulcer, skin lesion, ocular inflammation, GI, CNS or joint involvement at that time of examination. Of 30 BD patients, 15 (6 males, 9 females) were in the clinically active state with mean age of 41.86 yr. 15 (7 males, 8 females) were inactive state with mean age of 43.60 yr. In the active BD patients, 15 patients had mucocutaneous symptoms, including oral ulcer, genital ulcer, or skin manifestation, and 9 patients other symptoms, including uveitis, gastrointestinal, central nervous system or joint symptom. Six active BD patients were taking immunosuppressive and 9 patients were taking anti-inflammatory drugs when blood samples were taken. Seven inactive BD patients were taking anti-inflammatory drugs intermittently but not when blood samples were taken.
16
Table 2. Characteristics of Behçet’s disease (BD) patients and healthy controls
Demographics Active BD Inactive BD
Age (mean ± SD) 41.86 ± 7.54 43.6 ± 8.04
Sex (male:female) 6:9 7:8
Clinical Characteristics of BD patients Case number (%)
Major symptoms Oral ulcer 15 (100) Genital ulcer 10 (67) Skin lesion 10 (67) Ocular lesion 4 (27) Minor symptoms Arthritis 2 (13) GI involvement 1 (7) Epididymitis 0 Cardiovascular 1 (7) CNS involvement 1 (7)
Medication Case number (%)
Immunosuppressive agent* 6 (40) 0 (0)
Anti-inflammatory drugs** 9 (60) 7 (46)
* methylprednisolone 8-24 ㎎/d
17
B. Western blot analysis of NLRP3 inflammasome components before
stimulation of PBMCs from Behçet’s disease patients compared to healthy
control
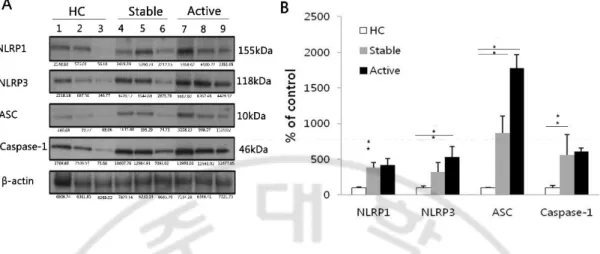
We investigated the protein levels of the different components of NLRP3 inflammasomes, and NLRP1 for comparison. The protein expression of NLRP1, NLRP3, ASC and caspase-1 from cell lysates of freshly isolated PBMCs of BD patients was analyzed by western blotting. A representative result of Western blotting is shown in Fig. 3A. Bands were normalized to β-actin by dividing the average intensity of the band by the average intensity of the β-actin band from the same sample labeled on the same gel. The result showed that the mean ± S.D values of normalized NLRP3 levels were significantly up-regulated in active (0.80 ± 0.44) and stable BD (0.49 ± 0.07) compared with HC (0.15 ± 0.02) (p < 0.05). The mean ± S.D values of normalized ASC protein levels were significantly up-regulated in active (0.20 ± 0.08) and stable BD (0.10 ± 0.10) compared with HC (0.01 ± 0.00) (p < 0.05). The mean ± S.D values of normalized caspase-1 protein levels were significantly up-regulated in active (1.65 ± 0.36) and stable BD (1.50 ± 0.41) compared with HC (0.27 ± 0.22) (p < 0.05). The mean ± S.D values of normalized NLRP1 protein levels were significantly up-regulated in active (0.60 ± 0.34) and stable BD (0.56 ± 0.28) compared with HC (0.14 ± 0.02) (p < 0.05) (Fig. 3B).
18
Fig. 3. The protein expression of NLRP3, ASC, caspase-1 is increased in Behçet’s
disease (BD) before stimulation. Representative western blot analysis (lane 1-3: healthy
control, lane 4-6: stable BD, lane 7-9: active BD) (A) and quantitation of NLRP1, NLRP3, ASC and caspase-1 from cell lysates of freshly isolated PBMCs (B). β-actin was used as loading control. (n=5 per group) Data are represented as mean ± S.D. (* p < 0.05), (−) no treatment. NLRP3: NACHT, LRR, and PYD domains-containing protein 3; PBMCs: peripheral blood mononuclear cells; HC: healthy volunteers.
19
C. mRNA expression levels of NLRP3 inflammasome components before
stimulation of PBMCs from Behçet’s disease patients compared to healthy
control
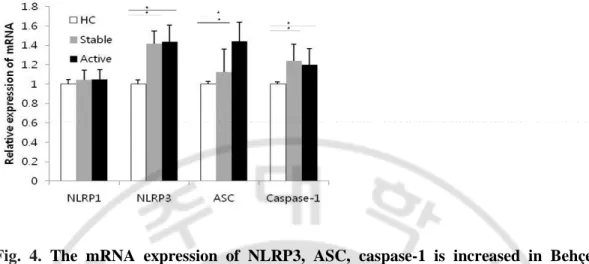
We investigated the mRNA levels of the different components of NLRP3 inflammasomes, and NLRP1 for comparison. The mRNA expression of NLRP1, NLRP3, ASC and caspase-1 from cell lysates of freshly isolated PBMCs of BD patients was analyzed by quantitative real time PCR. NLRP3, ASC and caspase-1 mRNA expression was significantly up-regulated in active and stable BD compared with HC (p < 0.05). However there were no significant difference in NLRP1 mRNA expression among active and stable BD and HC (Fig. 4).
20
Fig. 4. The mRNA expression of NLRP3, ASC, caspase-1 is increased in Behçet’s
disease (BD) before stimulation. The mRNA expression of NLRP1, NLRP3, ASC and
caspase-1 was measured from freshly isolated PBMCs by real time PCR and normalized against the expression levels of glyceraldehyde 3-phosphate-dehydrogenase. The relative values are shown as a fold change to HC with no treatment. (n=8 per group) Data are represented as mean ± S.D. (* p < 0.05), (−) no treatment. NLRP3: NACHT, LRR, and PYD domains-containing protein 3; PBMCs: peripheral blood mononuclear cells; HC: healthy volunteers.
21
D. Immunohistochemical analysis of NLRP3 and ASC in erythema
nodosum-like skin lesions of Behçet’s disease patients
To correlate our in vitro findings we examined skin lesions for in situ expression of NLRP3 and ASC. EN-like lesion is one of cutaneous manifestations of BD. It is characterized by tender, erythematous subcutaneous nodules commonly affecting the lower extremities. Histopathologic features are the septal pannicular inflammation associated with some peripheral lobular inflammation and a superficial and deep dermal lymphocytic inflammatory infiltrate. The degree of inflammatory cell infiltration were graded on a semi-quantitative scale of 0-4: 0, absent; 1, minimal; 2, mild; 3, moderate; 4, marked staining. The semi-quantitative analysis of inflammatory cell infiltration was as follows; CD3+ T lymphocytes (2.01 ± 1.11), CD20+ B lymphocytes (0.75 ± 1.05), CD68+ monocytes/ macrophages (3.14 ± 0.65), MPO+ neutrophils (2.69 ± 1.27). Thus the EN-like lesions were infiltrated with CD3+ T lymphocytes, CD68+ monocytes and macrophages, and Myeloperoxidase (MPO)+ neutrophils (Fig. 5).
The cellular expression of NLRP3 and ASC in skin lesions of BD and control EN patients were detected by immunohistochemistry. NLRP3 was expressed in inflammatory cells infiltrating the subcutaneous tissue. When compared in serial section staining, NLRP3 was expressed by a proportion of CD68+ monocytes/macrophages, MPO+ neutrophils but was rarely found in CD3+ T cells. As for ASC, it was also detected in CD68+ monocytes/macrophages, MPO+ neutrophils. The stained area per total area was measured and the ratio of positive Ab area to the septal and lobular panniculitis area was calculated
22
using Image analysis. Then the fold difference between EN-like lesion and EN for NLRP3 and ASC was calculated. The result showed that NLRP3 (mean ± S.D. for EN-like lesion of BD vs EN; 2.34 ± 1.14 vs 1 ± 0.49) (p < 0.05) and ASC (mean ± S.D. for EN-like lesion of BD vs EN; 4.92 ± 0.33 vs 1 ± 0.79) (p < 0.05) expression was significantly increased in EN-like lesion of BD compared to EN (Fig. 6).
23
Fig. 5. Immunohistochemical staining of erythema nodosum-like lesions. Erythema
nodosum-like lesions are infiltrated with CD3+ T lymphocytes, CD68+ monocytes and macrophages, and Myeloperoxidase (MPO)+ neutrophils. (A) anti-CD3, (B) anti-CD20 , (C) anti-CD68, (D) anti-MPO antibody (200x) (n=25).
24
Fig. 6. NLRP3 and ASC expression in erythema nodosum-like skin lesions of Behçet’s disease patients. Immunohistochemistry staining of skin lesions of BD patients and control
EN lesions with anti-NLRP3 (left) and anti-ASC (right) antibody showed NLRP3 and ASC expression in inflammatory cells infiltrating the subcutaneous tissue (200x). Computer assisted image analysis showed significant increase in NLRP3 and ASC stained area in BD patients. The values are shown as a fold change to EN. The smaller box is a magnified region (400x) (n=25 per group). Data are represented as mean ± S.D. (* p < 0.05), (−) no treatment. NLRP3: NACHT, LRR, and PYD domains-containing protein 3; PBMCs: peripheral blood mononuclear cells; HC: healthy volunteers; EN: erythema nodosum.
25
E. Western blot analysis of NLRP3 inflammasome components in stimulated
PBMCs from healthy control
The precise mechanism leading to the activation of the NLRP3 inflammasome is still unclear. It is proposed that intracellular activities such as induction of hypokalemia, reactive oxygen species, or calcium-dependent phospholipase2 indirectly activate the inflammasome (Pétrilli et al., 2007). However, stimulation of cells solely with ATP, a known inducer of potassium efflux through P2X7-mediated mechanisms, is unable to activate caspase-1, and NF-кВ dependent transcription of NLRP3 by TLR ligands such as LPS is necessary to induce inflammasome activation (Netea et al., 2010).
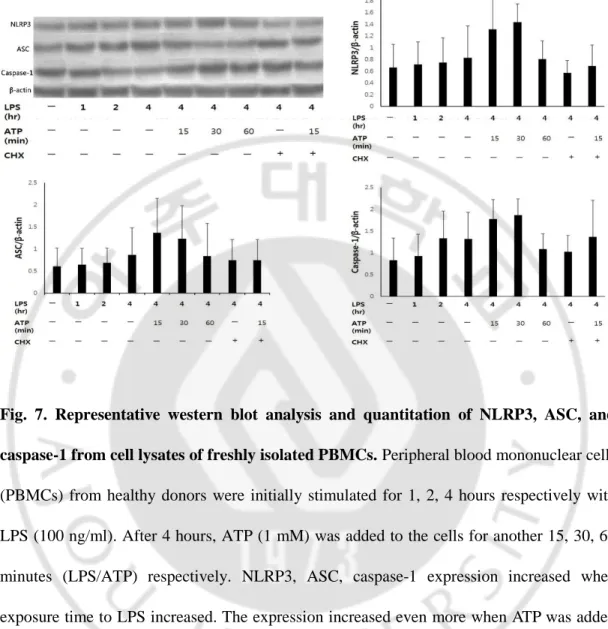
We checked the increase of NLRP3, ASC, and caspase-1 protein productions along the time course using PBMCs from healthy donors to see whether LPS alone, or ATP stimulation after LPS priming (LPS/ATP) increased the components of NLRP3 inflammasomes (Fig 7). NLRP3, ASC, caspase-1 expression increased when exposure time to LPS increased. The expression increased even more when ATP was added for 15 mins and 30 mins respectively. However after 60 mins, the expression level decreased. This may be because prolonged ATP exposure can lead to cell death by apoptosis or lysis/necrosis (Gonnord et al., 2009). The previously mentioned increase in protein expression may be due to decrease in protein degradation or an increase in protein translation rate. When protein synthesis inhibitor cycloheximide (10μg/ml) was added the protein level decreased to LPS stimuli only suggesting that ATP had a role in de novo protein synthesis.
26
Fig. 7. Representative western blot analysis and quantitation of NLRP3, ASC, and caspase-1 from cell lysates of freshly isolated PBMCs. Peripheral blood mononuclear cells
(PBMCs) from healthy donors were initially stimulated for 1, 2, 4 hours respectively with LPS (100 ng/ml). After 4 hours, ATP (1 mM) was added to the cells for another 15, 30, 60 minutes (LPS/ATP) respectively. NLRP3, ASC, caspase-1 expression increased when exposure time to LPS increased. The expression increased even more when ATP was added for 15 mins and 30 mins respectively. However after 60 mins, the expression level decreased. When protein synthesis inhibitor cycloheximide (CHX) (10μg/ml) was added the protein expression levels of NLRP3, ASC, caspase-1 decreased. β-actin was used as loading control.(n=5 per group).
27
F. Western blot analysis of NLRP3 inflammasome components in stimulated
PBMCs from Behçet’s disease patients compared to healthy control
As expression of the components of the NLRP3 inflammasome are increased in BD patients, we then examined whether LPS alone, or ATP stimulation after LPS priming (LPS/ATP), affected the protein expression of NLRP3 inflammasome components in PBMCs of BD patients. Then we compared the results among no treatment, after LPS alone, and ATP stimulation after LPS priming (LPS/ATP) and also among active BD, stable BD and HC. The results showed that the protein levels of, NLRP3, ASC and caspase-1 were significantly increased in active and stable BD compared to HC. Following LPS/ATP stimulation, NLRP3 and ASC protein levels were significantly up-regulated in active BD compared to HC (Fig. 8).
28
Fig 8. The induced protein expression of NLRP3, ASC and caspase-1 is increased in Behçet’s disease (BD). PBMCs were initially stimulated for 4 hours with LPS (100 ng/ml).
After 4 hours, ATP (1 mM) was added to the cells for another 15 minutes (LPS/ATP). Representative western blot analysis and quantitation of NLRP3, ASC and caspase-1 from cell lysates of stimulated PBMCs. β-actin was used as loading control. (lane 1,4,7: no treatment, lane 2,5,8: LPS, lane 3,6,9: LPS/ATP) (n = 5 per group). Data are represented as mean ± S.D. (* p < 0.05), (-) no treatment. NLRP3: NACHT, LRR, and PYD domains-containing protein 3; PBMCs: peripheral blood mononuclear cells; LPS: lipopolysaccharide; ATP: adenosine 5-triphosphate; HC: healthy volunteers.
29
G. mRNA expression levels of NLRP3 inflammasome components in stimulated
PBMCs from Behçet’s disease patients compared to healthy control
As expression of the components of the NLRP3 inflammasome are increased in BD patients, we examined whether LPS alone, or ATP stimulation after LPS priming (LPS/ATP), affected the mRNA expression of NLRP3 inflammasome components in PBMCs of BD patients. Then we compared the results among no treatment, after LPS alone, and ATP stimulation after LPS priming (LPS/ATP) and also among active BD, stable BD and HC. The result of quantitative real time PCR showed that the mRNA levels of, NLRP3, ASC and caspase-1 were higher after LPS/ATP stimulation compared to single stimulus of LPS in all the groups. Furthermore, they were significantly increased in the presence of LPS or LPS/ATP in active and stable BD compared to HC (Fig. 9).
30
Fig 9. The induced mRNA expression of NLRP3, ASC and caspase-1 is increased in Behçet’s disease (BD). PBMCs were initially stimulated for 4 hours with LPS (100 ng/ml).
After 4 hours, ATP (1 mM) was added to the cells for another 15 minutes (LPS/ATP). The mRNA expression of NLRP3, ASC and caspase-1 was measured by quantitative real time PCR and normalized against the expression levels of glyceraldehyde 3-phosphate-dehydrogenase. The relative values are shown as a fold change to HC with no treatment. (n=8 per group) Data are represented as mean ± S.D. (* p < 0.05), (-) no treatment. NLRP3: NACHT, LRR, and PYD domains-containing protein 3; PBMCs: peripheral blood mononuclear cells; LPS: lipopolysaccharide; ATP: adenosine 5-triphosphate; HC: healthy volunteers.
31
H. Cytokine level from stimulated PBMCs of Behçet’s disease
The increased expression of components of the inflammasome after LPS/ATP stimulation raises the possibility that IL-1β level can also be increased after LPS/ATP stimulation, as activated inflammasomes induce IL-1β production and secretion. To ascertain whether the induction of NLRP3 inflammasome components contributes to increased secretion of IL-1β in BD patients, we assessed IL-1β secretion by PBMCs stimulated with LPS or LPS/ATP. PBMCs from BD patients were treated with different levels of LPS for 1, 4 and 24 hrs. The IL-1β level after stimulation with 10ng LPS was 18.70 ± 9.49, 21.53 ± 13.55, 26.87 ± 9.85 respectively for 1hr, 4hr and 24hr of stimulation. The IL-1β level after stimulation with 100ng LPS was 22.77 ± 10.13, 26.22 ± 12.55, 31.34 ± 13.07 respectively for 1hr, 4hr and 24hr of stimulation. The IL-1β level after stimulation with 1ug LPS was 25.42 ± 11.46, 29.00 ± 12.59, 35.32 ± 12.23 respectively for 1hr, 4hr and 24hr of stimulation. Thus IL-1β level showed a tendency to increase with LPS dose and priming time (Fig. 10A). The IL-1β level after 4hr LPS priming and 15 minutes stimulation with control, 0.5mM, 1mM, 2.5mM, 5mM, 10mM of ATP were 4.08 ± 0.70, 133.53 ± 15.99, 146.02 ± 7.36, 156.55 ± 9.08, 156.88 ± 6.82, 186.80 ± 5.03, 211.17 ± 4.43 respectively. The result showed that IL-1β level had a tendency to increase with ATP dose (Fig. 10B).
32 A.
B.
Fig. 10. The time and dose dependent release of IL-1β secretion from PBMCs of Behçet’s disease. PBMCs were stimulated with (A) 10ng, 100ng and 1ug of LPS for 1hr,
4hr and 24hr and with (B) 0.5mM, 1mM, 2.5mM, 5mM, 10mM of ATP after 4hr LPS priming. IL-1β secretion was quantitated in the supernatant of PBMCs by ELISA. Data are represented as mean ± S.D. (-) no treatment. NLRP3: NACHT, LRR, and PYD domains-containing protein 3; PBMCs: peripheral blood mononuclear cells; LPS: lipopolysaccharide; ATP: adenosine 5-triphosphate; HC: healthy volunteers.
33
Tumour necrosis factor (TNF)-α is an important proinflammatory cytokine. It has been implicated in the pathogenesis of a number of inflammatory disorders. TNF-α also plays an important role in the immunopathogenesis of Behcet’s disease. Increased levels of circulating TNF-α and soluble TNF receptors in the peripheral blood of BD patients with active disease, as well as high levels of TNF in the aqueous humor from patients with BD associated uveitis have been reported (Chronopoulou et al., 2001). IL-18 is cleaved to the bioactive form through inflammasomes mediated caspase-1 activation similar to IL-1β. Serum IL-18 is also increased in BD. So we studied the level of TNF-α and IL-18 level from LPS and LPS/ATP stimulated PBMCs of BD to see whether the expression show similar trend to IL-1β (Fig.11). The TNF-α level after LPS (729.06 ± 63.02) and LPS/ATP (885.25 ± 39.38) was higher compared to no treatment. There was only a small difference between LPS and LPS/ATP. The IL-18 level after LPS (11.47 ± 6.43) and LPS/ATP (42.93 ± 10.99) was higher compared to no treatment and the increase after LPS/ATP was much higher than LPS alone.
34
A.
B.
Fig. 11. TNF-α and IL-18 secretion from PBMCs of Behçet’s disease. PBMCs were
initially stimulated for 4 hours with LPS (100 ng/ml). After 4 hours, ATP (1 mM) was added to the cells for another 15 minutes (LPS/ATP). (A) TNF-α and (B) IL-18 secretion was quantitated in the supernatant of PBMCs by ELISA. (n= 4 per group). Data are represented as mean ± S.D. (-) no treatment. PBMCs: peripheral blood mononuclear cells; LPS: lipopolysaccharide; ATP: adenosine 5-triphosphate.
35
I. IL-1β level from stimulated PBMCs of Behçet’s disease patients compared to healthy control
Next, we examined whether IL-1β secretion from stimulated PBMCs of Behçet’s disease was more significant compared to healthy control. In accordance with previous reports (Netea et al., 2010) showing that peripheral blood monocytes stimulated with LPS release ATP and consequently secrete IL-1β, treatment of PBMCs with LPS alone (mean ± S.D for HC, stable BD and active BD were 28.15 ± 6.83, 111.66 ± 32.90, 104.49 ± 34.93 respectively) significantly increased IL-1β secretion compared to no stimulation (mean ± S.D for HC, stable BD and active BD were 1.27 ± 1.51, 2.26 ± 2.14, 1.93 ± 1.14 respectively) (p < 0.05). As caspase-1 is known to convert pro IL-1β to mature secreted form, we investigated the requirement for caspase-1 function in ATP induced IL-1β release. LPS primed PBMCs were inhibited with caspase-1 inhibitor, zYVAD(Ome)-FMK. IL-1β secretion was significantly suppressed by caspase-1 inhibition (mean ± S.D for HC, stable BD and active BD were 9.11 ± 4.24, 16.07 ± 11.80, 33.49 ± 25.91 respectively) compared to priming with LPS (p < 0.05). IL-1β secretion was significantly higher in BD patients than in HC (Fig. 12A).
Mature IL-1β (17kD) level was significantly higher after LPS/ATP (mean ± S.D for HC, stable BD and active BD were 51.54 ± 19.75, 80.67 ± 15.81, 84.64 ± 20.40 respectively) compared to LPS alone (mean ± S.D for HC, stable BD and active BD were 10.34 ± 11.54, 22.37 ± 16.78, 26.28 ± 18.75 respectively) or no treatment (p < 0.05). The mature IL-1β secretion in the presence of LPS/ATP was significantly higher in active and stable BD
36
patients than in HC. Similar to total IL-1β, mature IL-1β secretion was significantly suppressed by caspase-1 inhibition (mean ± S.D for HC, stable BD and active BD were 8.69 ± 3.84, 18.56 ± 3.31, 26.41 ± 5.16 respectively) compared to LPS/ATP (p < 0.05) (Fig. 12B).
37 A.
B.
Fig 12. Total and mature IL-1β secretion is increased in peripheral blood mononuclear cells (PBMCs) following NLRP3 activation. PBMCs were initially stimulated for 4 hours
with LPS (100ng/ml) with or without 20 μM zYVAD(Ome)-FMK, an irreversible caspase-1 inhibitor (LPS/CaspI). After 4 hours, ATP (1 mM) was added to the cells for another 15 minutes (LPS/ATP or LPS/ATP/CaspI). (A) Total IL-1β (n=15 per group) and (B) mature processed IL-1β (n=9 per group) was quantitated in the supernatant of stimulated PBMCs by ELISA. Data are represented as mean ± S.D. (* p < 0.05), (-) no treatment. PBMCs: peripheral blood mononuclear cells; NLRP3: NACHT, LRR, and PYD domains-containing protein 3; LPS: lipopolysaccharide; ATP: adenosine 5-triphosphate.
38
J. mRNA expression levels of Il-1β in stimulated PBMCs from Behçet’s disease patients compared to healthy control
Quantitative real time PCR was performed to examine whether the increase of total and mature IL-1β correlated with the expression of IL-1β mRNA. IL-1β mRNA expression was increased after LPS/ATP stimulation compared to no stimulation or LPS stimulation alone.
There were significant differences in LPS-induced and LPS/ATP-induced IL-1β mRNA levels between active BD and HC. However, inhibition with caspase-1 inhibitor, zYVAD(Ome)-FMK caspase-1 inhibitor failed to decrease in IL-1β mRNA levels (Fig. 13). This may be because first, LPS binding to TLRs can induce IL-1β mRNA transcription (Netea et al., 2009). Second, IL-1β may have been released before caspase-1 inhibitor had effect. LPS may induce active release of endogenous ATP to the extracellular environment with consequent activation of the P2X7R in an autocrine loop leading to IL-1β secretion (Piccini et al., 2008). Also, freshly isolated primary blood monocytes contain activated form of caspase-1, i.e. constitutively activated caspase-1 leading to release of IL-1β (Netea et al., 2009).
39
Fig. 13. IL-1β mRNA level is increased in peripheral blood mononuclear cells (PBMCs) following NLRP3 activation. PBMCs were initially stimulated for 4 hours with LPS
(100ng/ml) with or without 20 μM zYVAD(Ome)-FMK, an irreversible caspase-1 inhibitor (LPS/CaspI). After 4 hours, ATP (1 mM) was added to the cells for another 15 minutes (LPS/ATP or LPS/ATP/CaspI). The mRNA expression of IL-1β was measured by real time PCR and normalized against the expression levels of glyceraldehyde 3-phosphate-dehydrogenase. The relative values are shown as a fold change to HC with no treatment. (n=8 per group) Data are represented as mean ± S.D. (* p < 0.05), (-) no treatment. PBMCs: peripheral blood mononuclear cells; NLRP3: NACHT, LRR, and PYD domains-containing protein 3; LPS: lipopolysaccharide; ATP: adenosine 5-triphosphate.
40
K. Expression of
NLRP3 inflammasome components
from stimulated PBMCs ofBehçet’s disease patients compared to recurrent apthous ulcer patients (RAU)
The increase in the expression of NLRP3 inflammasome components and IL-1β may be a coincidental finding due to tissue damage and release of DAMP. So we examined the NLRP3 inflammasome components expression and total IL-1β level of recurrent apthous ulcer (RAU) patients, who also show tissue damage. The result showed that the increase of both NLRP3 inflammasome components expression and total IL-1β level were more significant in BD compared to RAU (Fig. 14).
41
A. B.
C.
42 E.
Fig. 14. The induced expression of NLRP3, ASC, caspase-1 and IL-1β is increased in Behçet’s disease (BD) compared to healthy control or recurrent apthous ulcer patients (RAU). PBMCs were initially stimulated for 4 hours with LPS (100ng/ml) with or without
20 μM zYVAD(Ome)-FMK, an irreversible caspase-1 inhibitor (LPS/CaspI). After 4 hours, ATP (1 mM) was added to the cells for another 15 minutes (LPS/ATP or LPS/ATP/CaspI). (A-D) The mRNA expression of NLRP3, ASC, caspase-1 and IL-1β was measured by real time quantitative RT-PCR and normalized against the expression levels of glyceraldehyde 3-phosphate-dehydrogenase. The relative values are shown as a fold change to HC with no treatment. (n=8 per group). (E) Total IL-1β (n=15 per group) was quantitated in the supernatant of stimulated PBMCs by ELISA. Data are represented as mean ± S.D. (* p < 0.05), (-): no treatment; LPS: lipopolysaccharide; ATP: adenosine 5-triphosphate; CaspI: caspase-1 inhibitor; HC: healthy volunteers, RAU: Recurrent apthous ulcer (disease control).
43
IV. DISCUSSION
BD is a systemic inflammatory disorder characterized basically by recurrent oral ulcers, genital ulcers, and uveitis. At times, BD may evolve to a widespread disorder involving the skin, joints, and the pulmonary, vascular, and central nervous systems. Although there are clinical and inflammatory response similarities between autoinflammatory disorders and BD, presence of a prolonged inflammation such as non-specific (pathergy) or urate induced skin responses suggests that innate and adaptive pathways are more integrated in BD (Gogus et al., 2006). Also, in contrast to classical autoimmune disorders, no significant high-titre autoantibodies or antigen-specific T-cells are present (Yazici, 1997). Thus it seems too simplistic to describe BD as either an autoimmune or an autoinflammatory disorder. The hypothesis of an environmental trigger in BD patients with genetics susceptibilities has long been advocated. These triggers range from infections to molecular mimicry such as heat-shock proteins (HSP), which are synthesized under cellular stress (Pineton de Chambrun et al., 2012). Recognition of triggers by innate immune cells induce inflammatory signals to combat infection and prevent disease. However, the same mechanism can also lead to uncontrolled inflammation resulting in tissue injury and disease. For example an infectious agent is possibly required to trigger the innate-derived inflammation, but unlike classical autoinflammatory disorders, an adaptive response might also be sustained through ‘bacterial persistance’ or autoantigen activated antigen-presenting cells (Direskeneli et al., 2006).
44
infections. Given the high frequency of oral ulcer and the decrease of some symptoms with minocycline, oral flora has been implicated in the pathogenesis of BD (Kaneko et al., 1997; Calguneri et al., 1996) Recently it was shown that NLRP3 inflammasome is upregulated when infected by Porphyromonas gingivalis, a G(-) bacteria, and in some cases lead to induction and sustained aortic or gingival inflammation (Yamaguchi et al., 2015). Also, peptidoglycan and LPS induced IL-1β via the TLR2/4 and reactive oxygen species–NLRP3 inflammasome-dependent pathways in ocular BD (Liang et al., 2013). As oral ulcer precede BD for many years and EN like lesions, one of the most common skin manifestations in BD, which are characterized by septal and lobular panniculitis and most importantly vasculitis, we investigated whether NLRP3 inflammasome expression is increased in BD patients with skin manifestations. We showed that NLRP3, ASC and caspase-1 mRNA and protein levels were upregulated in PBMCs of BD patients. This was further supported by in situ findings that showed NLRP3 and ASC expression was significantly increased in EN-like lesions of BD. These findings show that mRNA and protein levels of NLRP3 inflammasome components are increased in BD patients. This may be a coincidental finding as necrotic cells release endogenous molecules, such as heat shock proteins, uric acid, ATP, and DNA that alert the innate immune system of the danger associated with tissue damage and infection (Rock and Kono, 2008). These molecules act as endogenous “danger signals” by activating a variety of pattern recognition receptors, including TLR and NLR (Mariathasan et
al., 2006). However the increase was more significant compare to RAU, which also shows
tissue damage. Thus increase in NLRP3 inflammasomes may have a role in BD pathogenesis. NLRP3 inflammasome is activated by canonical stimuli like ATP or Nigericin and
45
noncanonical stimuli like live gram negative bacteria (Bauernfeind et al., 2013). Therefore, we checked whether LPS alone, or ATP stimulation after LPS priming (LPS/ATP), affected the expression of NLRP3 inflammasome components in PBMCs of BD patients. The protein levels of, NLRP3, ASC and caspase-1 were higher following LPS stimulation compared to no stimulation in all the groups and the levels increased significantly in active and stable BD compared to HC. Following LPS/ATP stimulation, NLRP3 and ASC protein levels were significantly up-regulated only in active BD compared to HC. The mRNA levels of, NLRP3, ASC and caspase-1 were higher after LPS/ATP stimulation compared to single stimulus of LPS in all the groups. Furthermore, they were significantly increased in the presence of LPS or LPS/ATP in active and stable BD compared to HC. These findings show that LPS/ATP stimulation resulted in significantly higher expression of NLRP3 inflammasome component at protein and mRNA levels in PBMCs of BD patients. Thus in BD, formation of NLRP3 inflammasome may be mediated by two signals, first a PAMP binds to its PRR and second, a danger signal, such as ATP, ligates to its purinergic receptor P2X7.
The function of NLRP3 inflammasome is to detect the presence of microbial products, toxins, and danger signals in the cytoplasm leading to caspase-1 activation, a necessary step for the processing and secretion of the immature forms of IL-1β, IL-18, and IL-33. IL-1β has significant role in BD pathomechanism. Specific IL-1β gene polymorphisms cause increased susceptibility to BD and IL-1β blockade with recombinant anti-IL-1β antibody gevokizumab has been used for BD treatment (Gul et al., 2012) Thalidomide, an anti-inflammatory drug, used to treat BD, was found to decrease IL-1β through caspase-1 inhibition (Keller et al., 2009). These are some of the studies that indicate IL-1β plays an important role in the
46
pathogenesis of BD. IL-1β synthesis, maturation, and secretion are tightly regulated by TLR signaling and inflammasome activation (Dinarello, 2009). Unlike most cytokines, IL-1β lacks a secretory signal peptide and is externalized by monocytic cells through a nonclassic pathway, arranged in 2 steps (Rubartelli et al., 1990; Andrei et al., 1999). First, TLR ligands, such as LPS, induce gene expression and synthesis of the inactive IL-1β precursor (proIL-1β). Monocytes stimulated with LPS alone release only ~20% of the IL-1β over 24–48 hours (Dinarello, 1998). A second stimulus, such as exogenous ATP, strongly enhances the proteolytic maturation and secretion of IL-1β (Perregaux and Gabel, 1994; Solle et al., 2001). The emergence of NLR proteins and the inflammasome complex as critical components for this processing step suggests that they may have a role in BD. However there are only limited data on the influence of NLRP3 inflammasome in BD. To ascertain whether the increased NLRP3 inflammasome components might contribute to increased secretion of IL-1β in BD, we assessed IL-IL-1β secretion by PBMCs stimulated with LPS or LPS/ATP. In accordance with previous reports (Netea et al., 2010) showing that peripheral blood monocytes stimulated with LPS release ATP and consequently secrete IL-1β, treatment of PBMCs with LPS alone increased IL-1β secretion compared to no stimulation. This effect was suppressed by caspase-1 inhibition and was significantly higher in BD compared to HC. Additionally, mature IL-1β secretion in the presence of LPS/ATP was significantly higher in active and stable BD than HC and suppressed by caspase-1 inhibitor. There were significant differences in LPS-induced and LPS/ATP-induced IL-1β mRNA levels between BD and HC. However, caspase-1 inhibitor suppressed mature IL-1β secretion in the presence of LPS/ATP without a decrease in mRNA levels. These findings suggest that LPS/ATP stimulation
47
induced IL-1β expression. And higher expression of NLRP3 inflammasome components in BD might contribute to increased IL-1β secretion in BD patients.
Pathways of inflammasome and pathogen-associated molecular pattern such as TLRs intersect as both are sensors of bacterial products. Previous studies have shown that TLR2 and TLR4 are expressed in inflammatory cells of BD patients (Do et al., 2008). LPS from oral microorganisms could initiate IL-1β synthesis process through TLRs and the inflammasomes. The extracellular ATP released from stressed or infected cells in oral tissue may further activate NLRP3 inflammasomes leading to IL-1β maturation and release. The released IL-1β can activate the release of other proinflammatory cytokines. Thus after a trigger such as oral infection initiates the innate-derived inflammation, an adaptive response might be sustained through NLRP3 inflammasomes. Therefore understanding the mechanism of NLRP3 in BD will not only help to elucidate their role in BD pathogenesis, but may also contribute to understanding the host responses to infectious agents and to danger signals. Further studies on the functional effect and activation pathway of NLRP3 inflammasomes in BD are needed to elucidate the exact role of NLRP3 inflammasomes in BD.
48
V. CONCLUSION
1. Expression of NLRP3, ASC and caspase-1 protein and mRNA in PBMCs from Behçet’s disease patient was significantly up-regulated in active and stable BD compared with HC. Also, NLRP3 and ASC expression was significantly increased in BD skin lesions
2. The protein levels of NLRP3, ASC and caspase-1 increased significantly in active and stable BD compared to HC following LPS stimulation. Following LPS/ATP stimulation, NLRP3 and ASC protein levels were significantly up-regulated in active BD compared to HC
3. The mRNA levels of, NLRP3, ASC and caspase-1 were significantly increased in the presence of LPS or LPS/ATP in active and stable BD compared to HC.
4. Treatment of PBMCs with LPS alone increased IL-1β secretion compared to no stimulation. Mature IL-1β secretion in the presence of LPS/ATP was significantly higher in active and stable BD than HC. These findings suggest that LPS/ATP stimulation induced IL-1β expression.
This study showed that the expression of NLRP3 inflammasome by PBMCs of Behçet's disease patients are increased at baseline and after LPS/ATP stimulation. And this led to increased level of total IL-1ß and mature IL-1ß. These findings may imply that after a trigger such as oral infection initiates the innate-derived inflammation, an adaptive response might be sustained through NLRP3 inflammasomes.in Behçet’s disease.
49
REFERENCES
1. Afifi AK, Frayha RA, Tekian A: The myopathology of Behcet’s disease – a histological, light-, and electron microscopic study. J Neurol Sci. 48: 333-332, 1980
2. Al-Mutawa SA, Hegab SM: Behcet's disease. Clin Exp Med 4: 103-131, 2004
3. Al-Otaibi LM, Porter SR, Poate TW: Behçet's disease: a review. J Dent Res 84: 209-222, 2005
4. Alpsoy E, Zouboulis CC, Ehrlich GE: Mucocutaneous lesions of Behcet's disease.
Yonsei Med J 48: 573-585, 2007
5. Andrei C, Dazzi C, Lotti L, Torrisi MR, Chimini G, Rubartelli A: The secretory route of
the leaderless protein interleukin 1 β involves exocytosis of endolysosome-related vesicles. Mol Biol Cell 10:1463–75, 1999
6. Atzeni F, Sarzi-Puttini P, Doria A, Boiardi L, Pipitone N, Salvarani C: Behçet's disease and cardiovascular involvement. Lupus 14: 723-726, 2005
50
7. Bauernfeind FG1, Horvath G, Stutz A, Alnemri ES, MacDonald K, Speert D, Fernandes-Alnemri T, Wu J, Monks BG, Fitzgerald KA, Hornung V, Latz E: Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol 183:787–791, 2009
8. Bauernfeind F, Hornung V: Of inflammasomes and pathogens--sensing of microbes by the inflammasome. EMBO Mol Med 5:814-826, 2013
9. Bella J, Hindle KL, McEwan PA, Lovell SC: The leucine-rich repeat structure. Cell Mol.
Life Sci.65(15):2307–2333, 2008
10. Benoist, C., Mathis, D: Autoimmunity provoked by infection: how good is the case for T cell epitope mimicry? Nat. Immunol 2: 797– 801, 2001
11. Bryant C, Fitzgerald KA: Molecular mechanisms involved in inflammasome activation. Trends Cell Biol 19(9): 455-464, 2009
51
12. Calguneri M, Kiraz S, Ertenli I, Benekli M, Karaarslan Y, Celik I: The effect of prophylactic penicillin treatment on the course of arthritis episodes in patients with Behcet's disease. A randomized clinical trial. Arthritis Rheum 39:2062-2065, 1996
13. Chajek T, Fainaru M: Behçet's disease. Report of 41 cases and a review of the literature.
Medicine (Baltimore) 54: 179-196, 1975
14. Chronopoulou H, Tzavara N, Ekonomopoulos N, Zafirakis P, Theodosiadis P, Tzioufas T: High levels of TNFα in the aqueous humor in patients suffering from adamantiades-Behcet’s disease: preliminary results. Invest Ophthalmol Vis Sci 42 (Suppl. 4): S708, 2001
15. Conforti-Andreoni C, Ricciardi-Castagnoli P, Mortellaro A: The inflammasomes in health and disease: from genetics to molecular mechanisms of autoinflammation and beyond. Cell Mol Immunol 8(2):135-45, 2011
16. Cucuob P, Sbai A, Wechsler B, Brocherion I, Braesco J, Kieffer E, Piette JC: Vascular manifestations of Behcet’s syndrome associated with solitary ulcerations and resolved with immunosuppressants. Rev Med Intern 21: 353–357, 2000